
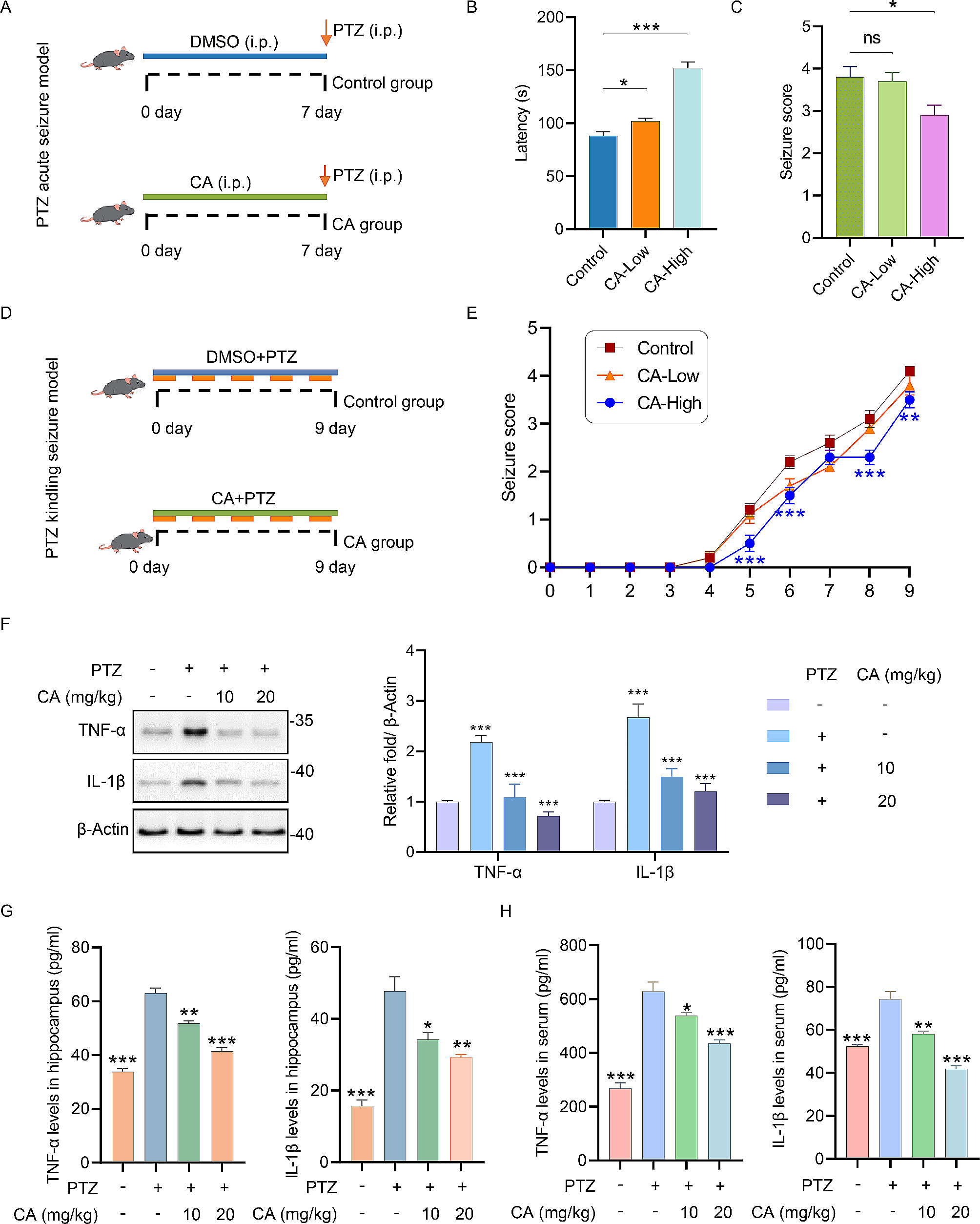
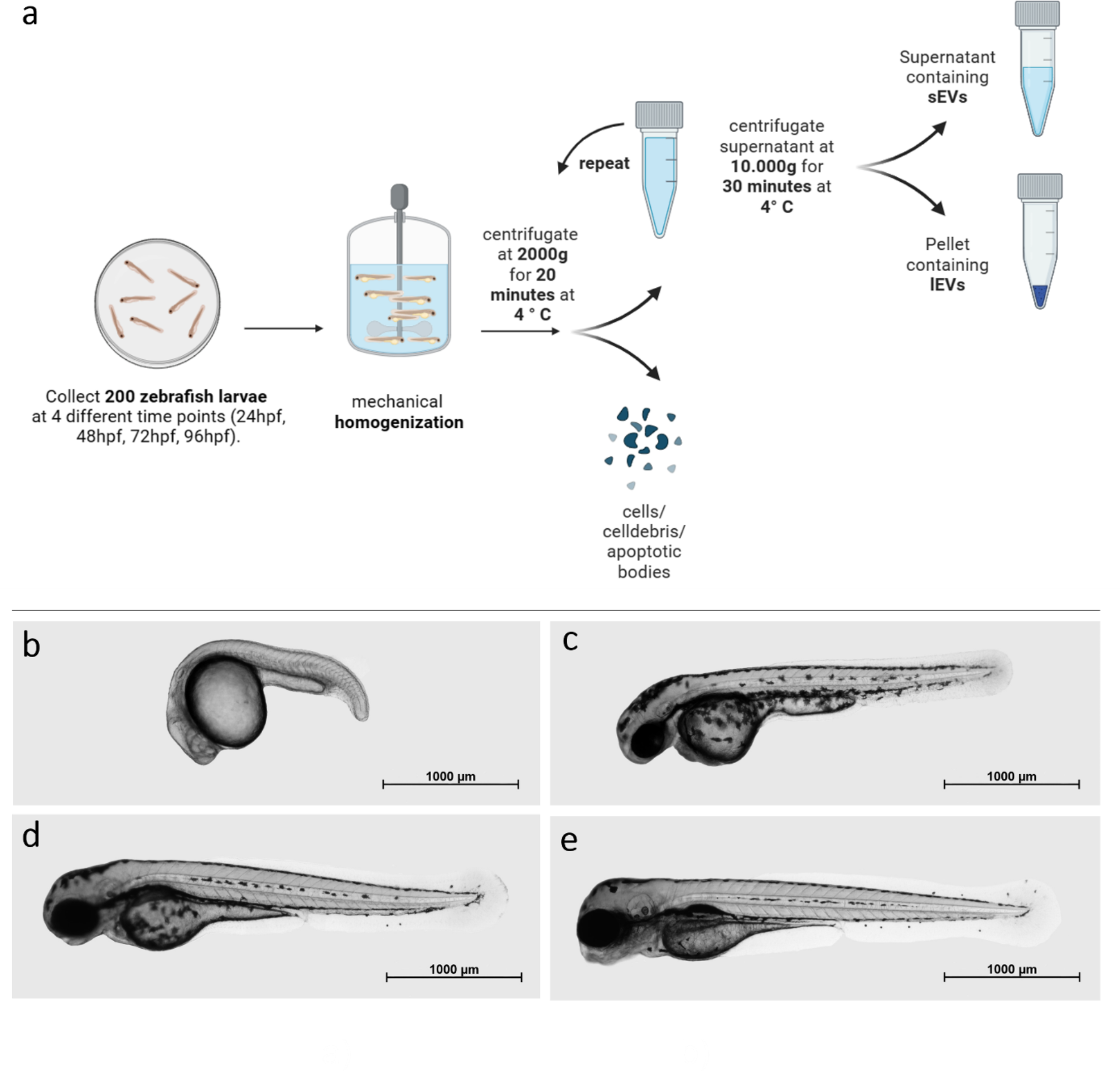
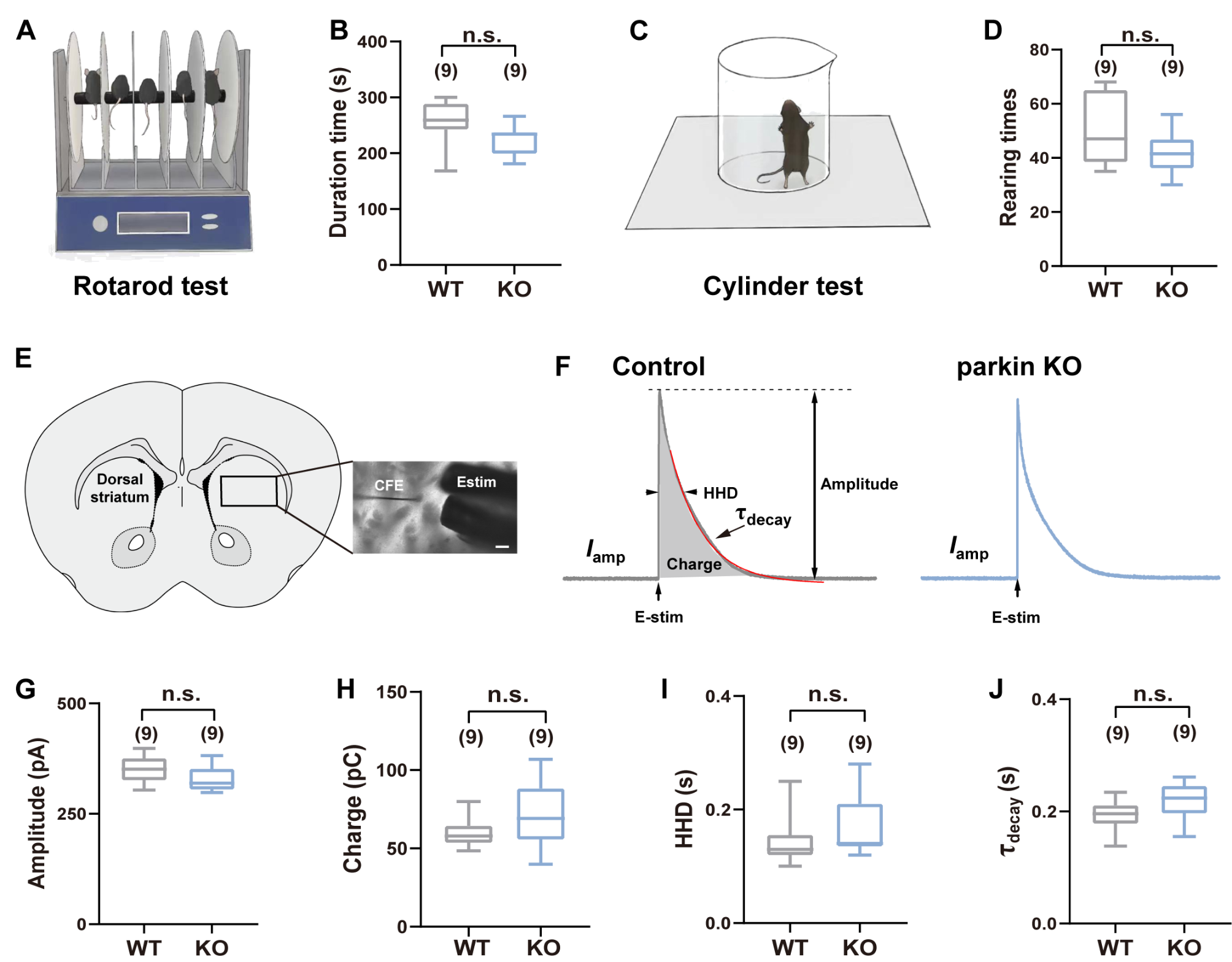
記住我







Kim et al. reported the presence of five ABCG-1 subfamily proteins in M. oryzae genome [18]. Here, we found that five of these proteins (MGG_13624/MoABC1, MGG_00447/MoABC2, MGG_07848, MGG_10277, MGG_07375) shared > 35% sequence similarity. Protein structure analysis showed that these identified proteins contain two typical ATP regions (NDBs) and two trans-membrane regions (TMDs) (Fig. 1A). Notably, MGG_07848 exhibited high sequence identity of 49.9%, 49.2%, 47.8%, 47.7%, and 48.3% with CaCdr1, CaCdr4, CaCdr2, and CaCdr3 of C. albicans and ScPdr5 of S. cerevisiae, especially in conserved regions (Fig. S1). Therefore, we designated MGG_07848 as MoCDR1 in this study.
Fig. 1
The sequence structure and cellular distribution of 5 ABCG-1 proteins in M. oryzae. (A) The phylogenic relationship and the sequence structures of the 5 ABCG-1 proteins. Blue boxes with AAA, ATP-binding domains; yellow boxes, transmembrane domains; pink boxes, low complexity regions; green box, coiled-coil. (B) The localization of MoCdr1and 4 ABCG-1 proteins in spores and hyphae under a laser confocal microscope. The GFP-labeled these proteins were expressed in wild type Guy11. Bar = 5 μm. (C) The localization of GFP-MoCdr1 in appressoria at 8 h post incubation (hpi). (D) The green fluorescence of GFP-MoCdr1 overlay well with a membrane-bound marker, FM4-64 straining (red fluorescence) in wild-type spores and hyphae. Bar = 5 μm
Further, we determined the subcellular localization of GFP-tagged ABCG-1 proteins, including MoCdr1, and found that all five proteins were membrane localized. The GFP signals were appeared on the cell membrane, vacuole membrane, or other inner membranes in mycelia, spores, and appressoria. This was further confirmed by co-localization with a membrane-bound marker FM4-64 (Fig. 1B-D). The membrane distribution of these proteins is consistent with the typical localization characteristics of ABC proteins.
MoCDR1 responds to metal ions and oxidative stress in M. oryzaeWe employed the gene replacement strategy to generate null mutants for each of the ABCG-1 genes, including MoCDR1 (Fig. S2). There were no significant differences in vegetative growth rates between deletion mutants and the wild type Guy11, when cultured on complete medium (CM) (Fig. S3A). However, drug sensitivity comparison revealed that these genes were involved in the fungus tolerance to various drugs. ∆Mocdr1 deletion mutants were more sensitive to fungicides myclobutanil (MBT) and benomyl (Table S1). We then assessed the growth of the wild type and ∆Mocdr1 deletion mutants (∆Mocdr1-8, ∆Mocdr1-13, and ∆Mocdr1-18) on CM containing metal stressors (calcium chloride, CaCl2; zinc chloride, ZnCl2; cuprous sulfate, Cu2SO4) and oxidative stressor (hydrogen peroxide, H2O2). The ∆Mocdr1 mutants grew more slowly on CaCl2, ZnCl2, Cu2SO4, and H2O2 media than the wild type (67.42%, 57.16%, 27.95%, and 62.0% reduction, respectively), indicating that loss of MoCdr1 caused significantly increased sensitivity to metal ions and oxidative stress. While reintroducing the intact MoCDR1 gene into the deletion mutant restored resistance to these stress factors (Fig. 2A and B). Intriguingly, the deletion of remaining four ABCG-1 genes did not affect the membrane distribution of GFP-MoCdr1 (Fig. S3C). Thus, these ABCG-1 proteins are likely independent in their subcellular distributions and functions but may complement each other in resistance against harmful chemicals.
Fig. 2
The ∆Mocdr1 mutants were sensitive to hydrogen peroxide and metal ions. (A) The wild type Guy11, ∆Mocdr1 mutants and complement strains were cultured on CM or the CM supplemented with ZnCl2, H2O2, Cu2SO4, CaCl2 for 9 days. (B) The relative growth rate was determined by measuring colony diameters on stress media against the control medium (CM). Asterisks indicate statistically significant differences between the wild type Guy11 and ∆Mocdr1 mutants (****, P < 0.0001)
MoCDR1 is involved in conidiation, appressorium development and pathogenicity of M. oryzaeDespite the vegetative growth rate of ∆Mocdr1 being comparable to the wild type Guy11 and complemented strain Mocdr1-c, the colony morphology of ∆Mocdr1 mutants displayed lighter colony pigmentation, drier and tighter mycelial surface in contrast to Guy11 (Fig. S4A and B). ∆Mocdr1 deletion mutants (∆Mocdr1-8, ∆Mocdr1-13 and ∆Mocdr1-18) showed similar genetic background and phenotypic defects, ∆Mocdr1-8 and corresponding complementation strain were selected for further analyses. Through analysis of spore production and conidiophores development in ∆Mocdr1-8 mutant, we found that the MoCDR1 deletion mutant produced significantly fewer conidia (about 1.5 × 106 conidia per plate) compared to the wild type (about 3 × 106 conidia per plate) (Fig. 3A and B). The conidial germination of ∆Mocdr1-8 was normal like the wild type (Fig. S4C). Then, we compared appressorium formation between the ∆Mocdr1-8 mutant and the wild type. The ∆Mocdr1-8 showed impaired appressorium formation, with the ratio of appressorium formation being significantly lower than that in the wild type after 4 - 10 h of incubation, and ∆Mocdr1-8 produced longer germ tubes (Fig. 3C and D). Additionally, during appressorium development of ∆Mocdr1-8, the translocation of lipid droplets and glycogen from conidia to appressoria was delayed at 8 h post inoculation (hpi), then the degradation of lipid droplets and glycogen in appressoria was hindered at 24 hpi (Fig. 3E-H). These results demonstrate that MoCDR1 is involved in regulating the translocation and utilization of storage compounds during appressorium maturation in M. oryzae.
Fig. 3
MoCDR1 is involved in conidiation and appressorium development. (A) Conidiophore development of Guy11, ∆mocdr1 and complement strain. Bar = 40 μm. (B) The conidiation of Guy11, ∆Mocdr1-8 mutant and the complement strain Mocdr1-8c. (C) The appressorial morphology of Guy11, ∆Mocdr1-8 and Mocdr1-8c on hydrophobic plastic coverslips. Bar = 20 μm. (D) The appressorium formation rate of Guy11, ∆Mocdr1-8 and Mocdr1-8c at 4, 6, 8, 10, and 24 hpi. (E and F) Observation of lipid droplets and glycogen in conidial and appressorial cells of Guy11, ∆Mocdr1-8 and Mocdr1-8c at 4 h, 8 h and 24 hpi. Lipid droplets were stained with the dye Bodipy, and glycogen was stained with KI/I2 solution. Bar = 5 μm. (G and H) The proportion of conidia containing lipid droplets and glycogen at 8 h, and appressoria containing lipid droplets and glycogen at 24 hpi. Asterisks indicate statistically significant differences between Guy11 and ∆Mocdr1-8 (*P < 0.05, **P < 0.01, ***P < 0.001, **** P < 0.0001)
To investigate whether ABCG-1 protein-encoding genes influence pathogenicity, we performed pathogenicity assays on rice and barley seedlings. Compared with the wild type Guy11, the pathogenicity was significantly decreased in the ΔMoabc1 mutant, while remained unchanged in the ΔMoabc2 mutant (Fig. S3B). Moreover, we showed that both MGG_07375 and MGG_10277 were dispensable for the pathogenicity, whereas MoCDR1 was required for the pathogenicity of M. oryzae. Notably, lesions caused by the ∆Mocdr1 mutants on both rice and barley leaves were significantly reduced in number and size compared to those caused by the wild type and complemented strains. Detached inoculation assays revealed that, ∆Mocdr1-inoculated rice and barley leaves showed markedly fewer lesions than the wild type and complemented strains-inoculated leaves (Fig. 4A and B). These results suggest that MoCDR1 is required for the full virulence of M. oryzae. We then performed appressorial infection assays with barley leaves. At 24 hpi, ∆Mocdr1 produced fewer penetration pegs compared to the wild type. At 48 hpi, the penetration rate of appressoria was near 90% in the wild type while less than 30% in the ∆Mocdr1-8 mutant. Furthermore, the colonization of ∆Mocdr1-8’s invasive hyphae inside the barley tissues were significantly delayed compared with the wild type (Fig. 4C and D). We also found that MoCDR1 deletion resulted in decreased phosphorylation levels of Pmk1 and Mps1 (Fig. 4E). These results suggest that MoCDR1 gene is involved in appressorium-mediated penetration and invasive growth by influencing the MAPK signaling pathway.
Fig. 4
The pathogenicity of ∆Mocdr1 mutants were decreased on rice and barley leaves. (A) The wild type Guy11, ∆Mocdr1 mutants and complement strains were spray-inoculated on rice and barley seedlings with 5 × 104 conidia mL− 1 suspensions, and droplet-inoculated on detached barley leaves with 1 × 105 or 1 × 104 conidia mL− 1 suspensions. (B) The lesions number of 5-cm rice and barley leaves caused by Guy11, ∆Mocdr1 mutants and complement strains. (C) The invasive hyphae of Guy11, ∆Mocdr1-8 and Mocdr1-c on barley leaves at 24 h and 48 h post incubation. The regions of invasive hyphae reached were surrounded by red dotted lines. Bar = 20 μm (D) The infection rates of appressoria of Guy11, ∆Mocdr1-8 and Mocdr1-c were counted at 24 h and 48 h post incubation. (E) The phosphorylation levels of Pmk1 (42 kDa) and Mps1 (44 kDa) in Guy11 and ∆Mocdr1-8 mutant. Actin was used as an internal reference protein (42 kDa). Asterisks indicate statistically significant differences between Guy11 and ∆Mocdr1 (**P < 0.01, **** P < 0.0001)
Identification and structural analysis of TmCDR1 in T. mentagrophytesTo identify the homologue of MoCDR1 in M. oryzae from T. mentagrophytes, we identified a total of 45 putative ABC family genes in T. mentagrophytes from the NCBI database. CDD analysis and protein structural analysis revealed that all 45 proteins possess a typical ABC structure. Phylogenetic analysis of the protein sequences further divided these 45 proteins into seven subfamilies, among which the ABCG-1 subfamily comprises six proteins (Fig. 5A). Remarkably, a gene, GBF63800, displayed 48.9%, 50.9% and 55.2% sequence homology with MoCDR1, CaCDR1 and ScPDR5, respectively. Here, GBF63800-encoded protein was designated as TmCDR1. Additionally, the two ATP binding domains (NBDs) and two transmembrane domains (TMDs) between TmCdr1 and its homologues MoCdr1, CaCdr1, ScPdr5, TruMdr3, FgAbc3 of Fusarium graminearum showed high sequence identity (Fig. 5B and S5). We further displayed the predictive 3D (three-dimensional) structures of TmCdr1, MoCdr1 and their homologues from AlphaFold2 and AlphaFold3 database models. The six proteins have greatly similar 3D structures, particularly the α helices of transmembrane domains (Fig. 5C). These data implys that TmCdr1, MoCdr1 and their homologues remain largely conserved in both sequence and transporter conformation.
Fig. 5
Phylogenetic analysis of ABC transporters in T. mentagrophytes and alignment of TmCDR1 homologues. (A) Phylogenetic analysis of the 45 putative ABC proteins in T. mentagrophytes. The predicted protein sequences were aligned with ClustalW, and the MEGA software version 5.0, using the neighbor-joining method. The terminology of ABC subfamily, ABCB, ABCC-1, ABCD, ABCE, ABCF, ABCG-1 and ABCG-2 were adopted from the previous study [2]. Arrow indicates TmCdr1-encoding gene. (B) Alignment of protein sequences of TmCdr1 homologues. The protein sequences of TmCdr1 and the homologues derived from M. oryzae (MoCdr1), C. albicans (CaCdr1), S. cerevisiae (ScPdr5), T. rubrum (TruMdr3) and F. graminearum (FgAbc3) were aligned with DANMAN software version 8.0. The partial amino acid sequence of ATP-binding domains (NBD1, NBD2) and transmembrane domains (TMD1, TMD2) were selected to show. (C) Cartoon models of TmCdr1 homologues. The structure was constructed using Pymol software based on AlphaFold2 and AlphaFold3
TmCDR1 is a functional homologue of MoCDR1 in drug resistance and pathogenicityNext, we determine the subcellular distribution of TmCdr1. GFP-TmCdr1 and mCherry-TmCdr1 were introduced into the wild type strain of T. mentagrophytes. In hyphae and spores, both GFP-TmCdr1 and mCherry-TmCdr1 displayed a typical membrane localization (Fig. 6A).
Fig. 6
TmCDR1 is a membrane protein and capable of complementing the defects of the ∆Mocdr1 mutant. (A) GFP-TmCdr1 or mCherry-TmCdr1 is localized in hyphae and spores of wild type ZJA-1. (B) The growth of the wild type Guy11, ∆Mocdr1-8 mutant, ∆Mocdr1-8::GFP-TmCDR1, ∆Mocdr1-8::mCherry-TmCDR1 and ∆ Mocdr1-8::GFP-MoCDR1 strains cultured on CM or the CM supplemented with ZnCl2, CaCl2, H2O2 for 9 days. (C) The relative growth rate was determined by measuring colony diameters on stress media against the control medium (CM). (D) Guy11, ∆Mocdr1-8 mutant, ∆Mocdr1-8::GFP-TmCDR1, ∆Mocdr1-8::mCherry-TmCDR1 and ∆Mocdr1-8::GFP-MoCDR1 strains were droplet-inoculated on detached barley leaves with 1 × 105 or 1 × 104 spore mL− 1 suspensions. Asterisks indicate statistically significant differences between the Guy11 and ∆Mocdr1-8 mutant (*** P < 0.001, **** P < 0.0001)
Given the sequence homology and similar membrane distribution between TmCdr1 and MoCdr1, we sought to investigate whether these two proteins are functionally related. For this purpose, we introduced the fusion proteins GFP-TmCdr1 and mCherry-TmCdr1 into the ∆Mocdr1-8 mutant to obtain the ∆Mocdr1::mCherry-TmCDR1 and ∆Mocdr1-8::GFP-TmCDR1 strains. Surprisingly, TmCdr1 restored the resistance of ∆Mocdr1-8 mutant to CaCl2, ZnCl2 and H2O2 (Fig. 6B and C). Furthermore, TmCDR1 effectively reinstated the pathogenicity of the ∆Mocdr1-8 mutant. The disease symptoms caused by ∆Mocdr1-8::mCherry-TmCDR1 and ∆Mocdr1-8::GFP-TmCDR1 strains were comparable to those of the wild type Guy11 and ∆Mocdr1-8::GFP-MoCDR1 strains (Fig. 6D). Thus, these results confirm that TmCDR1 is a functional homologue of MoCDR1.
TmCDR1 mediates multidrug resistance of T. mentagrophytesTo characterize the function of the TmCDR1 gene in T. mentagrophytes, TmCDR1 null mutant was generated using a gene replacement strategy (Fig. S6). The growth, colony morphology, and conidial germination of the ∆Tmcdr1 mutants were similar to those of the wild type ZJA-1 and complemented strains ΔTmcdr1::GFP-TmCDR1 (Tmcdr1-c) (Fig. 7A-C). Similar to ∆Mocdr1 mutants, the ∆Tmcdr1 mutants exhibited significantly higher sensitivity to CaCl2 than the wild type. Additionally, the ∆Tmcdr1 mutants were more sensitive to the fungicides, myclobutanil (MBT) and berberine (BBR), the major effective ingredients of the traditional Chinese medicine Coptis chinensis (Fig. 7C and D).
Fig. 7
The phenotypic analysis of ∆Tmcdr1 mutants. (A) The conidia development of the wild type ZJA-1, ∆Tmcdr1 mutants and the complement strains at 12 h, 14 h and 16 h post incubation. (B) The spore germination rates of ZJA-1, ∆Tmcdr1 mutants and the complement strains. No significant difference between these strains. (C) ZJA-1, ∆Tmcdr1 mutants and the complement strains were cultured on SDA and SDA containing berberine (BBR), CaCl2 and Myclobutanil (MBT) for 7 days. (D) The growth inhibition rate of berberine, CaCl2 and Myclobutanil on these strains. (E) Relative transcript levels of TmCDR1 in the wild type cultured on SDA containing berberine and clotrimazole (CMZ) compared to SDA. (F) The virulence of these strains was tested by inoculation on rabbit’s skin for 10 days post inoculation. (G) The different expression genes (DEGs) in Raw264.7 mouse macrophages inoculated with ZJA-1 and ∆Tmcdr1-1 mutant compared with untreated mouse macrophages (CK). The red points shows the up-expression genes, green points shows the down-expression genes, and blue points shows the unchanged genes. Asterisks indicate statistically significant differences between ZJA-1 and ∆Tmcdr1 mutants (*P < 0.05, **P < 0.01, ***P < 0.001, **** P < 0.0001)
Furthermore, we analyzed the expression of 26 out of the 45 TmABC genes in the wild type strain of T. mentagrophytes treated with distinct drugs. Treatment with BBR at a concentration of 500 µg/mL led to an upregulated expression of 9 TmABC genes while decreasing the expression of 6 TmABC genes. Similarly, clotrimazole (CMZ) treatment significantly decreased the expression of 14 TmABC genes while markedly increasing the expression of 3 TmABC genes (GBF60718, GBF64815, and TmCDR1) (Fig. S7). Notably, TmCDR1 was upregulated following treatment with both BBR or clotrimazole (Fig. 7E). These results suggest an association between the ABC family genes and drug resistance in T. mentagrophytes, with TmCDR1 mediating resistance to both BBR and clotrimazole.
TmCDR1 is required for the animal infection of T. mentagrophytesTo determine whether TmCDR1 participates in the infection process of T. mentagrophytes on animal skin, the conidia of the wild type ZJA-1 and ∆Tmcdr1 were inoculated on rabbit skin. Following 10 d post-inoculation, the rabbit skin sites inoculated with the wild type ZJA-1 exhibited typical symptoms of dermatophytosis, including grayish-white skin rashes, and reddish skin. ∆Tmcdr1 strains produced slightly less lesions compared to the wild type, exhibiting typical skin damages (Fig. 7F). Considering that the skin lesions on the rabbits are difficult to quantify and make accurate comparisons, we further conducted comparative transcriptomic analysis of healthy and ∆Tmcdr1-1 or ZJA-1 infected mouse macrophages. Compared to healthy mouse macrophages, a total of 62 disease-resistant genes exhibited altered expression in macrophages following infection with ZJA-1, with 42 up-regulated and 20 were down-regulated (p-value < 0.05, log2fold change > or < 2). In ∆Tmcdr1-1-infected macrophages, 39 DEGs were detected, with 21 were up-regulated and 18 were down-regulated (p-value < 0.05, log2fold change > or < 2) (Fig. 7G). Among the upregulated genes in ZJA-1-infected mouse macrophages, the expression levels of FOXP3, ASB4, and WNT2B were significantly elevated (Fig. S8). FOXP3 is a transcriptional factor crucial for the development of conventional T-cells, regulating inflammation and immune response in mammalian cells [34, 35]. ASB4 (Ankyrin repeat and suppressor of cytokine signaling box protein 4) interacts with NF-κB (nuclear factor-κB) in endothelial cells and functions in the nervous system by triggering the expression of the proopiomelanocortin gene in mice [36, 37]. WNT2B activates the NF-κB pathway and enhances the expression of downstream inflammatory cytokines in mouse macrophages [38, 39]. These important disease-resistant genes were not detected in mouse macrophages infected with ∆Tmcdr1-1 strain. Collectively, these results indicate that TmCDR1 is required for the full virulence of T. mentagrophytes on animals.
MoCDR1 partially rescues the defects of drug resistance and virulence of the TmCDR1 deletion mutantSince TmCDR1 functions in drug resistance and full virulence of T. mentagrophytes, and restores the functional defects of ΔMocdr1-8 mutant, we infer whether MoCDR1 can also restore the defects of ΔTmcdr1-1 mutant. The GFP-MoCdr1 fusion vector was expressed in the ΔTmcdr1-1 mutant to obtain the complemented strain ΔTmcdr1-1::GFP-MoCDR1. Compared to the wild type ZJA-1 and ΔTmcdr1-1::GFP-TmCDR1 (Tmcdr1-1c), MoCdr1 effectively restored the ΔTmcdr1-1 mutant’s resistance to CaCl2, but only partially restored its resistance to berberine (Fig. 8A). Next, we conducted mouse skin infection experiments to test whether MoCdr1 could also restore the pathogenicity defect of ΔTmcdr1-1. Compared to the severe symptoms of dermatophytosis caused by ZJA-1 and Tmcdr1-1c on the mouse skin, the ΔTmcdr1-1 mutant exhibited reduced pathogenicity. The lesions caused by ΔTmcdr1-1::GFP-MoCDR1 were more severe than those caused by ΔTmcdr1-1 but weaker than those caused by the wild-type and Tmcdr1-1c. Furthermore, HE staining showed that the skin tissues infected with ZJA-1 and Tmcdr1-1c exhibited significant pathological features, including massive inflammatory cell infiltration, prominent granulation tissue, and subcutaneous fibrous tissue proliferation. The pathological feature caused by ΔTmcdr1-1::GFP-MoCDR1 was relatively mild than ZJA-1 and Tmcdr1-c, while it was more pronounced than that caused by the ΔTmcdr1-1 mutant (Fig. 8B). These results indicate that MoCdr1 can partially restore the defects of drug resistance and pathogenicity of the ΔTmcdr1-1 mutant.
Fig. 8
MoCDR1 is capable of complementing the defects of the ∆Tmcdr1-1 mutant. (A) The growth of the wild type ZJA-1, ∆Tmcdr1-1 mutant, ∆Tmcdr1-1::GFP-TmCDR1 and ∆Tmcdr1-1::GFP-MoCDR1 strains cultured on SDA or the SDA supplemented with CaCl2, BBR for 9 days. (B) The growth inhibition rate of CaCl2, BBR on these strains. (C) The virulence of ZJA-1, ∆Tmcdr1-1 mutant, ∆Tmcdr1-1::GFP-TmCDR1 and ∆Tmcdr1-1::GFP-MoCDR1 strains was tested by inoculation on mice’s skin for 10 days post inoculation. (D)The skin tissues from the lesioned areas was stained with for hematoxylin and eosin (HE) staining analysis to detect the pathological features. Bar = 50 μm. Asterisks indicate statistically significant differences between the ZJA-1 and ∆Tmcdr1-1 mutant (**** P < 0.0001)
Transcriptome changes in M. oryzae and T. mentagrophytes induced by MoCDR1 and TmCDR1 deletionsTo gain a more accurately understanding of the functional correlation between MoCDR1 and TmCDR1, we performed transcriptome analysis. Compared with the wild type Guy11 strain, the ∆Mocdr1-8 strain exhibited 1765 up-regulated DEGs and 1204 down-regulated DEGs (Fig. 9A). In the ∆Tmcdr1-1 strain, a total of 535 DEGs were detected compared to the wild type ZJA-1 strain, with 324 up-regulated and 211 down-regulated DEGs (Fig. 9B). KEGG analysis revealed that the DEGs of ∆Mocdr1-8 were mainly enriched in 11 pathways, including ribosom biogenesis, starch and sucrose metabolism, fructose and mannose metabolism, and the MAPK signaling pathway, etc. The DEGs of ∆Tmcdr1-1 were involved in 16 pathways, such as the biosynthesis of secondary metabolites, amino acids, glycolysis/gluconeogenesis, peroxisome, fructose and mannose metabolism, and MAPK signaling pathway, etc. Notably, the DEGs of both ∆Mocdr1-8 or ∆Tmcdr1-1 were significantly enriched in the same 5 KEGG pathways: starch and sucrose metabolism, fructose and mannose metabolism, cyanoamino acid metabolism, MAPK signaling pathway, biosynthesis of secondary metabolites, and tyrosine metabolism (Fig. 9C and D).
Fig. 9
Transcriptomic analysis of ∆Mocdr1-8 and ∆Tmcdr1-1 mutants. (A) The DEGs in ∆Mocdr-8 compared with back ground strain Guy11. (B) The DEGs in ∆Tmcdr1-1 compared with back ground strain ZJA-1. (C) Venn diagram analysis of KEGG pathways between two groups: ∆Mocdr1-8 and Guy11, ∆Tmcdr1-1 and ZJA-1. (D) KEGG enrichment of DEGs in ∆Mocdr1-8 mutant compared with Guy11, and KEGG enrichment of DEGs in ∆Tmcdr1 mutant compared with ZJA-1. Red asterisks indicate the same pathways between these two groups. P value of the KEGG enrichment terms are less than 0.05. (E) Heatmaps of selected DEGs in MAPK signaling pathway and other metabolic processes. Red indicates relatively high expression and blue indicates relatively low expression. The DEGs underwent unsupervised clustering using complete linkage and euclidean distance
Importantly, MAPK signaling pathway is involved in the pathogenic processes of M. oryzae and T. mentagrophytes. In the ∆Mocdr1-8 strain, the core genes of MAPK pathway, including the MAP kinase kinase kinase Mck1 (MGG_00883) [40], high osmoregulation signaling protein Sho1 (MGG_09125) [41], protein kinase regulator Mst50 (MGG_05199) [42], were significantly downregulated. In the ∆Tmcdr1-1 strain, we found that MAPK pathway-related genes, such as genes encoding glycerol-3-phosphate dehydrogenase (TMEN_4625) and GTP-binding protein (TMEN_5507), were significantly upregulated. Additionally, many DEGs related to ABC membrane proteins and other transporters, such as genes encoding drug multidrug resistance proteins (MGG_12612, MGG_07375, MGG_05044/ABC6, TMEN_8641), amino acids or glucose transporters (MGG_03341, MGG_10508, MGG_06203, TMEN_3047, TMEN_6844) and metal tolerance proteins (MGG_01862, MGG_08710, MGG_05190, TMEN_2052, TMEN_6869, TMEN_9261) showed significantly alterations in the ∆Mocdr1-8 and ∆Tmcdr1-1 mutants (Fig. 9E). These results indicate that the deletion of MoCDR1 and TmCDR1 affects membrane transport of drugs, as well as pathogenic and metabolic processes, thus leading to defects in multi-drug resistance and virulence in M.oryzae and T. mentagrophytes. These similar processes were likely to explain why TmCDR1 could rescue the defects of the deletion mutant ∆Mocdr1, and vice versa.
To further validate that the MoCDR1 or TmCDR1 deletions affect membrane transport activity, we have detected the transcriptional modulation in response to different drugs by qRT-PCR. Compared with the wild type, treatment with ZnCl2, the MoCDR1 deletion led to changes in the expression levels of MGG_03341, MGG_06203, MGG_05190, and the TmCDR1 deletion resulted in changes in the expression levels of TMEN_3047, TMEN_2052, TMEN_6869, TMEN_9261. Treatment with CaCl2, the MoCDR1 deletion led to changes in the expression levels of MGG_05044, MGG_03341, MGG_08710, MGG_05190, and the TmCDR1 deletion resulted in changes in the expression levels of TMEN_3047, TMEN_6844, TMEN_9261. