
記住我







Kinesin-3 motors are built with unique mechanical outputs, such as high velocity, superprocessivity and a strong MT-binding affinity than other kinesin family motors [17,18,19,20]. A single kinesin-3 motor can take thousands of steps before detaching from the microtubule and is fueled by ATP hydrolysis [18]. Despite rigorous motility analysis over the last few years [22, 25,26,27,28,29,30], the basic chemomechanical properties of these motors remain poorly studied. Often such analysis requires purified, soluble, active motor proteins. The prokaryotic expression system is used widely for recombinant protein expression. As a control, we used a constitutively active version of kinesin-1, KHC(1–560), the founding member of the kinesin superfamily, whose motility properties are well characterized. The expression and purification of KHC(1–560) in bacteria resulted in multiple low molecular bands (Additional file 1: Fig. S1A, B) and protein degradation. Additionally, single-molecule motility analysis resulted in rare motility events and majority of them were bound non-specifically to the glass surface. Studies have shown that bacterial expression and purification of motor proteins usually result in a large population of dead motors due to improper protein folding and/or limited capacity of the host system [31,32,33,34]. Thus, an additional step of microtubule affinity purification is critical to eliminate dead and inactive motor fractions to some extent [35, 36].
To overcome these setbacks, we used baculovirus expression, one of the most powerful and versatile eukaryotic expression systems. Baculovirus has a strong polyhedrin promoter to drive the high-level expression of heterologous genes. In addition, this system employs the ability of cultured Sf9 cells to perform post-translational modification of expressed proteins, similar to those that occur in the natural host cell. Thus, we generated bacmids to purify full-length and constitutively active versions of kinesin-3 and kinesin-1 motors (Additional file 1: Fig. S2A-C) and subjected them to a one-step purification approach (Additional file 1: Fig. S3A-C). Size exclusion-high-performance liquid chromatography (SEC-HPLC) and circular dichroism (CD) analysis of these purified motor proteins showed a homogeneous protein population with regular secondary structures and protein folding, respectively. For representation, HPLC elution profile and CD spectra of KIF1A(1-393LZ)-mCit-FLAG are shown in Additional file 1: Fig. S3D, E. These Sf9-purified motors were used for detailed biochemical and biophysical characterizations.
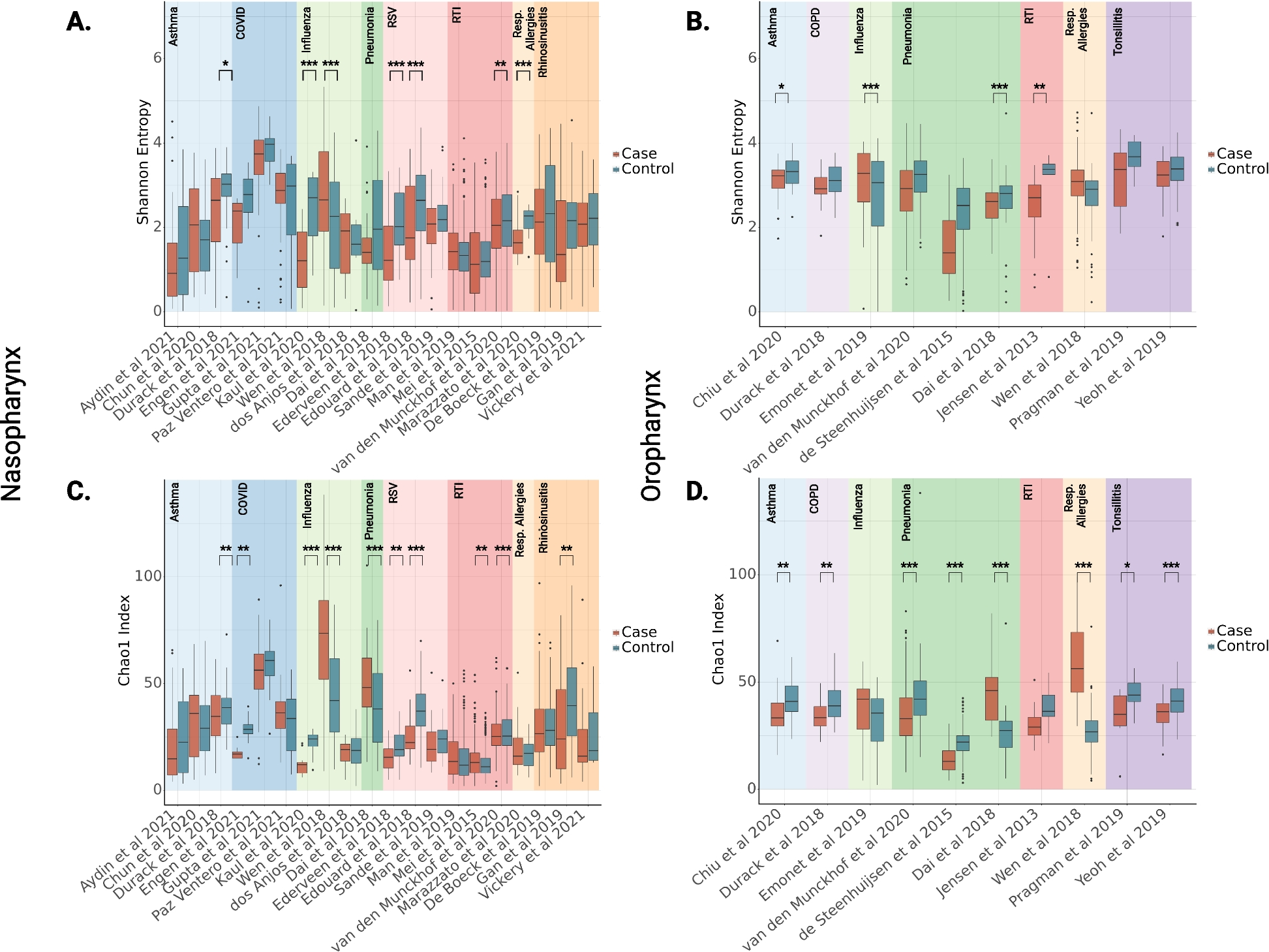
Sf9-purified kinesin-3 motors are fast and superprocessiveFirst, we wanted to establish whether Sf9-purified kinesin-3 motors can support MT-based superprocessive motility. We performed in vitro single-molecule motility assays of dimeric active kinesin-3 motors as described previously [17, 18]. The control constitutively active kinesin-1, KHC(1–560) motor, showed processive motion along the MT tracks with an average velocity of 0.82 µm s−1 and 1.14 ± 0.04 µm run length (Fig. 1A; Additional file 1: Fig. S4A; Table 1) [6, 7, 18, 37]. As kinesin-1 motor takes 8-nm steps [7, 8, 38], the measured velocity renders a stepping rate (average number of steps taken in 1 s) of 102.15 s−1 and a mean run time (average time spent on the MT) of 1.39 ± 0.3 s with a motor off-rate (frequency of motor detachment from the MT in 1 s) 0.72 ± 0.05 s−1.
Fig. 1
Sf9-baculovirus-purified kinesin-3 motors are robust and superprocessive. A–E Fluorescently tagged truncated constitutively active motors were purified using Sf9-baculovirus expression system. In vitro single-molecule motility assays of Sf9-purified A kinesin-1, KHC (1–560) and B–E kinesin-3 motor B KIF1A(1-393LZ), C KIF13A(1-411ΔP), D KIF13B(1-412ΔP), and E KIF16B(1–400) motors. For each population of motors, histograms of velocities (left panel) and run lengths (right panel) were plotted and fit to a single Gaussian. Average velocity and run length of the corresponding population of motors (N) are indicated on top-right or left corner as mean ± SEM. Data presented from three independent experiments
Table 1 Biochemical and biophysical properties of kinesin-3 family motorsNext, single-molecule motility analysis of Sf9-purified active dimeric kinesin-3 motors displayed robust superprocessive motion with high velocities along the MT (Fig. 1B–E; Additional file 1: Fig. S4B-E; Table 1) when compared to kinesin-1. It has been shown that the wild-type KIF1A(1–393) motors exhibit diffusive motion and short processive motion in vitro owing to weak neck coil dimerization potential [18, 28]. Therefore, we used its stable version, KIF1A(1-393LZ), in which a leucine zipper (LZ) segment of GCN4 was fused to the C-terminus of KIF1A(1–393) [18]. The KIF1A(1-393LZ) motor traveled with an average speed of 2.41 ± 0.02 µm s−1 and a distance of 10.56 ± 0.26 µm (Fig. 1B; Additional file 1: Fig. S4B; Table 1), akin to our previous measurements using mammalian cell lysates [17, 18]. As KIF1A motors take 8-nm step/ATP hydrolysis [22, 23, 28], the determined velocity corresponds to a stepping rate of 301.29 s−1, which is approximately threefold faster than kinesin-1. Furthermore, considering the determined average run length and the velocity over the total track, we computed a motor average run time of 4.38 ± 1.3 s, which corresponds to an off-rate of 0.23 ± 0.05 s−1.
The wild-type KIF13A(1–411) and KIF13B(1–412) motors exist as inactive monomers due to proline-mediated intramolecular neck coil-coiled coil1 (NC-CC1) interaction [18]. We, therefore, used proline-deleted dimeric versions, KIF13A(1-411ΔP390) and KIF13B(1-412ΔP391), respectively. The motility analysis of KIF13A(1-411ΔP390) [from now on referred to as KIF13A(1-411ΔP)] and KIF13B(1-412ΔP391) [from now on referred to as KIF13B(1-412ΔP)], exhibited average velocities of 1.55 ± 0.02 µm s−1 and 1.36 ± 0.01 µm s−1 and run lengths of 10.24 ± 0.22 µm and 10.96 ± 0.22 µm, respectively (Fig. 1C,D; Additional file 1: Fig. S4C-D; Table 1). Assuming these motors take 8-nm steps, their velocities reveal stepping rates of 194.19 s−1 and 170.16 s−1, respectively. Furthermore, based on the measured average run lengths and the velocities over the total track, we computed an average run time of 6.61 ± 2.6 s and 8.06 ± 2.4 s, which corresponds to motor detachment rates of 0.15 ± 0.07 s−1 and 0.12 ± 0.09 s−1, respectively.
Furthermore, motility analysis with active KIF16B(1–400) also exhibited long uniform motion along the MT with an average velocity of 0.95 ± 0.01 µm s−1 and run length 9.34 ± 0.22 µm (Fig. 1E; Additional file 1: Fig. S4E; Table 1). These motility properties showed remarkable consensus with previously measured kinesin-3 motility properties using mammalian cell lysate [17,18,19, 23, 39]. Considering KIF16B takes 8-nm steps, the determined velocity translates to a stepping rate of 118.75 s−1. Based on the measured run length and the velocity over the entire track, we found that the motor run time of 9.83 ± 1.2 s corresponds to a motor off-rate of 0.10 ± 0.02 s−1.
Together, these results demonstrate that the functional output of motor proteins purified from the Sf9-baculovirus system are comparable to those expressed in mammalian cells. Their motility properties are on par with those measured previously using mammalian cell lysates. The results also suggest that the high velocity and superprocessivity of kinesin-3 motors is inherent to their motor domains. Notably, this one-step motor purification protocol can be adapted to purify any other protein of interest.
Kinesin-3 motors exhibit high ATP turnover ratesThe ability of kinesin motors to take processive steps along the microtubule is tightly coupled to the ATP hydrolysis cycle because binding of ATP causes conformational changes in the motor domain. Studies on kinesin-1 and myosin have directly correlated their velocity and ATP turnover rate [4, 40]. However, previous chemomechanical studies of kinesin-3 motors have reported significantly lower ATP turnover rates [21, 25, 26, 28], which does not correlate with the measured high velocity and superprocessive motility [18, 19]. Thus, understanding the MT-stimulated chemomechanical behavior of kinesin-3 motors can help to elucidate the observed unique kinesin-3 motility properties. Therefore, we decided to do MT-stimulated ATPase measurements of full-length and constitutively active kinesin-3 motors. To do this, we adapted an ATPase assay based on phosphomolybdate complex formation, as described for smooth muscle myosin [41].
As a control, we used a dimeric KHC(1–560), constitutively active kinesin-1 motor whose catalytic rate constants have been well characterized, to optimize assay conditions. First, we determined the optimal motor concentration required to measure the ATPase activity by assaying a range of KHC(1–560) motor concentrations (1 to 100 nM) with a constant MT concentration. We found that a motor concentration of 10 nM is optimal for reliable and consistent measurement of ATPase activity across the preparations [24]. Next, we determined the range of MT concentrations desired to accurately measure ATPase activity for the individual motor type, depending on when the ATPase activity attains its steady-state maxima. For reliable ATPase measurements, an assay mixture containing purified motor protein, ATP, and varying MTs concentrations was incubated at room temperature for 2 h. Samples were collected every 30 min and inorganic phosphate release was measured [41].
The assay with full-length KHC showed low basal ATPase activity even after adding MTs (Fig. 2A; Additional file 2: Table S1) [42, 43]. Consistent with the fact that kinesin-1 predominantly exists in a compact autoinhibited conformation, the tail domain folds back to interact with the motor domain directly [44,45,46]. This folded tail-to-head intramolecular interaction precludes the motor-MT association and subsequent ATP hydrolysis and holds the motor in an inactive folded conformation [47,48,49,50]. In contrast, constitutively active dimer, KHC(1–560), displayed approximately tenfold higher activity following the addition of MTs (Fig. 2B; Table 1). The determined kcat and Km were 95.44 ± 15.5 s−1 and 10.23 ± 3.63 µM, respectively, which agrees with previous reports [4, 44, 51, 52]. The ratio of the speed of kinesin-1 to its rate of MT-stimulated ATP hydrolysis yields a step size of ~ 8.54 nm and stoichiometry of 1.07 step/ATP hydrolyzed, suggesting a tight coupling between their chemical and mechanical cycles [6, 7].
Fig. 2
Kinesin-3 motors exhibit high ATPase activity and differential microtubule affinity. MT-stimulated ATPase activity of full-length and truncated constitutively active motors was measured using Sf9-purified proteins. A Comparison of ATPase activity between full-length and constitutively active kinesin motors. ATPase activity for full-length motors was measured at MT concentration that had highest ATPase activity for the respective constitutively active motor. B–F Plots showing ATPase activity against varied concentrations of MTs for B constitutively active kinesin-1 motor, KHC (1–560) and kinesin-3 motors C KIF1A(1-393LZ), D KIF13A(1-411ΔP), E KIF13B(1-412ΔP), and F KIF16B(1–400) and fit to Michaelis–Menten equation. Maximal turnover (kcat) and MT affinity (Km) parameters were determined using GraphPad Prism. Error bars represent mean ± SD. Data presented from three independent experiments
A similar analysis using bacterially purified KHC(1–560) (Additional file 1: Fig. S1) exhibited deficient activity, indicating that majority of the motors were inactive. This finding again lends support to the importance of purifying kinesin motors from the eukaryotic expression system rather than from the prokaryotic system. Together, these results support the general autoinhibition mechanism in the field proposed for kinesin-1 and that the stepping rate of kinesin-1 is directly proportional to the speed of ATP hydrolysis.
Full-length kinesin-3 motors exhibit low ATPase activitiesNext, we measured the ATPase activity of kinesin-3 family motors with and without MTs. Analogous to kinesin-1, the addition of MTs to full-length kinesin-3 motors did not stimulate their ATP hydrolysis (Fig. 2A; Additional file 2: Table S1). This observation is consistent with non-cargo-bound full-length kinesin-3 motors that exist in an autoinhibited state. Autoinhibition of motors prevents unnecessary hydrolysis of cellular ATP and interference with cargo trafficking on the MT. For kinesin-3, studies have shown that intramolecular interaction between the NC and CC1 domains keeps the motor in a monomeric, autoinhibited state [18, 53,54,55,56,57,58,59,60]. However, the detailed mechanism of kinesin-3 autoinhibition is still poorly understood.
KIF1A dimers exhibit the highest ATP turnover ratesUnlike full-length motors, the addition of MTs to constitutively active, dimeric kinesin-3 motors remarkably stimulated their ATPase activity, which is 1.3- to threefold higher than kinesin-1 (Fig. 2C–F). The rate of ATP hydrolysis for dimeric active KIF1A(1-393LZ) motor [17, 18] was found to be kcat 290 ± 9 s−1, which is 3 times faster than the activity of KHC(1–560) (Fig. 2C; Table 1). The determined high kcat is in close agreement with the estimated stepping rate (~ 301.29 s−1) with a step size of ~ 8.28 nm and stoichiometry of 1.0 step/ATP, suggesting a tight coupling between chemical and mechanical cycles. A similar step size of ~ 8 nm has been reported recently for KIF1A and UNC-104 motors [23]. Together, these results explain the fast velocity reported for this motor [17, 18, 28].
The measured ATPase activity is considerably higher than pioneering work from Hirokawa lab that demonstrated an ATP turnover rate of 110 ± 5 s−1 for a monomeric KIF1A [25, 26]. Similarly, an elegant and detailed experimental analysis of active UNC104 dimers showed a lower ATP turnover rate of 100 s−1/head [28]. Recent work from Zaniewski et al. also reported a significantly lower kcat 115 s−1 for a stable KIF1A(1–406) dimers, suggesting a significant fraction of the molecules may be inactive, presumably due to motors purified from bacterial expression system [52]. The determined higher MT affinity of KIF1A(1-393LZ) (Km 7.03 µM) compared to KHC(1–560) (Km = 10.23 µM) agrees with previous studies on KIF1A/CeUnc104 [17, 26, 28].
KIF13 dimers show higher ATPase activityContrary to full-length KIF13 motors, the addition of MTs to the dimeric active KIF13A(1-411ΔP) and KIF13B(1-412ΔP) motors strongly stimulated their ATPase rates with kcat of 181.7 s−1 and 171.8 s−1, respectively (Fig. 2D,E; Table 1). These rates match their measured in vitro velocities (1.55 µm/s and 1.36 µm/s) and estimated stepping rates of 194.19 s−1 and 170.16 s−1, respectively [17, 18, 49]. Similar to KIF1A, the ratio of KIF13A and KIF13B motor velocities with their rates of ATP hydrolysis generates a step size of 8.53 and 7.91 nm, respectively, yielding a stoichiometry of ~ 1, suggesting a tight relation between ATP hydrolysis and motor stepping. Additionally, the measured ATPase rate constants of KIF13 motors were twice the rate of kinesin-1, corresponding to the observed difference in their velocities [18, 30, 49]. However, earlier studies using the truncated KIF13B(1–368) motor containing a catalytic motor domain and neck linker exhibited MT-stimulated ATP turnover rate 45 s−1 and Km of 0.1 µM [61]. The observed low catalytic activity was presumably due to motor truncation lacking the dimerization domain (NC) and purified from bacterial expression system.
Similarly, previous MT-stimulated ATPase measurements for KIF13B full-length and motor truncation after FHA domain showed low ATP turnover rates (kcat = 0.3 s−1 and kcat = 4.6 s−1, respectively) [
留言 (0)