
記住我

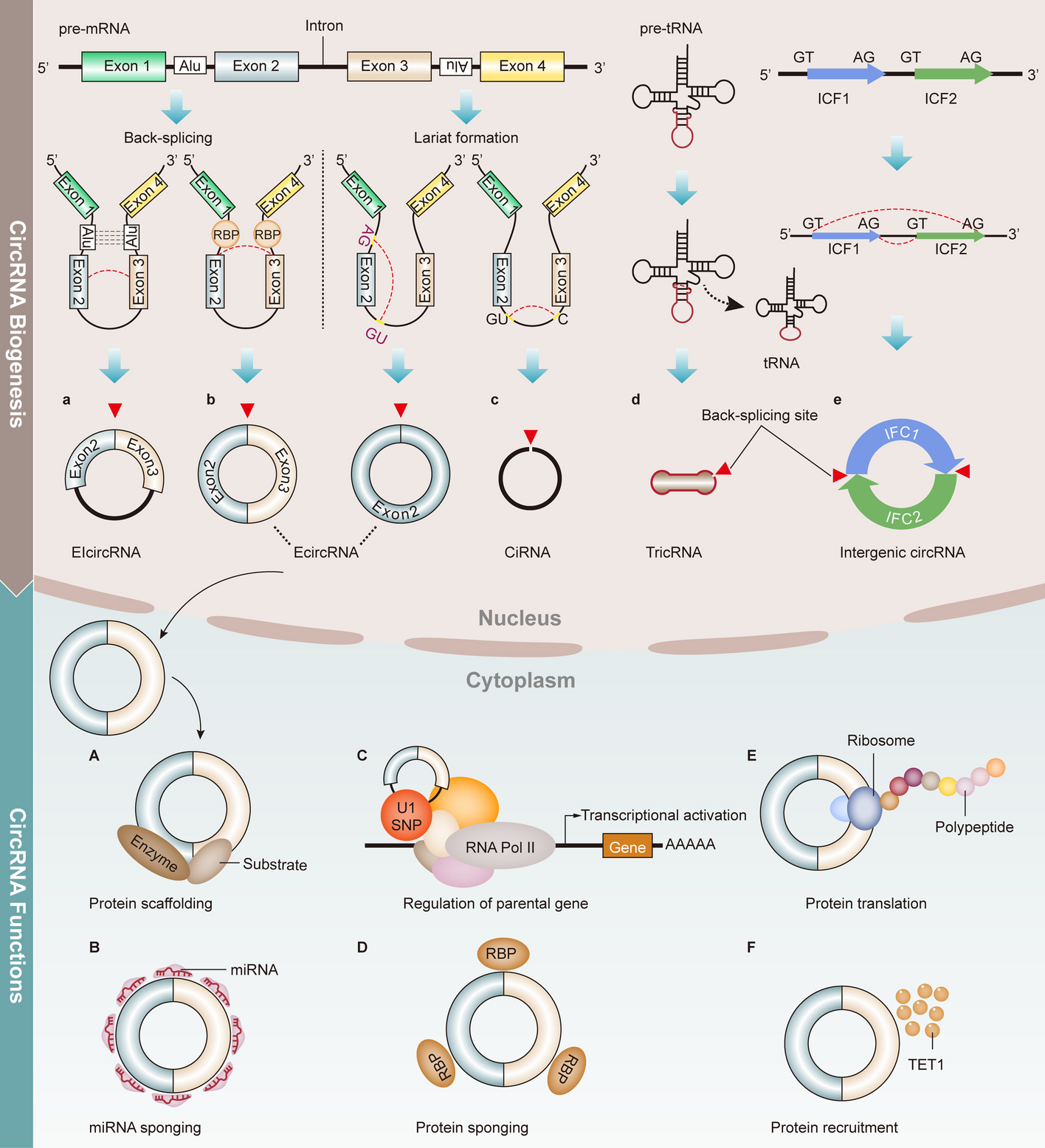

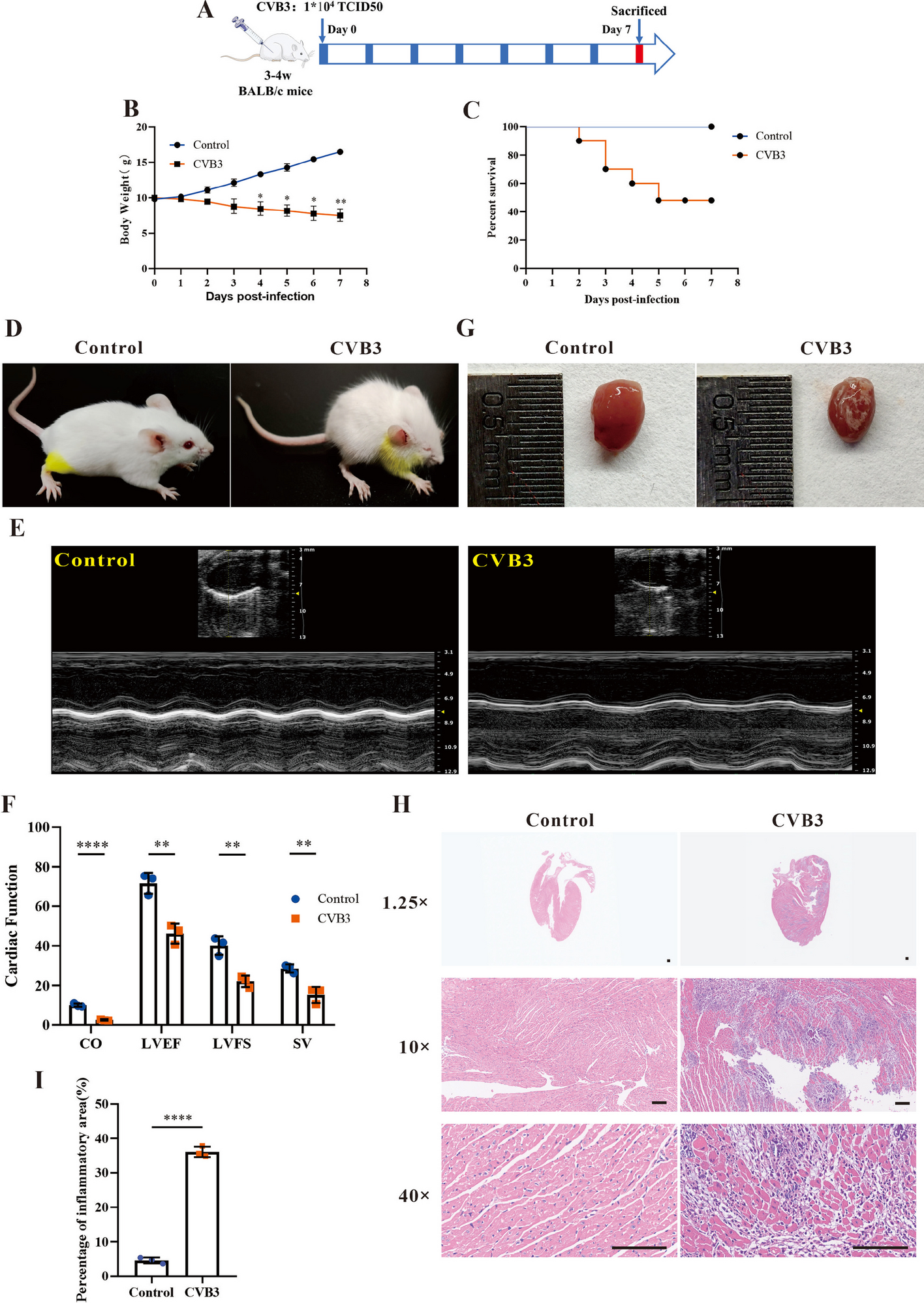
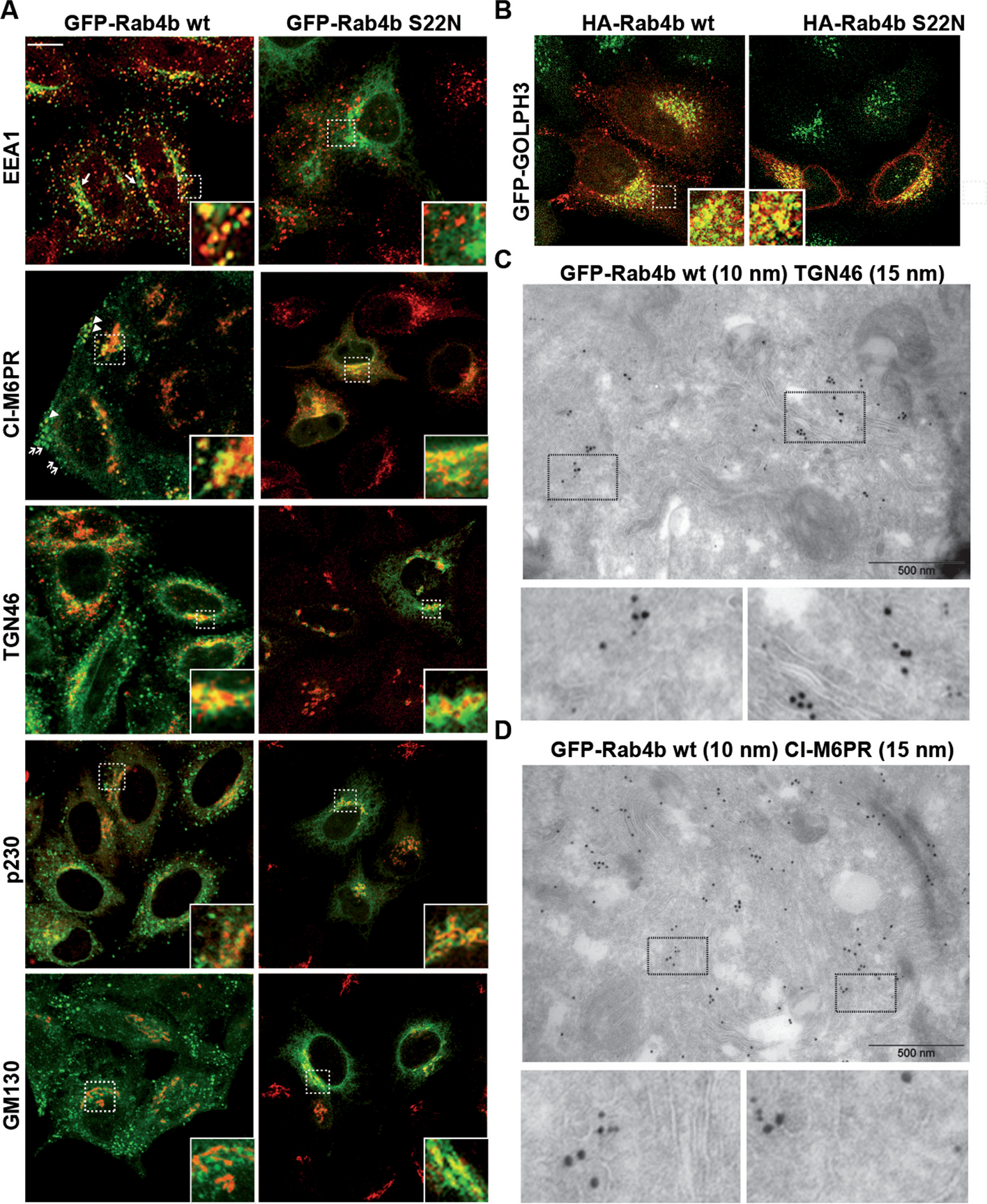
To study whether Rab4b was involved in early endosome to TGN retrograde trafficking, we first better characterized its subcellular localization. We characterized the distribution of Rab4b between intracellular organelles in HeLa cells stably expressing low amounts of Green-fluorescent protein-Rab4b (GFP-Rab4b), in order to minimize changes in organelle morphology that could be caused by a strong overexpression [27]. GFP-Rab4b was found colocalized with early endosomes (EEA1), the TGN (p230), and to a lower extent with the cis Golgi (GM130). Whereas nearly all the EEA1-positive endosomes were decorated by GFP-Rab4b, around 60% of GFP-Rab4b-positive structures were devoid of EEA1 (Additional file 1: Fig. S1B). The distribution of these GFP-Rab4b-positive and EEA1 negative compartments appeared bimodal with some structures at the cell periphery and some others in the perinuclear region. Because we previously characterized the peripheral vesicles as endosomes accessible to internalized transferrin (Tf) [27], we specifically studied the uncharacterized GFP-Rab4b-positive perinuclear tubular compartment (Fig. 1A, arrows in the panel showing EEA1 and GFP-Rab4b wt co-labeling). In these compartments, Rab4b was partly co-localized with CI-M6PR (cation-independent mannose-6-phosphate receptor) and TGN46 (also named trans Golgi network integral membrane protein 2, TGOLN2), two cargoes known to be recycled from the endo-lysosomes back to the trans Golgi network (TGN) (Fig. 1A). Consistently, ultrastructural exploration of GFP-Rab4b positive compartments by immuno-gold electron microscopy revealed that Rab4b was associated with tubulo-vesicular structures near the Golgi stacks and co-labeled with TGN46 (Fig. 1C) or CI-M6PR (Fig. 1D). In addition, we found that these compartments were partly co-labeled with the endogenous trans Golgi coiled-coil protein p230/golgin-245 (Fig. 1A) and over-expressed GOLPH3 (Fig.1B), but were only weakly co-labeled with the cis Golgi marker GM130/GOLGA2 (Fig. 1A and for quantification Additional file 1: Fig. S1A). To functionally discriminate between early endosomes and the TGN, which are very close in the perinuclear region of HeLa cells, we used two fluorescently labeled cargoes, the B subunits of cholera toxin (ChTx) and Tf, which, when internalized at steady state, labeled either the endosome to TGN retrograde pathway or the early and recycling endosomes, respectively (Fig. 2A). The perinuclear compartments labeled by GFP-Rab4b that accumulated the ChTx but not Tf (in yellow) probably corresponded to the TGN. Using the HeLa cell line stably expressing GFP-GOLPH3, which is located at the trans Golgi, and the chimeric retrograde cargoes consisting of the CI-M6PR fused to an extracellularly exposed CD8 antigen, we showed that internalized anti-CD8 antibodies trafficked back to perinuclear structures positive for both HA-Rab4b and GFP-GOLPH3 (white compartments in the image with the merge of all channels, Fig. 2B). Altogether, our results highlight that Rab4b is decorating the TGN in addition to early endosomes.
Fig. 1
Localization of Rab4b wt and S22N. A EEA1, CI-M6PR, TGN46, p230, or GM130 labeling (red) in HeLa cells expressing the same amount of GFP-Rab4b (wt) or GFP-Rab4b S22N (green) shown as merged images. were treated for immunofluorescence using antibodies and anti-species Texas Red-coupled secondary antibodies. The bar corresponds to 10 µm. Enlarged views of the demarcated area are also shown. Arrowheads indicate to GFP-Rab4b-positive vesicles containing M6PR, whereas arrows point to Rab4b-positive structures not containing M6PR. B merged images of GFP-GOLPH (green) and HA-tag (red) labeling. The bar corresponds to 10 µm. Enlarged views of the delineated area are shown. C, D TEM images of GFP-Rab4b (10 nm gold particles) and TGN46 C or CI-M6PR D (15 nm gold particles) using mouse anti-GFP and rabbit anti-TGN46 C or rabbit anti-GFP and mouse anti-CI-M6PR (D). Enlarged views of the delineated area are shown and bars are for 500 nm
Fig. 2
Cargoes accessibility. A Labeling of co-internalized Tf (iTf) and ChTx (iChTx), and of EEA1 in GFP-Rab4b expressing cells shown individually in white and black (left). The merged image of iTf (blue), iChTx (red), and GFP-Rab4b (green) (right). B Labeling of internalized CD8-M6PR (iCD8-M6PR, blue), in cells expressing GFP-GOLPH3 (green) and HA-Rab4b wt (red) shown individually in black and white, or as a merged image. Bars are for 10 µm
It was described that Rab GTPases cycle between an active GTP-bound form and an inactive GDP-bound form to coordinate membrane trafficking between two adjacent intracellular compartments. According to the current knowledge, inactive Rab GTPases are first recruited to the donor compartment and activated by the guanine nucleotide exchange factor (GEF). Afterwards, active Rab GTPases drive, through the recruitment of its effectors, the trafficking steps toward the target compartment, before its inactivation by the GTPase activating protein (GAP) [28, 29]. Therefore, we characterized where Rab4b-S22N, an inactive Rab4b protein accumulated. We found that Rab4b S22N was not localized in EEA1-positive early endosome (Fig. 1A and Additional file 1: Fig. S1A-B), but was detected in the TGN, where it colocalized with CI-M6PR, TGN46, p230, and GOLPH3 (Fig. 1A, B and Additional file 1: Fig. S1A). Moreover, a colocalization with the cis Golgi marker GM130 was also found (Fig. 1A and Additional file 1: Fig. S1A). These results suggest that GDP-bound Rab4b is associated with the Golgi, whereas GTP-bound Rab4b is rather present in early endosomes and peripheral vesicular structures. Hence, it is likely that Rab4b controls a trafficking step between endosomes and the TGN.
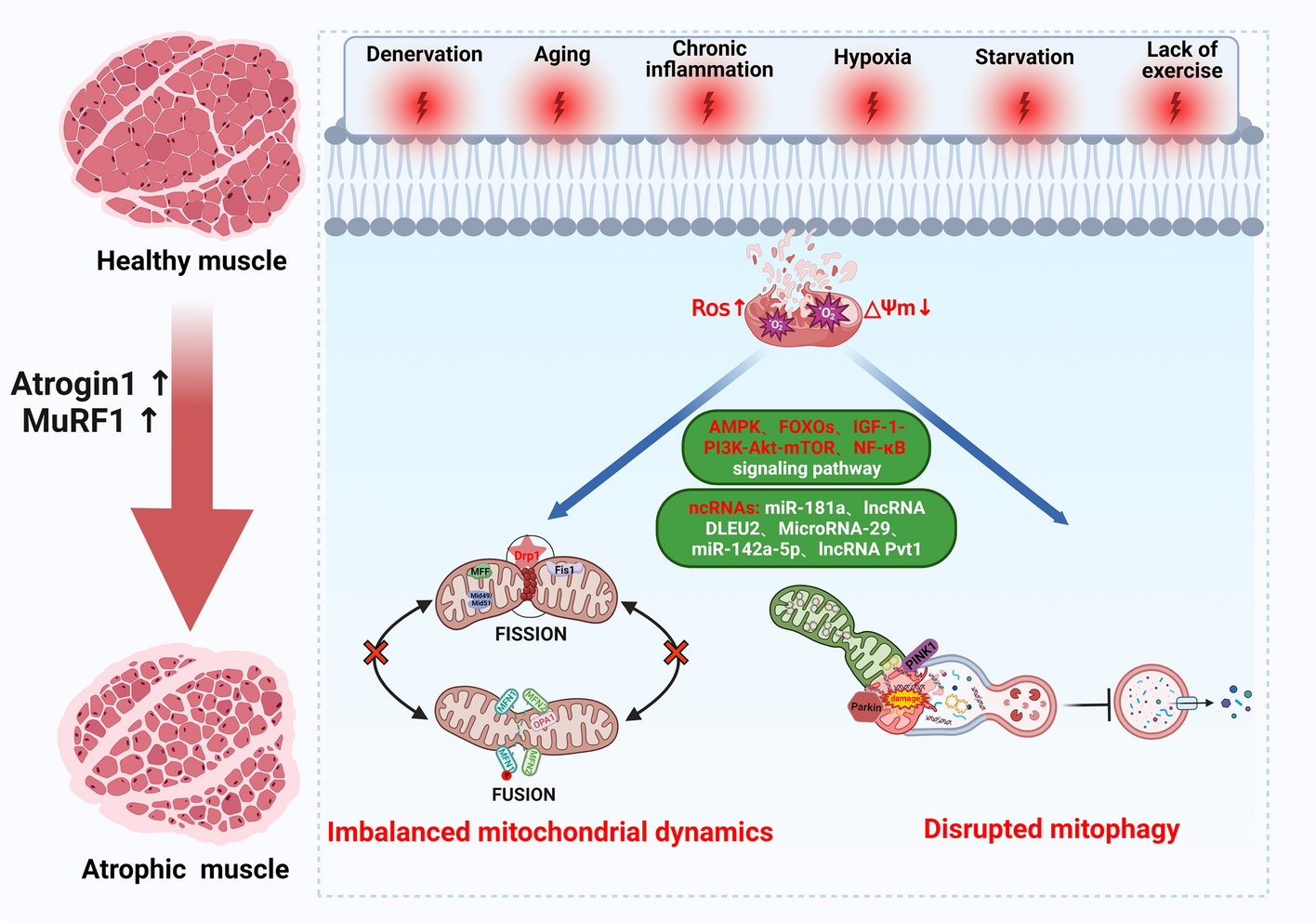
Retrograde transport to the trans-Golgi network is dependent on Rab4bWe next determined whether Rab4b was indeed involved in the retrograde transport to the TGN. First, we determined the consequences of the over-expression of Rab4b on the subcellular localization of Sortilin and/or CI-M6PR. These two proteins act as carriers for lysosomal enzymes, loading them at the level of the TGN. They then traffic from the TGN to the plasma membrane and/or the early endosomes, where they release the lysosomal enzymes into the endosome lumen, allowing them to reach late endosomes and lysosomes. The carriers are recycled from the endosomes back to the TGN by the retrograde trafficking pathway (for review [30]). Using density centrifugation on glycerol gradients, we observed that the overexpression of wt GFP-Rab4b increased the density of the intracellular compartments containing Sortilin (Fig. 3A).
Fig. 3
Active Rab4b favors retrograde trafficking to TGN. A Western blot of sortilin on homogenates separated on a glycerol gradient prepared from HeLa cells or HeLa cells expressing GFP-Rab4b wt. Representative blots and the quantification of 4 independent experiments. * indicate that the differences are significant with p < 0.05 (One Way ANOVA) B, C Quantification of the percentage of iCD8-M6PR in the Golgi area labelled with GFP-GOLPH3 in single cells expressing HA-Rab4b wt B or HA-Rab4b Q67L C compared to the surrounding and non-transfected cells. Representative images are shown (left) using the protocol 1 described in Additional file 1: Fig. S2A. Red stars indicate the HA-Rab4b expressing cells. Red and yellow arrowheads point to the periphery of the cells in cells expressing or not-expressing Rab4b, respectively. *** indicates that the differences are significant with p < 0.0001 (student t test). The bars are for 10 µm
We then investigated the effect of the overexpression of HA-Rab4b WT or a constitutively active form of Rab4b (HA-Rab4b Q67L) on the retrograde trafficking of CI-M6PR to the Golgi. For that, HeLa cells stably expressing CD8-CI-M6PR and GFP-GOLPH3 were transiently transfected to express HA-Rab4b WT or HA-Rab4b Q67L and then incubated with anti-CD8 antibodies at 37°C for 1h. In non-transfected cells (NT), the overexpressed CD8-CI-M6PR fusion protein that is internalized (hereafter referred to as internalized CD8-CI-M6PR, abbreviated iCD8-CI-M6PR) was found both in the Golgi and in the periphery of the cells (red arrowhead). iCD8-CI-M6PR localized in the periphery of the cells most likely labelled early endocytic compartments because they disappeared when a pulse-chase experiment was performed (see Fig. 4H and Additional file 1: Fig. S2C). In cells transfected with HA-Rab4b WT (Fig. 3B) or HA-Rab4b Q67L (Fig. 3C), the targeting of iCD8-CI-M6PR to the Golgi was more efficient. Only a faint signal of anti-CD8 antibodies was observed in HA-Rab4bQ67L-positive and GFP-GOLPH3 negative structures at the edge of the cells (yellow arrowhead). Thus, Rab4b overexpression favors CD8-CI-M6PR retrograde trafficking to the TGN and its GTPase activity is not required for this trafficking pathway.
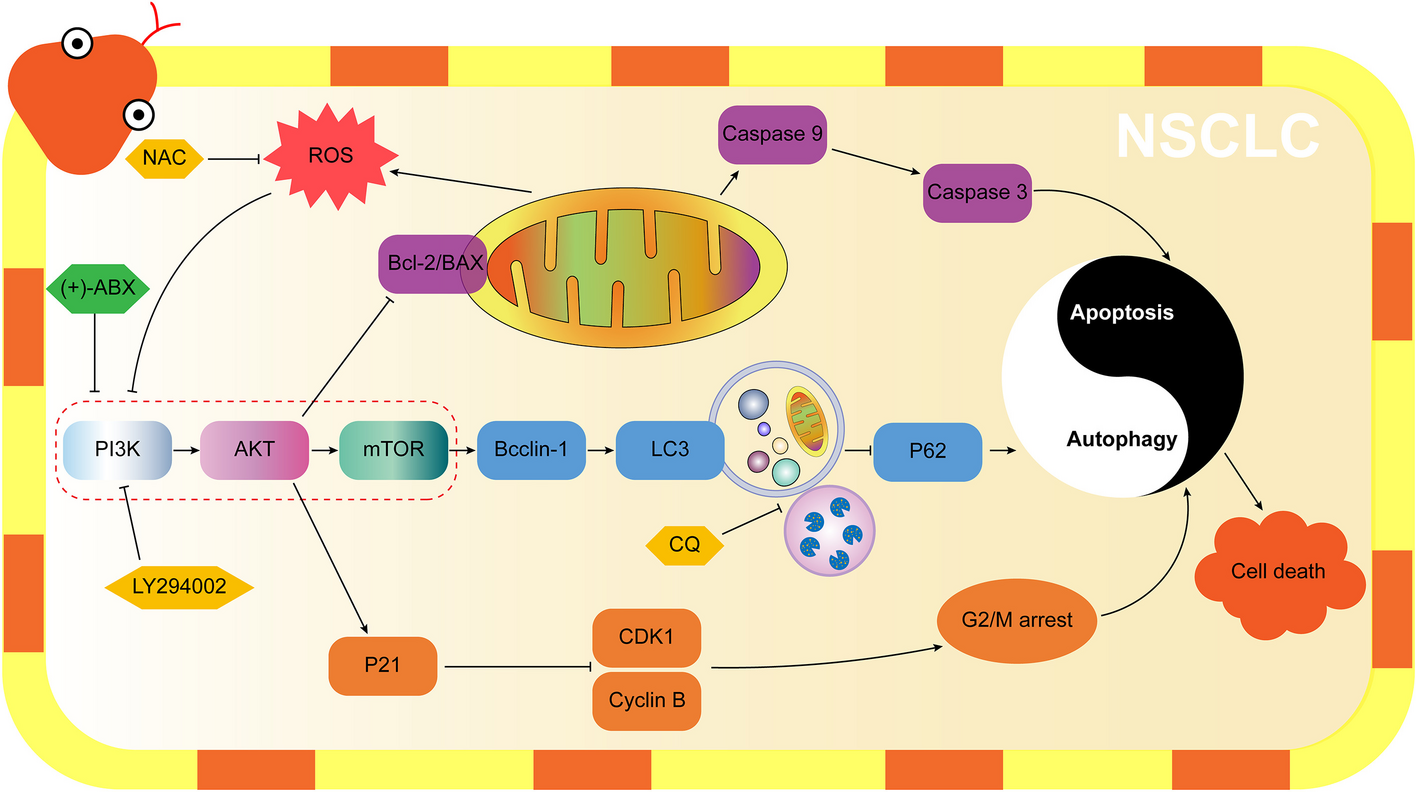
Fig. 4
Inactive Rab4b blocks retrograde trafficking to the TGN. A Western blots of Rab4b on lysates prepared from cells transfected with (ctrl: 30 nM) or anti-Rab4b siRNA (5, 30 nM) as described in [27]. Quantification of Rab4b mRNA expression in cells treated with 20 nM ctrl or anti-Rab4b siRNA. B Western blot of sortilin as in Fig. 3A from control- or Rab4b siRNA-treated cells, and quantification of 3 experiments. *, p < 0.05 (two-way ANOVA). (C) Western blots of CI-M6PR, sortilin, and tubulin as loading control from controlor Rab4b siRNA-treated cells incubated without or with leupeptin and E64 and the quantification of 3 independent experiments. Significance compared to the ctrl/no leu/E64 condition with * p < 0.05, ** p < 0.01, *** p < 0.001; significance compared with the Ctrl within the condition no leu/E64 and leu/E64 # p < 0.05 (one-way ANOVA). D Western blots of sortilin, and tubulin as loading control, from control or Rab4b siRNA treated wt cells or Rab4b overexpressing cells. E Quantification of the percentage of iCD8-M6PR at the Golgi in randomly acquired whole fields in control- or Rab4b siRNA-treated cells using the protocol 1 (Additional file 1: Fig. S2A). F I colocalization index (RColoc) between iCD8-CI-M6PR and Golgi, or iCD8-CI-M6PR and co-internalized Tf, determined in the same experiments as in E, using the colocalization plugin of the open ImageJ software [51]. G Quantification the percentage of iCD8-M6PR at the Golgi in randomly acquired whole fields in control- or Rab4b siRNA-treated cells using the protocol 2 (Additional file 1: Fig. S2A) and representative images on the left panel. H Quantification of iCD8-M6PR in the Golgi of single cells overexpressing HA-Rab4b S22N (red star marks) and in the surrounding non-transfected cells (NT) and representative images according to protocol 1 (Additional file 1: Fig. S2A). E–H Significance compared to siRNA ctrl condition with *p < 0.05, **p < 0.01, ****p < 0.0001 (Student t test). Bars are for 10 µm
We next determined whether Rab4b was necessary for endosomes to TGN retrograde trafficking. First, we knocked-down the expression of Rab4b by RNAi gene silencing as in [27] (Fig. 4A) and studied the consequences on Sortilin and/or CI-M6PR (Fig. 4C–G). Using density centrifugation on glycerol gradients, we observed that, in contrast to GFP-Rab4b WT overexpression (Fig. 3A), the Rab4b silencing reduced the density of the intracellular compartments containing Sortilin (Fig. 4B). Furthermore, the protein levels of CI-M6PR and Sortilin were also decreased (Fig. 4C, D). This decrease in expression was prevented by inhibition of lysosomal proteases with leupeptin and E64 (Fig. 4C), indicating that in the absence of Rab4b, Sortilin and CI-M6PR are misdirected to the lysosomes where they are degraded. Importantly, Sortilin expression was rescued in cells expressing a form of Rab4b insensitive to Rab4b siRNA (Fig. 4D). In addition, we knocked down Rab4b in HeLa cells stably expressing CD8-CI-M6PR and GFP-GOLPH3 and we determined the percentage of internalized anti-CD8 antibodies bound to CD8-CI-M6PR that reach the Golgi at steady state (Additional file 1: Fig. S2A condition 1). We found that this percentage is decreased by 2-fold while the index of colocalization with concomitantly internalized fluorescent Tf is increased (Fig. 4E–G and Additional file 1: Fig. S2B for representative images). Similar observations were made for the ChTx (Additional file 1: Fig. S2C). To determine whether Rab4b altered CD8-CI-M6PR sorting at the early endosomes, we performed a functional early endosome to TGN retrograde assay in the HeLa cells, depleted or not for Rab4b, and stably expressing CD8-CI-M6PR and GFP-GOLPH3. To do so, we loaded the early endosomes with internalized CD8-CI-M6PR by incubating the cells with anti-CD8 antibodies for 15 min at 20°C [31], before unleashing its trafficking by shifting to 37°C for the next 30 min (Additional file 1: Fig. S2A condition 2). We observed that the downregulation of Rab4b expression decreased the percentage of internalized CD8-CI-M6PR that reached the Golgi apparatus (Fig. 4G), as did the downregulation of VPS54 which is specifically involved in this retrograde trafficking pathway [32] (Additional file 1: Fig. S2D). Because Rab4b is required but its GTPase activity appears to be dispensable for its function in the retrograde trafficking pathway (see Fig. 3C), we determined whether GTP binding is needed. We expressed a form of Rab4b unable to bind GTP (HA-Rab4b S22N) in HeLa cells stably expressing CD8-CI-M6PR and GFP-GOLPH3 and incubated them for 1h at 37°C with anti-CD8 antibodies. In cells overexpressing HA-Rab4b S22N, the percentage of internalized CD8-CI-M6PR that reaches the Golgi area was reduced compared to the surrounding non-transfected cells and the non-Golgi internalized CD8-CI-M6PR remained dispersed throughout all the cytoplasm (Fig. 4H).
Overall, our results support that Rab4b is involved in the early endosomes to TGN retrograde trafficking pathway, and that its GTP binding activity, but not its GTPase activity, is required.
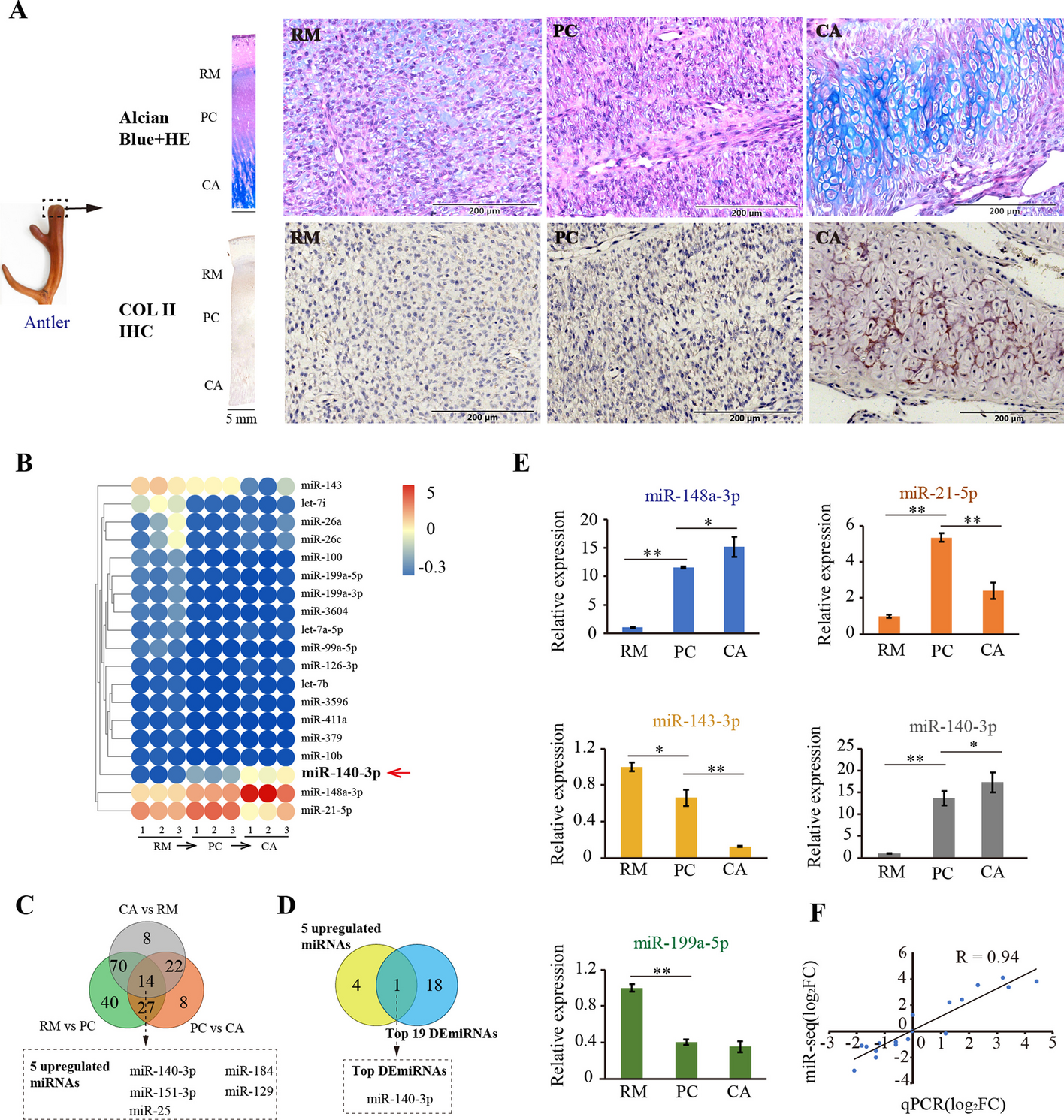
Active Rab4b favors endosomes to TGN retrograde trafficking in a GARP-dependent mannerWe first confirmed the interaction between VPS52 and Rab4b that was found in the high-throughput screen. We used the active form of Rab4b (Rab4bQ67L) as a bait and the full length VPS52 protein (aminoacid 1 to 723) as the prey in YTH. We found an interaction with Rab4bQ67L, but not with the inactive forms of Rab4b, Rab4bN121I, and Rab4bS22N (Fig. 5A, B), indicating that VPS52 can be an effector of Rab4b.
Fig. 5
Rab4b-dependent retrograde trafficking involves the GARP complex. A Schematic representation of VPS52 structures. The blue circles represent the coiled-coil domain. B Schematic representation of the interaction detected in the yeast two-hybrid system, performed as in [26, 27], between full-length VPS52 and the indicated forms of Rab4b or lamin as negative control. + + + + + is for an interaction is detectable before 30 min.—no detectable interaction after 24 h. (C) Quantification of iCD8-M6PR in the Golgi area of cells overexpressing HA-Rab4b Q67L (empty circles) and in the surrounding non-transfected cells (NT, filled circles) according to protocol 1 (Additional file 1: Fig. S2A). Each dot represents one cell. Each color (black, blue, or green) indicates independent experiments. ***,**** indicates that the differences are significant compared to NT siCtrl with and #### indicates that the difference is significant compared to NT siVPS54 with p < 0.001 or p < 0.0001 (one-way ANOVA). The significance is the same whether the 3 experiments are analyzed together or independently. D Representative images corresponding to the quantification shown in (C), stars are for transfected cells. The downregulation of VPS52 is validated in an independent well in each experiment by Western blot or immunolabeling as shown in (Additional file 1: Fig. S3A-B)
To explore whether the effect of Rab4b on the retrograde trafficking pathway of CD8-CI-M6PR depended on the GARP complex, we knocked-down VPS52, the effector of Rab4b, or VPS54, the specific subunit of the GARP complex, in cells overexpressing active Rab4b. While we confirmed that the overexpression of active Rab4b increased the targeting of CD8-CI-M6PR to the Golgi area, the efficient downregulation of VPS52 or VPS54 expression (Additional file 1: Fig. S3A) prevented this (Fig. 5C, D). The downregulation of the expression of these components of the GARP complex redistributed the internalized CD8-CI-M6PR to peripheral structures more efficiently in cells overexpressing HA-Rab4b Q67L, where internalized CD8-CI-M6PR and HA-Rab4b Q67L were colocalized, than in the non-transfected surrounding cells (Fig. 5D).
Our results support that the Rab4b-dependent early endosome to TGN retrograde trafficking route requires the GARP complex.
VPS52 is localized with Rab4b and p230 along the CI-M6PR retrograde trafficking pathwayTo strengthen the relationship between VPS52 and Rab4b in the retrograde trafficking route, we examined whether VPS52 is co-localized with GFP-Rab4b and how these two proteins were located relative to the marker of the TGN, p230/golgin-245. VPS52 was present in the Golgi area defined by the presence of p230, but was not particularly enriched there (Additional file 1: Fig. S3C, D). However, we observed some structures containing the three markers (white in the overlay) at the TGN, but also structures containing VPS52 and p230 without GFP-Rab4b (in purple). An orthogonal view of the Golgi area showed that GFP-Rab4b is over the TGN decorated by p230, and when GFP-Rab4b and p230 were in close contact, VPS52 was also present. In the periphery of the cells, there were enlarged vacuoles decorated by GFP-Rab4b and VPS52 without p230. Consistent with this, we found that the co-localization between GFP-Rab4b and VPS52 was greater than between p230 and VPS52 based on both intensity (Additional file 1: Fig. S3C) and object colocalization analyses in 3D reconstruction of confocal sections (Additional file 1: Fig. S3E). In addition, some vesicle-like structures between the Golgi area and the periphery of the cells were only positive for VPS52.
We then determined the distribution of CI-M6PR internalized at steady state between these different compartments. We applied super-resolution radial fluctuation (SRRF) analysis to increase the image resolution (Additional file 1: Fig. S3F). We confirm the extent of colocalization between VPS52, GFP-Rab4, and p230 as a function of the subcellular distribution (Fig. 6). In addition, it was shown that while VPS52 labeling is mostly punctate, GFP-Rab4 decorated tubulo-vesicular structures in contact with VPS52. Internalization of endogenous CI-M6PR (hereafter referred to as internalized CI-M6PR) filled GFP-Rab4b tubulo-vesicular structures in the Golgi area that were in contact with VPS52. This was either separated from the TGN (arrows) or in close contact with the TGN marker p230 (stars). These observations suggest that Rab4b and VPS52 work together to connect Rab4b-positive endosome-derived vesicles to the TGN for the retrograde trafficking of CI-M6PR.
Fig. 6
VPS52, Rab4b and p230 localize to the CI-M6PR retrograde pathway. A confocal section taken at the center of cells (left panels) treated with the super-resolution algorithm SRRF. Individual channels were shown with p230 in blue, GFP-Rab4b in green, VPS52 in red, and internalized CI-M6PR (iCI-M6PR) in light grey as well as the merged image of p230, GFP-Rab4b, and VSP52 or of all the channels. Enlarged views of the periphery and Golgi areas are shown, the regions are marked by squares in the whole field of the merge of all colors. Arrows point to vesicles labelled by GFP-Rab4b, VPS52 and iCI-M6PR without p230, whereas the stars point to structures where all the labeling are close together. Bar represents 10 µm
Rab4b is required for CI-M6PR to traffic from endosome to VPS52-positive vesicle-like structures in route to the TGNTo further elucidated the mechanism by which Rab4b through its effector VPS52 regulated the GARP-dependent retrograde trafficking of CI-M6PR, we knocked-down Rab4b and kinetically followed the retrograde trafficking steps from the endosomes towards the TGN, focusing not only on the cargoes but also on the VPS52-positive structures. First, we observed that the downregulation of Rab4b expression did not alter neither the number of VPS52-positive vesicles, nor the pattern of VPS52 localization within the cells (Fig. 7A, B). Second, using 3D object-based colocalization, as in Additional file 1: Fig. S3, we observed that the arrival of internalized CI-M6PR in VPS52-positive structures was identical in cells expressing or not Rab4b when internalized at 20°C for 15 min. Thirdly, the amount of VPS52-positive structures with internalized CI-M6PR was increased in control cells after 10 min chase at 37°C. However, the downregulation of Rab4b expression reduced the arrival of internalized CI-M6PR in VPS52-positive vesicles (Fig. 7A, B).
Fig. 7
CI-M6PR sorting to endosome carriers is impaired without Rab4b. Internalization of CI-M6PR (iCI-M6PR) is tracked by fluorescently coupled anti-CI-M6PR antibodies using protocol 2 (Additional file 1: Fig. S2A, 10 min chase instead of 30 min) in cells treated with control or anti-Rab4b siRNA. A Representative images after the 10 min chase (iCI-M6PR in 3D, after its volume reconstruction, VPS52 converted to spot, and VPS52 spot close to iCI-M6PR volume)The bar is for 20 µm. Image processing is described in the Material and Methods section. The same threshold and the same filters were applied for all the images. B Quantification of the number of VPS52 spots near the iCI-M6PR volume in cells treated with control siRNA (filled circles) and anti-Rab4b siRNA (empty circles) before and after the 10 min chase. **, *** indicate that the differences are significant with p < 0.01 and p < 0.001, respectively (one-way ANOVA)
This observation supports a role of Rab4b in the sorting of CI-M6PR from EE to VPS52-positive endosome-derived carriers. This step can therefore be considered as a prerequisite for the acquisition of directional specificity in the sorting of cargo carriers at the early endosomes (Fig. 8).
Fig. 8
Model for the role of Rab4b in the GARP-dependent retrograde trafficking pathway. The proposed possible model is based on results from this article and previous publications cited in the text. A In the absence of Rab4b, CI-M6PR and of the TfR are segregated and their trafficking route to the TGN and the recycling endosomes, respectively, is inhibited. The Rab4b effectors AP-1 and VPS52 remain on the endosomes. B In the absence of VPS52 and VPS54, CI-M6PR and of the TfR are not segregated, and they are targeted in a Rab4b/AP-1-dependent
留言 (0)