
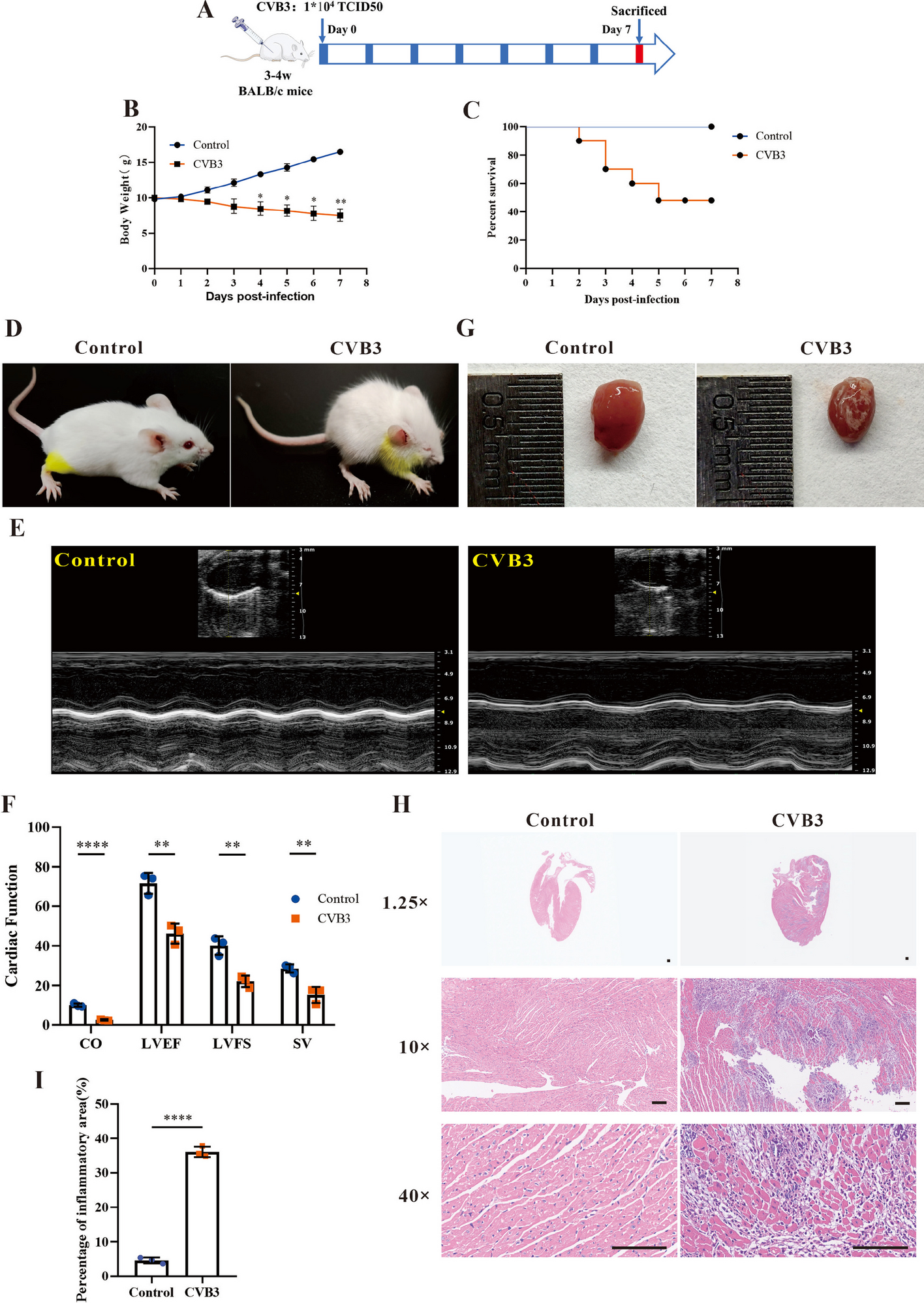
記住我


To reveal the relationship between the stemness of GCSCs and m6A modification of lncRNAs, the m6A-modified lncRNAs of GCSCs sorted from the solid tumors of a GC patient were characterized. The results indicated a tumorsphere formation rate of 97.9% for the identified potential GCSCs, whereas no tumorspheres were observed in the case of the GCNSCs (Fig. 1A). Correspondingly, a single cell from a tumorsphere of potential GCSCs had the capability to form another tumorsphere (Fig. 1B). Western blot analysis demonstrated a notable upregulation of some stemness genes in GCSCs (Fig. 1C). The migration efficiency and the chemoresistance capacity of GCSCs were significantly increased compared with those of GCNSCs (Fig. 1D and E). These data confirmed a successful isolation of GCSCs from solid tumors.
Fig. 1
N6-methyladenosine lncRNAs in GCSCs. A Percentage of tumorsphere formation of GCSCs and GCNSCs. B Tumorsphere formation assay of a single cell from GCSCs; C detection of the expressions of stemness genes in GCSCs and GCNSCs with Western blot. D Efficiency of the migration of GCSCs. GCSCs and GCNSCs were subjected to Transwell migration assays to examine the cell migration. The representative images of the migrated cells are shown on the left. The percentage of the migrated cells is indicated on the right (**, p < 0.01). Scale bar, 50 μm. (E) Chemoresistance capacity of GCSCs. GCSCs or GCNSCs were treated with different concentrations of cisplatin. At 48 h after treatment, the cells were subjected to cell viability assays (**, p < 0.01). The half maximal inhibitory concentration (IC50) values were calculated. F The m6A modification of RNAs in GCSCs and GNCSCs. G Heatmap of the differentially expressed m6A-modified lncRNAs in GCSCs and GNCSCs. H Expression profiles of methylated lncRNAs in GCNSCs and GCSCs. **, p < 0.01
The results of dot blot using m6A-specific antibody revealed a considerably elevated m6A level in RNAs within GCSCs compared with GCNSCs (Fig. 1F), suggesting that the m6A modification of RNAs play a crucial role in GCSCs. Therefore, the m6A-modified lncRNAs of GCSCs were sequenced. On the basis of MeRIP-seq, the m6A level of 128 lncRNAs were elevated in GCSCs, yet the m6A level of 1 lncRNA was decreased in GCSCs (Fig. 1G). The MeRIP-seq data showed that the m6A levels of six lncRNAs (PAPPA-AS1, PSMA3-AS1, MIR22HG, LINC00342, LINC01410, and LINC00680) were significantly upregulated in GCSCs compared with those in GCNSCs (Fig. 1G). To confirm the expression profiles of the methylated lncRNAs in GCSCs, the m6A levels of these six lncRNAs were analyzed by MeRIP-qPCR. The results indicated that the six lncRNAs were significantly upregulated in GCSCs compared with the control (Fig. 1H), confirming the MeRIP-seq data.
Collectively, the findings unveiled a substantial increase in m6A levels of lncRNAs within GCSCs compared with those in GCNSCs, implying the essential roles of m6A modification of lncRNAs in GCSCs.
Mechanism of lncRNA methylation in GCSCsTo elucidate the function of methylated lncRNAs in GCSCs, the GCSCs were further sorted from HGC-27 and MGC-803. Using flow cytometry, we sorted ALDH1-positive cells from HGC-27 and MGC-803, and the tumorsphere formation assay demonstrated that a tumorsphere could be formed by a single cell derived from the sorted ALDH1-positive cells of HGC-27 or MGC-803 (Fig. 2A). Meanwhile, Western blot analysis showed that the genes related to stemness were upgraded in the ALDH1-positive cells (Fig. 2B). The capacities of migration and chemoresistance of the ALDH1-positive cells were significantly increased compared with those of the control (Fig. 2C, D). These results showed a successful isolation of GCSCs from HGC-27 and MGC-803.
