
記住我

Given their highly potent therapeutic activity, fewer side effects, and cost-effectiveness, natural products may be promising candidates for pharmaceutical development as anticancer, anti-autoimmunity, and anti-allergic agents. There is a family of natural compounds called flavonoids that are found widely in flowers and fruits in the form of benzo-gamma-pyrone derivatives. Quercetin is one of these natural products [1]. Quercetin (3,3′,4′,5,7-pentahydroxyflavone) represents the most abundant flavonoid in human diets and can be found at high concentrations in apples, onions, red wine, red grapes, tea (Camelia sinensis), capers, broccoli, lovage, and a wide range of berries [2, 3]. Various cancer cell types, including liver cancer, have been shown to be affected by antineoplastic activity of quercetin via its influence on proliferation, differentiation, and apoptosis [4]. Additionally, quercetin downregulates multiple signal transduction pathways, including NF-κB, MEK–ERK, PI3K–Akt–mTOR, Wnt–β-catenin, and Nrf2–keap1, which contribute to inflammation and carcinogenesis processes [5,6,7]. However, the pleiotropic properties of quercetin warrant further study of other proteins that might also be involved in its mechanisms of action.
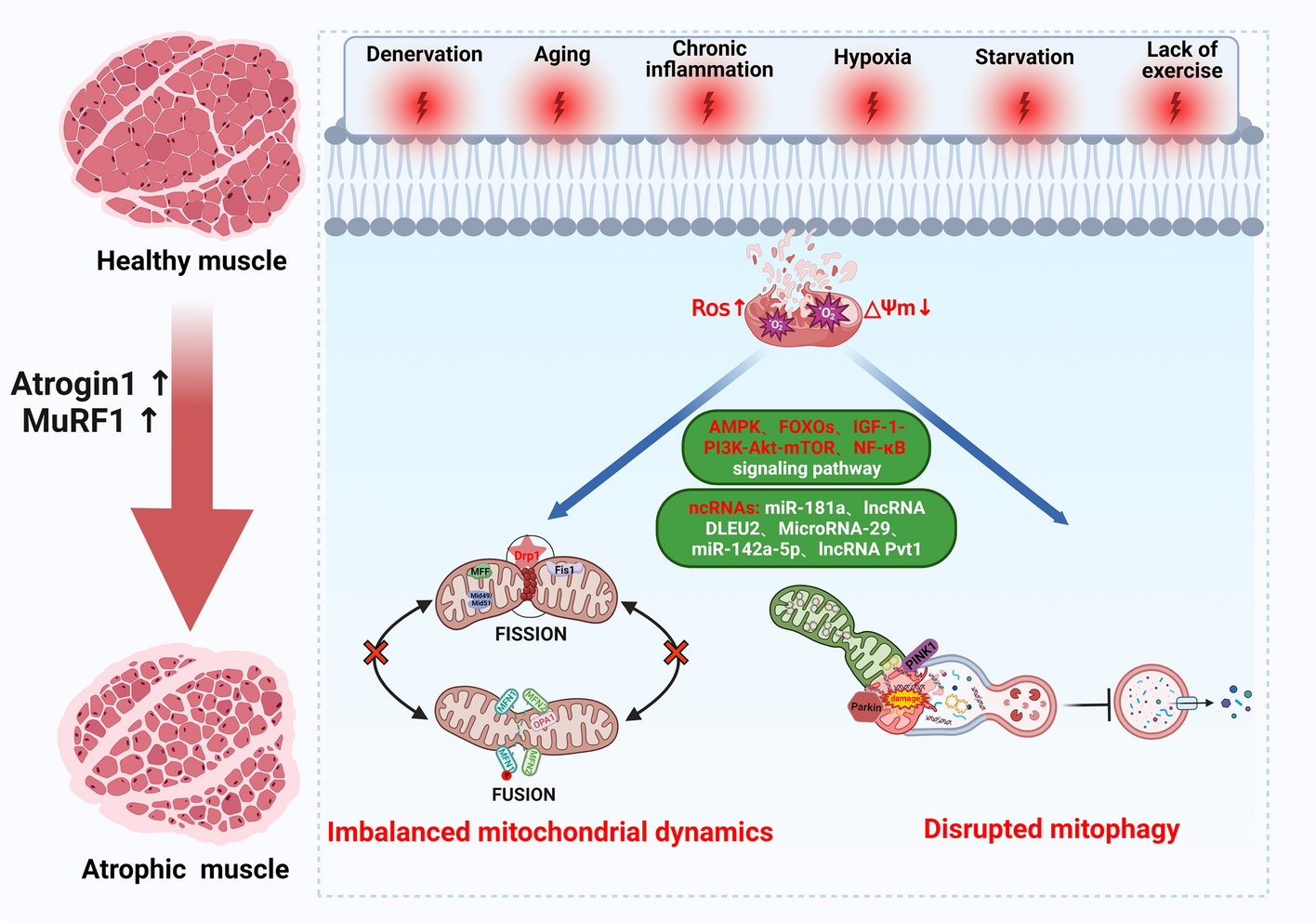
JAK–STAT signaling pathwayThe Janus kinase–signal transducer and activator of transcription (JAK–STAT) pathway exerts an important role in transducing signals from cell membrane receptors to the nucleus [8, 9]. The JAK family of tyrosine kinases bind to the cytoplasmic regions of type I and type II cytokine receptors. The binding of ligands to receptors results in the multimerization of receptors. Some receptor subunits are expressed as homodimers, such as growth hormone and erythropoietin, while others are expressed as heteromultimers such as interleukins (ILs) and interferons (IFNs). Receptors associated with JAKs are activated to start transphosphorylation of JAKs and subsequent recruitment of STATs. Human JAK family has four members: JAK1, JAK2, JAK3, and TYK2 [8, 10]. JAK family members have several distinct domains, including N-terminal FERM domain, Src homology 2 (SH2) domain, pseudokinase domain, and conserved PTK domain [8]. STAT is one of the most prominent transcriptional factor families in cancers, consisting of seven structurally similar members: STAT1, STAT2, STAT3, STAT4, STAT5a, STAT5b, and STAT6 [11, 12]. Generally, STAT family members contain six common functional domains: a coiled-coil domain (CCD), an N-terminal domain (NH2), a DNA-binding domain (DBD), an SRC homology 2 domain (SH2), a linker domain, and a transactivation domain (TAD). Aberrant activation of the JAK–STAT pathway is evident in various diseases such as cancers [13] and neurodegenerative diseases [14]. In many cancers, STAT3 is constitutively activated. There are multiple original studies on this issue investigating the role of STAT3 signaling in several solid tumors [13].