Fig. 2

Mechanism of lncRNA methylation in GCSCs. A Tumorsphere formation assay of a single cell from the GCSCs sorted from HGC-27 (GCSC-HGC) and MGC-803 (GCSC-MGC). B Western blot analysis of the stemness genes in GCSC-HGC and GCSC-MGC. C Migration capacity of GCSCs (GCSC-HGC and GCSC-MGC). GCSC-HGC, GCNSC-HGC, GCSC-MGC, and GCNSC-MGC were analyzed with Transwell migration assays to examine the cell migration. The representative images of the migrated cells are indicated on the left. The percentage of the migrated cells is shown on the right (**, p < 0.01). Scale bar, 50 μm. D Efficiency of chemoresistance of GCSCs (GCSC-HGC and GCSC-MGC). GCSCs or GCNSCs were treated with cisplatin at different concentrations. At 48 h after treatment, the cells were characterized using cell viability assays (**, p < 0.01). The half maximal inhibitory concentration (IC50) values were calculated. E Examination of the expressions of methyltransferases METTL3 and METTL14 in GCSCs. F Western blot for METTL3 in GCSCs with METTL3-shRNA or METTL3-shRNA rescue. G m6A level of lncRNAs in GCSC with METTL3-shRNA or METTL3-shRNA rescue. H Detection of the expressions of stemness genes in the METTL3-silenced or rescued GCSCs using Western blot. β-tubulin was used as a control. I Western blot for METTL14 in GCSCs with METTL14-shRNA or METTL14-shRNA rescue. J m6A level of lncRNAs in GCSC with METTL14-shRNA or METTL14-shRNA rescue. K Detection of the expressions of stemness genes in METTL14-silenced or rescued GCSCs by Western blot. **, p < 0.01
To reveal the mechanism of lncRNA methylation in GCSCs, the enzymes responsible for lncRNA methylation, including METTL3 and METTL14, the key methyltransferases for the m6A modification of mRNAs and lncRNAs, were characterized in GCSCs, GCSC-HGC, and GCSC-MGC. Western blot analysis revealed a substantial upregulation of METTL3 and METTL14 in three distinct GCSC types (Fig. 2E), suggesting the significant roles of METTL3 and METTL14 in the GCSCs.
To investigate the impact of METTL3/METTL14 on lncRNA methylation in GCSCs, sequence-specific shRNA (METTL3/METTL14-shRNA) was employed to knock down METTL3/METTL14 in three GCSC types. Additionally, the silenced METTL3/METTL14 was rescued (METTL3/METTL14-shRNA rescue) in the METTL3/METTL14-silenced GCSCs, and subsequent examination focused on lncRNA methylation. The results of western blot demonstrated the successful silencing or rescue of METTL3/METTL14 in all three GCSC types (Fig. 2F, I). The silence of METTL3/METTL14 caused a reduction of m6A level of lncRNAs, including MIR22HG, LINC00342, PSMA3-AS1, LINC01410, PAPPA-AS1, and LINC00680, in GCSCs, while METTL3/METTL14-shRNA rescue repaired the discrepancy of m6A level in lncRNAs (Fig. 2G, J).
To evaluate the impact of METTL3/METTL14 on GCSCs, the protein levels of stemness genes were assessed in GCSCs with METTL3/METTL14-shRNA or METTL3/METTL14-shRNA rescue by western blot analysis. The findings demonstrated a significant reduction in protein levels of those genes in GCSCs with METTL3/METTL14-shRNA, whereas the expression levels of those genes in the METTL3/METTL14-shRNA rescued GCSCs were similar to those of the controls (Fig. 2H, K). These findings suggested that METTL3/METTL14 enhanced the stemness of GCSCs.
In summary, METTL3 and METTL14 have demonstrated their crucial roles in lncRNA methylation of GCSCs, which could promote the stemness of GCSCs.
Roles of methylated lncRNAs in GCSCsOn the basis of the MeRIP-Seq data, six lncRNAs (PAPPA-AS1, PSMA3-AS1, MIR22HG, LINC00342, LINC01410, and LINC00680) had a high m6A level in GCSCs. To further elucidate the functions of methylated lncRNAs in GCSCs, the expression profiles of six lncRNAs were investigated. The results showed that two lncRNAs (PSMA3-AS1 and MIR22HG) were substantially elevated in all GCSCs compared with the corresponding GCNSCs (Fig. 3A), suggesting the crucial roles played by these lncRNAs in GCSCs.
Fig. 3


Roles of methylated lncRNAs in GCSCs. A Expression profiles of six lncRNAs (PAPPA-AS1, PSMA3-AS1, MIR22HG, LINC00342, LINC01410, and LINC00680) in GCSCs. B Validation of silencing of lncRNAs in GCSCs. C Detection of the expressions of stemness genes in GCSCs with lncRNAs-siRNA. D Cell viability for PSMA3-AS1 or MIR22HG silencing GCSCs. E Cell cycle analysis of PSMA3-AS1 or MIR22HG-silenced GCSCs. F Caspase-3/7 assay for PSMA3-AS1 or MIR22HG silencing GCSCs. G Prediction of the methylation sites of lncRNAs. H The m6A levels of lncRNAs between GCSCs and GCNSCs. I The m6A levels of METTL3-silenced GCSCs with site-specific methylation of lncRNAs. J Cell viability for METTL3-silenced GCSCs with site-specific methylation of lncRNAs. K Cell cycle analysis of METTL3-silenced GCSCs with site-specific methylation of lncRNAs. L Caspase-3/7 assay for METTL3-silenced GCSCs with site-specific methylation of lncRNAs. M Western blot for stemness genes of METTL3-silenced GCSCs with site-specific methylation of lncRNAs. N The percentage of tumorsphere formation of METTL3-silenced GCSCs with site-specific methylation of lncRNAs. O Tumorsphere formation assay of a single cell from METTL3-silenced GCSCs with methylated lncRNAs. **, p < 0.01
To evaluate the impact of lncRNAs on GCSCs, PSMA3-AS1 or MIR22HG was knocked down in GCSCs, followed by examining the stemness of knockdown GCSCs. The RT-qPCR data demonstrated that PSMA3-AS1 and MIR22HG were respectively silenced in three types of GCSCs (Fig. 3B). The silencing of PSMA3-AS1 or MIR22HG caused a significant downregulation of the stemness markers in GCSCs (Fig. 3C), showing the important roles of PSMA3-AS1 or MIR22HG in GCSCs. And the results of the cell viability assay showed that the PSMA3-AS1 or MIR22HG downregulation significantly reduced the viability of GCSCs (Fig. 3D). The suppression of GCSCs’ viability mediated by the PSMA3-AS1 or MIR22HG silencing resulted from cell cycle arrest in the G0/G1 phase, subsequently leading to apoptosis of GCSCs (Fig. 3E, F). These findings indicated that PSMA3-AS1 and MIR22HG exert positive influences on GCSCs.
To investigate the impact of m6A modification on lncRNAs in GCSCs, we initially predicted the m6A sites within the lncRNAs PSMA3-AS1 and MIR22HG by SRAMP (http://www.cuilab.cn/sramp). The analysis revealed the presence of 12 potential m6A sites in PSMA3-AS1 and 10 in MIR22HG (Fig. 3G). According a previous report, single-base elongation- and ligation-based qPCR amplification analysis (SELECT) was performed to determine the m6A sites of lncRNAs [27].The data of qPCR analysis confirmed that 11 and 8 of the predicted m6A sites of PSMA3-AS1 and MIR22HG were methylated in GCSCs, respectively (Additional file 1: Fig. S1A). Among the m6A sites of lncRNAs, the N6-methyladenosine modification level at position 1225 of PSMA3-AS1 (m6A1225) or at position 2041 of MIR22HG (m6A2041) was significantly increased in GCSCs compared with GCNSCs (Fig. 3H), implying that the site-specific methylated adenosine of PSMA3-AS1 and MIR22HG play a significant role in GCSCs.
To elucidate the impact of the site-specific m6A modification of lncRNAs on GCSCs, the A1225 site of PSMA3-AS1 and the A2041 site of MIR22HG were specifically methylated in the METTL3-silenced GCSCs and then the methylation on those lncRNAs were confirmed (Fig. 3I). Meanwhile, the results of the cell viability assay indicated that silencing of METTL3 obviously impaired the viability of GCSCs, while the methylation of A1225 of PSMA3-AS1 (METTL3-shRNA + methylated PSMA3-AS1) or A2041 of MIR22HG (METTL3-shRNA + methylated MIR22HG) significantly increased the viability of METTL3-silenced GCSCs compared with METTL3-silenced GCSCs (METTL3-shRNA) (Fig. 3J), implying that the site-specific m6A modification of lncRNAs could promote the proliferation of GCSCs. The data of cell cycle analysis revealed that GCSCs with METTL3-shRNA induced cell cycle arrest in G1/G0 phase, and the methylation of A1225 of PSMA3-AS1 or A2041 of MIR22HG recovered the cell cycle of GCSCs (Fig. 3K). Correspondingly, the METTL3 knockdown-mediated cell cycle arrest activated apoptosis of GCSCs, whereas the apoptotic activity of METTL3-silenced GCSCs with the methylation of A1225 of PSMA3-AS1 (METTL3-shRNA + methylated PSMA3-AS1) or A2041 of MIR22HG (METTL3-shRNA + methylated MIR22HG) was commensurate with that of the control (METTL3-shRNA-scrambled; Fig. 3L). Evidently, the site-specific m6A modification of lncRNAs was required for the proliferation of GCSCs.
Furthermore, the western blots analysis revealed a significant elevation of stemness in METTL3-silenced GCSCs with methylation of the A1225 site of PSMA3-AS1 or the A2041 site of MIR22HG (Fig 3M). Consistently, the methylation of PSMA3-AS1 or MIR22HG significantly promoted the tumorsphere formation of the METTL3-silenced GCSCs (Fig 3N and O). These data unveiled a positive contribution of lncRNA methylation to stemness of GCSCs.
Together, these data demonstrated that site-specific m6A modification of lncRNAs (PSMA3-AS1 and MIR22HG) was required for the proliferation of GCSCs via suppressing apoptosis and promoting the stemness of GCSCs.
Influence of m6A modification on the stability of lncRNAsTo investigate the effect of m6A modification on lncRNA stability, the RNA levels of PSMA3-AS1 and MIR22HG in METTL3-silenced GCSCs were determined. A considerable decrease of the RNA levels of PSMA3-AS1 and MIR22HG were exhibited in METTL3-silenced GCSCs (Fig. 4A), intimating that m6A modification could enhance the stability of lncRNAs.
Fig. 4
Influence of m6A modification on the stability of lncRNAs. A Relative expression level of lncRNAs PSMA3-AS1 and MIR22HG in METTL3-silenced GCSCs. B Examination of lncRNAs stability in METTL3-silenced GCSCs by treating with actinomycin D. **, p < 0.01
To further validate the influence of m6A modification on lncRNAs stability, the METTL3-silenced GCSCs were exposed to actinomycin D, followed by the assessment of lncRNAs enrichment. The findings indicated elevated expression levels of PSMA3-AS1 and MIR22HG in METTL3-silenced GCSCs treated with actinomycin D than those in GCSCs (Fig. 4B). It can be inferred that m6A modification could stabilize PSMA3-AS1 and MIR22HG in GCSCs.
Underlying mechanism of lncRNAs in GCSCsGenerally, lncRNAs function by binding to miRNAs. On the basis of literature and prediction using ENCORI and DIANA-LncBase, the potential miRNAs targeted by PSMA3-AS1 or MIR22HG were obtained (Additional file 1: Table S1). The results of lncRNA pulldown assays showed that miR-101, miR-4429, and miR-411-3p were notably enriched in the pulled down RNAs using PSMA3-AS1, while miR-24-3p was significantly enriched in the pulled down RNAs using MIR22HG (Fig. 5A). Thereafter, the dual luciferase reporter assays revealed that miR-411-3p or miR-24-3p significantly reduced the luciferase activity of GCSC cotransfected with miR-411-3p and pmirGLO-PSMA3-AS1 or miR-24-3p and pmirGLO-MIR22HG (Fig. 5B), suggesting the direct interaction between miR-411-3p and PSMA3-AS1, in a similar manner to miR-24-3p and MIR22HG.
Fig. 5

Underlying mechanism of lncRNAs in GCSCs. A The relative enrichment of miRNAs interacted with lncRNAs in GCSCs. B Dual-luciferase reporter assay for potential miRNAs-lncRNAs interaction in GCSCs. C The expression profiles of miR-411-3p and miR-24-3p in GCSCs and GCNSCs. D Prediction of potential target genes of miRNAs using microT, miRmap, and TargetScan. E Validation for overexpression of miRNAs in GCSCs. F Detection of the expression levels of target genes in the miRNA-overexpressed GCSCs. G Validation of interaction between miRNAs and target genes by dual-luciferase reporter assay. H The expression levels of target genes of miRNAs in GCSCs and GCNSCs. I Western blot for target genes of miRNAs in GCSCs and GCNSCs. J Caspase-3/7 assay for AP1G1 or SERTAD1 silencing GCSCs. K Western blot for stemness genes in AP1G1 or SERTAD1 silencing GCSCs. L The expression level of AP1G1 or SERTAD1 in PSMA3-AS1 or MIR22HG silencing GCSCs. M Western blot analysis for target genes of miRNAs in PSMA3-AS1 or MIR22HG-silenced GCSCs. **, p < 0.01
To elucidate the roles of miR-411-3p and miR-24-3p in GCSCs, we examined the expression profiles of these miRNAs in both GCSCs and GCNSCs. The findings indicated an obvious upregulation of miR-411-3p and miR-24-3p in GCSCs (Fig. 5C), suggesting that these two miRNAs played important roles in GCSCs.
To further explore the roles of the miRNAs interacting with lncRNAs, the potential targets of miRNAs were predicted using microT, miRmap, and TargetScan algorithms. The results showed that four genes were the potential targets of miR-411-3p, while ten genes might be targeted by miR-24-3p (Fig. 5D). To confirm the target genes of miRNAs, the change of expression of target genes were examined after transfecting with miR-411-3p or miR-24-3p in GCSC. Upon overexpression of miR-411-3p or miR-24-3p in GCSC (Fig. 5E), the expression level of AP1G1 or TRPC4AP and SERTAD1 was substantially diminished (Fig. 5F), implying that miR-411-3p or miR-24-3p might target AP1G1 or TRPC4AP and SERTAD1. And the dual luciferase assays were performed and the data demonstrated that AP1G1 or SERTAD1 was the gene targeted by miR-411-3p or miR-24-3p (Fig. 5G).
To explore whether the target genes of miRNAs had effects on GCSCs, the expression levels of AP1G1 and SERTAD1 in GCSCs and GCNSCs were assessed. The findings indicated a remarkable upregulation of AP1G1 and SERTAD1 in GCSCs (Fig. 5H and I), suggesting that AP1G1 and SERTAD1 might play essential roles in GCSCs. The AP1G1 or SERTAD1 silencing induced apoptosis of GCSC (Fig. 5J) and downregulated the protein abundance of stemness genes in GCSC (Fig. 5K), showing that AP1G1 and SERTAD1 were required for shaping GCSC characteristics. As reported, AP1G1 and SERTAD1 promote gastric cancer progression [28, 29]. In this context, AP1G1 and SERTAD1 could promote tumorigenesis of GCSCs.
The m6A sites of lncRNAs PSMA3-AS1 and MIR22HG were far from the base sequences bound to the seed sequences of miRNAs. Thus, the m6A modification of lncRNAs did not affect the interaction between PSMA3-AS1 and miR-411-3p, similarly to MIR22HG and miR-24-3p. To assess whether PSMA3-AS1 or MIR22HG functioned as a ceRNA, the expression profiles of AP1G1 or SERTAD1 were determined in the PSMA3-AS1 or MIR22HG-silenced GCSCs. The findings revealed that the PSMA3-AS1 or MIR22HG silencing noticeably decreased the expression level of miRNA-targeted mRNA of AP1G1 or SERTAD1 (Fig. 5L and M), indicating that lncRNAs PSMA3-AS1 and MIR22HG functioned as ceRNAs in GCSCs.
Collectively, these findings demonstrated that lncRNA PSMA3-AS1 or MIR22HG promoted the stemness of GCSCs via the PSMA3-AS1–miR-411-3p–AP1G1 or MIR22HG– and miR-24-3p–SERTAD1 axis.
Interactions between m6A-modified lncRNAs and proteinsTo reveal the proteins that interacted with lncRNAs, lncRNA pulldown assays were performed. The results exhibited that a specific protein from GCSCs, identified to be EFF1A1 or LRPPRC, was bound to PSMA3-AS1 or MIR22HG (Fig. 6A). However, the protein level from the METTL3-silenced GCSCs was lower than that from GCSCs (Fig. 1A), connoting that the methylation of PSMA3-AS1 or MIR22HG affected the EFF1A1–PSMA3-AS1 and LRPPRC–MIR22HG interactions. Western blots confirmed the interaction between EFF1A1 and PSMA3-AS1 or LRPPRC and MIR22HG and the lncRNA–protein interaction was weakened by lncRNA demethylation (Fig. 6B). These data indicated that lncRNA methylation promoted the interaction between lncRNA and protein.
Fig. 6
Interactions between m6A-modified lncRNAs and proteins. A SDS-PAGE for lncRNAs pulldown assay. The arrows indicate the proteins identified. M, protein marker. B Western blot analysis of the proteins bound to lncRNAs. C Detection of the stability of proteins bound to lncRNAs in PSMA3-AS1 or MIR22HG silencing GCSCs. D The levels of mRNAs and proteins of EEF1A1 and LRPPRC in GCSCs. E Validation of silencing of EEF1A1 or LRPPRC in GCSCs by RT-qPCR and Western blot assay. F Cell viability for EEF1A1 or LRPPRC silencing GCSCs. G Cell cycle analysis of EEF1A1 or LRPPRC silencing GCSCs. H Caspase-3/7 assay of EEF1A1 or LRPPRC silencing GCSCs. I Western blot for stemness genes of EEF1A1 or LRPPRC silencing GCSCs. **, p < 0.01
To unveil the impact of lncRNA methylation on the proteins bound to lncRNAs, protein stability was detected and the findings showed that PSMA3-AS1 or MIR22HG silencing decreased the EFF1A1 or LRPPRC protein level in GCSCs compared with the controls (Fig. 6C). However, PSMA3-AS1 or MIR22HG silencing had no impact on the EFF1A1 or LRPPRC mRNA level (Fig. 6C). These data demonstrated that the methylation of lncRNAs enhanced the stability of the proteins bound to PSMA3-AS1 or MIR22HG.
To investigate the roles of EEF1A1 and LRPPRC in GCSCs, the differential expression of EEF1A1 or LRPPRC in GCSCs and GCNSCs was evaluated. The results showed that at both RNA and protein levels, abundance of EEF1A1 and LRPPRC were considerably elevated in GCSCs (Fig. 6D), suggesting that EEF1A1 and LRPPRC played important roles in GCSCs. To reveal the function of EEF1A1 and LRPPRC in GCSCs, EEF1A1 and LRPPRC were knocked down in GCSCs (Fig. 6E). EEF1A1 or LRPPRC silencing led to the inhibition of GCSCs’ viability (Fig. 6F) and caused cell cycle arrest in G0/G1 phase of GCSCs, which triggered apoptosis of GCSCs (Fig. 6G, H). Consistently, the expression of stemness genes were impaired in EEF1A1-silenced or LRPPRC-silenced GCSCs (Fig. 6I). These data revealed the influential role of EEF1A1 and LRPPRC in maintaining stemness and proliferation of GCSCs.
Taken together, these results implied that the methylated PSMA3-AS1 or MIR22HG was interacted with EFF1A1 or LRPPRC protein to enhance the protein stability, leading to the suppression of apoptosis and the promotion of stemness of GCSCs.
Influence of m6A modification of lncRNAs on tumorigenesis of GCSCs in vivoTo investigate the influence of methylated PSMA3-AS1 and MIR22HG on tumorigenesis of GCSCs in vivo, METTL3-silenced GCSC with or without the m6A modification of PSMA3-AS1 or MIR22HG were injected into BALB/c mice (Fig. 7A). The tumor volume exhibited a more rapid growth in the mice injected with METTL3-silenced GCSCs containing the methylated PSMA3-AS1 or MIR22HG than mice without the lncRNA methylation (Fig. 7B). Uniformly, much bigger and heavier tumors were developed in the mice with methylated PSMA3-AS1 or MIR22HG (Fig. 7C and D). Thus, it can be inferred that m6A modification of PSMA3-AS1 or MIR22HG prominently contributes to the promotion of tumorigenesis in GCSCs in vivo.
Fig. 7
Influence of m6A modification of lncRNAs on tumorigenesis of GCSCs in vivo. A Schematic diagram for model mice with lncRNA-methylated GCSCs. B The tumor volume of the xenograft mice injected with METTL3-silenced GCSCs alone (control) or METTL3-silenced GCSCs with methylated PSMA3-AS1 or MIR22HG. C The size of solid tumors of the mice with METTL3-silenced GCSCs alone (control) or METTL3-silenced GCSCs with methylated PSMA3-AS1 or MIR22HG. D The weight of solid tumors of mice with METTL3-silenced GCSCs alone (control) or METTL3-silenced GCSCs with methylated PSMA3-AS1 or MIR22HG. E m6A levels of lncRNAs in the solid tumors of the mice injected with METTL3-silenced GCSCs alone (control) or METTL3-silenced GCSCs with methylated PSMA3-AS1 or MIR22HG. F Immunohistochemical analysis of the expressions of Ki67 and Caspase 3 in the solid tumors of the mice injected with METTL3-silenced GCSCs alone (control) or METTL3-silenced GCSCs with methylated PSMA3-AS1 or MIR22HG. G Detection of the expressions of stemness genes in solid tumors of the mice treated with the methylated PSMA3-AS1 or MIR22HG using Western blot. H Model for the roles of the methylated PSMA3-AS1 and MIR22HG in GCSCs. **, p < 0.01
Then m6A level of lncRNAs were detected for confirming the methylation of lncRNAs in solid tumors (Fig. 7E). To assess the function of lncRNA methylation on solid tumors, the apoptosis and proliferation of solid tumors were examined using immunohistochemical analysis. The immunohistochemical analysis data exhibited that Caspase 3 was dramatically reduced in the solid tumors of mice with methylated PSMA3-AS1 or MIR22HG, whereas Ki67 was upregulated (Fig. 7F), implying that the methylation of lncRNAs promoted tumor growth by suppressing apoptosis in vivo. Furthermore, the expression of stemness genes in the tumors of mice with the treatment of methylated PSMA3-AS1 or MIR22HG were upregulated (Fig. 7G). These results illustrated the positive contribution of lncRNA methylation to tumor progression in vivo.
In conclusion, these results underscore the potential of m6A modification in lncRNAs (PSMA3-AS1 and MIR22HG) to drive tumorigenesis in GCSCs through both miRNA-mediated and protein-mediated pathways (Fig. 7H).
留言 (0)