Quercetin properties: structure and biologic functionsQuercetin, also known as 2,3,4,5,7-pentahydroxyflavone or 2,3,4,5,7-dihydroxyphenyl-3,5,7-trihydroxychromen-4-one, contains two benzene rings, A and B, that are connected by a pyrone ring. Owing to the presence of two antioxidant pharmacophores in quercetin’s structure, it can effectively neutralize free radicals and join transitional metal ions. Furthermore, catechol, in conjunction with the OH group present at position C3 in the structure of quercetin, is a highly effective way to scavenge free radicals. As a pentalhydroxyl flavonol, hydroxyl groups of quercetin are positioned on the 3rd, 30th, 40th, and 7th carbon positions in the flavonol structure. Interestingly, with the replacement of diverse functional groups, quercetin becomes more biochemically and pharmacologically active [15,16,17]. Some studies suggest that quercetin is present in two forms: as a standalone compound and as an aglycone or as a mixture with other compounds. In addition to carbohydrates, lipids, alcohols, and sulfate groups, it can be reacted with other molecules to produce numerous derivatives of quercetin, including glycosides, ethers, prenylated quercetins, and sulfated derivatives [17]. Quercetin glycoside is the most abundant flavonoid compound in propolis as well as many other dietary supplements, including fruits and vegetables, such as onions, broccoli, apple, tea, and red wine [15]. Aside from acting as an anticancer, antitumor, anti-ulcer, anti-allergy, antiviral, anti-inflammatory, and antidiabetes agent, it also exerts gastroprotection, antihypertension, immune modulation, and anti-infection effects [18]. A preclinical study demonstrated that quercetin significantly reduced levels of inflammation moderators, including NO synthase, COX-2, and CRP, in a human hepatocyte-derived cell line [19]. Studies conducted on rats have shown that quercetin (80 mg equivalent dose) inhibits acute and chronic inflammation and that it also has significant anti-arthritic properties against adjuvant-induced arthritis [20, 21]. This bioactive compound has also been studied for its anticancer properties. Quercetin is believed to inhibit cancers of various solid tissues, such as breast, lung, nasopharyngeal, kidney, colorectal, prostate, pancreatic, and ovarian cancers, as well as neurodegenerative diseases [15]. In this regard, quercetin is not harmful to healthy cells, while it can impose cytotoxic effects on cancer cells through a variety of mechanisms, making it an effective supplementary agent for treating solid tumors and neurodegenerative diseases in combination with other anticancer medications [22]. Despite the range of biological benefits mentioned above, quercetin has been restricted in its pharmaceutical application as a result of its low hydrosolubility, lack of stability in physiology, high metabolism in the liver prior to reaching the bloodstream, and low bioavailability [23, 24].
In patients with cancer, quercetin was intravenously injected at a dose of 60–2000 mg/m2. The researchers determined a safe dose to be 945 mg/m2. A toxic dose resulted in vomiting, high blood pressure, nephrotoxicity, and a decrease in serum potassium. In intravenous administration, quercetin has a half-life of 0.7–7.8 min. Dispensation content is 3.7 L/m2, and elution is 0.84 L/min/m2 [25]. At a dose of 200 mg, Graefe and colleagues investigated the pharmacokinetics of quercetin. Quercetin has been reported to have a Cmax and a Tmax of 2.3–1.5 g/mL and 0.7–0.3 h, respectively [26]. According to these observations, numerous quercetin analogs and nanoconjugates of quercetin containing various delivery systems have been demonstrated, including liposomes, silver nanoparticles, poly lactic-co-glycolic acid (PLGA), and polymeric micelles, for example, DOX-conjugated micelles, metal-conjugated micelles, nucleic-acid-conjugated micelles, and antibody-conjugated micelles, which show a variety of biological implications such as biphasic, inotropic impacts [27, 28]. For instance, Iacopetta et al. synthesized a series of quercetin derivatives referred to as Q2–Q9 using catechol ketal as the starting material in which the OH groups were all or partially replaced with hydrophobic functional groups such as acetyl esters, ethyl or benzyl ethers, or diphenyl ketals of the catechol system. By inhibiting topoisomerases I and II and modulating intracellular reactive oxygen species (ROS) production, these chemical modifications produced compounds with higher anticancer activity relative to the canonical quercetin in two breast cancer models. Accordingly, quercetin exerts its antioxidant effects via competitive inhibition of the enzyme xanthine oxidase and noncompetitive inhibition of the enzyme xanthine dehydrogenase [29].
In addition, a study by Guan and colleagues prepared and evaluated quercetin nanoparticles (QPTN) as a means of improving the solubility of quercetin with PLGA and TPGS. The results demonstrated that QPTN was capable of inducing HepG2 cell apoptosis in a dose-dependent way and suppressing tumor growth by 59.07% [30].
JAK–STAT inhibitory effects of quercetin on cancerThe specific effect of quercetin on tumor cells without any impact on normal or nontransformed cells has compelled many researchers to explore its potential use as an adjuvant in suppressing proliferation, metastasis, epithelial–mesenchymal transition (EMT), and oxidative stress [31,32,33]. Having more potent antitumor activity, fewer side effects, and cost-effectiveness, natural products are promising candidates for pharmaceutical development as anticancer agents. Recently, studies reported that the expression of antiviral genes, such as 2′-5′-oligoadenylate synthetase (2′5′-OAS) and RNA-activated protein kinase (PKR), are stimulated by type-I IFNs through the JAK–STAT pathway [34,35,36]. Although the exact mechanism of action on the JAK–STAT pathway remains unclear, tofacitinib and ruxolitinib are JAK inhibitors that have been approved by the Food and Drug Administration (FDA) for their inhibitory effect on some lymphoma tumor cells. High doses of these drugs cause side effects despite their “pan-JAK” inhibitory effects, including infections and malignant tumors, gastrointestinal perforation, venous thromboembolism, dyslipidemia, and other complications [37]. Patients with rheumatoid arthritis taking tofacitinib, the first JAK inhibitor on the market, have suffered pulmonary thrombosis and death due to high doses of the drug. On the other hand, studies have shown that quercetin increased type-I IFN-induced JAK–STAT signaling through the inhibition of SHP2. It was reported that quercetin has a synergistic effect when concomitantly using some anticancer drugs [12]. According to Tiwari et al., an investigation compared quercetin and gefitinib loaded onto polyvinylpyrrolidone (PVP)-functionalized graphene oxide (GO-PVP) with quercetin and gefitinib separated by polyvinylpyrrolidone (PVP)-functionalized graphene oxide. The combined drugs loaded onto the GO-PVP nano platform were significantly more toxic than the individual drugs loaded onto the platform, as well as the free drugs, in regard to PA-1 cells [38]. Here, we described antitumor effects of quercetin by targeting JAK–STAT signaling pathways (Fig. 1).
Fig. 1
Therapeutic effects of quercetin on inhibiting the JAK–STAT signaling pathways in cancers and neurodegenerative diseases
Moreover, we suggest that more investigations based on natural products are needed to provide a promising therapeutic approach to enhance the efficacy of anticancer therapy (e.g., chemotherapy, radiotherapy, or radiochemotherapy) in a less toxic manner, also before or after tumor surgery.
Glioblastoma multiformeGlioblastoma multiforme (GBM) is a malignant tumor of connective tissue, and has been known as the most deadly primary brain tumor [1, 39,40,41]. Flavonoids such as quercetin are able to cross the blood–brain barrier (BBB) via different mechanisms, including carrier-mediated transcellular transport transcellular diffusion or paracellular distribution via tight junctions between BBB endothelial cells [1, 42]. In gliomagenesis, STAT3 may act as an oncogene [43]. Consequently, inflammatory microenvironments may favor tumor development [44]. A potential role for quercetin in the prevention and treatment of glioblastoma has been demonstrated as a suppressor of the STAT3 activation signaling pathway stimulated by IL-6 [44, 45]. In this line, IL-6-induced JAK–STAT3, Rb phosphorylation, cyclin D1 expression, and MMP-2 secretion are inhibited by 48 h treatment with 25 µM quercetin in T98G and U87 GBM cell lines [44].
On the other hand, mouse peritoneal macrophages were isolated and quercetin was used to treat the ex vivo inflammation induced by lipopolysaccharide in a study by Liao et al. There was a significant decrease in TNF-αTNF-α gene expression levels as well as pro-/anti-inflammatory ratios (TNF-α/IL-10) and inactivation of inflammatory macrophages, but IL-10 gene expression levels were elevated in quercetin-treated macrophages. This suggests that quercetin treatment has overall anti-inflammatory potential. In the activated inflammatory macrophages, quercetin inhibited TLR2 gene expression as well as STAT3 protein phosphorylation [46]. Moreover, as a potential prognostic marker for glioblastoma multiforme (GBM), Axl plays a crucial role. Signals downstream of it include those from NF-κB, JAK–STAT, and PI3K–Akt–mTOR [47]. With 0, 25, 50, and 100 µM doses of quercetin, dose-dependent apoptosis was observed in two GBM cell lines, U87MG and U373MG, resulting from decreased Axl protein expression, IL-6 expression levels, and STAT3 phosphorylation [48]. Furthermore, it was found that quercetin inhibited glioblastoma cell invasion and angiogenesis in vitro. It is possible that quercetin inhibits migration and angiogenesis via downregulating the levels of VEGFA, MMP9, and MMP2 [49].
Moreover, a previously conducted study synthesized an anticancer drug commonly used in treating glioblastoma that combined nanoquercetin (nanohydrogels) with temozolomide to enhance its specificity and efficacy. As a codelivery vehicle for temozolomide and quercetin, the nanohydrogel facilitated the internalization and cytotoxicity of quercetin. In addition, using quercetin-containing nanohydrogels significantly reduced IL-8 production, IL-6 production, and VEGF production in a dose-dependent manner in pro-inflammatory conditions, with meaningful implications for glioblastoma cells’ drug resistance. However, it was used at 100 and 200 µg doses [50].
Recent evidence suggests that supplementation of standard therapy with quercetin enhances the efficacy of treatment of experimental glioblastoma by inducing apoptosis through caspase-3 and PARP-1 cleavage, and suppressing PI3K–Akt activation. When combining quercetin and irradiation, the breakdown of caspase-3, PARP-1, phosphorylated ERK, JNK, p38, and RAF1 were increased, whereas phosphorylated Akt was considerably decreased. By enhancing apoptosis through inducing caspase-3 and PARP-1 cleavage and suppressing activation of the Akt pathway, quercetin supplements standard therapy with improved efficacy in treating experimental glioblastoma [51].
Quercetin also induces apoptosis in human glioblastoma multiform T98G cells, which is associated with activation of caspase 3 and 9 and release of cytochrome C from the mitochondria. It has been suggested that apoptosis is initiated by stress in the endoplasmic reticulum by increasing expression of caspase 12 and the presence of multiple granules in the cytoplasm upon temozolomide treatment with or without quercetin [52].
Hepatocellular carcinomaHepatocellular carcinoma (HCC), one of the most common cancers worldwide, is the second and sixth leading cause of cancer-related death in men and women, respectively [53]. It is estimated that 600,000–800,000 new cases of HCC occur annually on a global level [54]. Various intracellular signaling pathways activation implicated in cell growth, survival, apoptosis, angiogenesis, and differentiation have been found to facilitate HCC development and progression. These oncogenic signaling pathways include JAK–STAT, PI3K–Akt–mTOR, Wnt–β-catenin, and Ras–Raf–MAPK pathways [55]. According to recent investigations, the signal transducer and activator of transcription protein 1 (STAT1) was phosphorylated by quercetin, promoting endogenous IFN-α-regulated gene expression. Additionally, quercetin reduced the proliferation rate of hepatocellular carcinoma HepG2 and Huh7 cells when IFN-α expression was stimulated [6]. In the development of HCC, STAT3 is generally accepted as an oncogene. Activation of STAT3 stimulates the expression of several genes that play a significant role in cancer development, emphasizing the importance of STAT3 in HCC [54]. Wu et al.’s study revealed that quercetin suppresses hepatocellular carcinoma progression via modulating cellular invasion, migration, and autophagy. Moreover, its effect may partly be attributed to the downregulation of the JAK2–STAT3 signaling pathway [56]. Also, quercetin plays a suppressive role against HCC cells through apoptosis and p16-mediated cell cycle arrest, and its combination with cisplatin displayed synergistic inhibitory activity in triggering apoptosis and suppressing cancer cell growth [57]. A microprecipitation high-pressure homogenization technique, with methyl polyethylene glycol-Deoxycholic acid (mPEG-DCA) as a stabilizer, was used to synthesize three quercetin nanosuspensions containing different particle sizes. All three quercetin nanoparticles (QUR-NPs) were found to have significantly stronger inhibitory activity against MCF-7 cells and hepatocyte 7702 cells in vitro, while inhibiting tumors against the murine hepatic carcinoma H22 model in vivo [58].
The effects of quercetin on in vivo and in vitro hepatocellular carcinoma progression were also examined in a study using LM3 and nude mouse tumor models, in which quercetin suppressed cell apoptosis, migration, invasion, and autophagy by inhibiting the JAK2–STAT3 signaling pathway [56].
Gastric cancerDespite declines in incidence rates, increased awareness, and advances in treatment strategies, gastric cancer remains one of the most prevalent forms of cancer worldwide [59]. The development and progression of gastric cancer are influenced by Helicobacter pylori infection, lifestyle factors, dietary factors, and various genetic aberrations [60, 61]. Upon entry into human gastric epithelial cells, the CagA protein of H. pylori may exhibit tyrosine phosphorylation with downstream effects on signal transduction. Researchers have found that the CagA phosphorylation status affects the signal switch between the JAK–STAT3 and SHP2–ERK pathways via gp130, elucidating a novel mechanism for the H. pylori signaling pathway [62, 63].
Furthermore, González-Segovia et al. found that oral quercetin administration reduced H. pylori infection and both inflammatory response and lipid peroxidation in vivo [64]. Quercetin inhibits the proliferation of human gastric cancer (GC) MGC-803 cells. This may be related to the downregulation of leptin and leptin receptor protein, leptin mRNA, and leptin receptor mRNA expression by the JAK–STAT pathway [65]. Also, combined treatment with quercetin and curcumin was observed with substantial inhibition of cell proliferation, associated with loss of mitochondrial membrane potential (ΔΨm), cytochrome C release, and decreased ERK and AKT phosphorylation. These findings suggest that the combination of quercetin and curcumin leads to apoptosis through the mitochondrial pathway [66].
Breast cancerWomen worldwide are most likely to develop breast cancer with a high rate of mortality [67, 68]. Despite improvements in chemotherapeutic agents, new strategies to overcome tumor cell survival remain elusive despite progress in understanding the mechanisms of chemoresistance [69, 70].
To create an ideal cancer treatment protocol, we need to identify key target molecules, as well as safe and stable delivery systems that will allow us to overcome resistance and minimize the side effects of chemotherapeutic agents [71, 72].
In this regard, Safi et al. indicated that concomitantly using docetaxel and quercetin synergistically results in inhibition of both cell growth and cell survival, as well as apoptosis induction in MDA-MB-231 human breast cancer cell line [12]. The results showed that single-agent treatment with docetaxel or quercetin leads to a decrease in the viability of the MDA-MB-231 cells at 48 h, whereas the combination of docetaxel (7 nM) and quercetin (95 μM) displayed the greatest synergistic effects with a combination index value of 0.76 accompanied by enhancing expression of the p53 gene as well as pro-apoptotic protein BAX, and reducing the expression of STAT3 anti-apoptotic protein BCL2, phosphorylated AKT, and ERK1/2 [12]. Further, quercetin has been revealed to have a role in antiproliferation of HER2-overexpressing BT-474 BC cells via STAT3 signaling suppression and caspase-dependent extrinsic apoptosis activation, while causing an increase in sub-G0/G1 apoptotic population [73].
Pancreatic cancerPancreatic cancer (PC), a highly fatal disease, is one of the most commonly occurring digestive malignancies. PC is one of the leading causes of cancer-related death worldwide [53, 74, 75]. Moreover, patients with PC have a 5-year survival rate of 21.3% for local-stage cancer, 8.9% for regional-stage cancer, and 1.8% for distant-stage cancer [76].
According to Yu et al., quercetin inhibited PATU-8988 and PANC-1 cell growth and reduced MMP release. In this study, they examined whether quercetin treatment affected malignancy in cells by activating STAT3 and IL-6. They indicated that, when MMP secretion and EMT occur, STAT-3 signaling is stimulated, and quercetin reverses IL-6-induced EMT and invasion. It has been revealed that quercetin can inhibit EMT, invasion, and metastasis, as well as r
留言 (0)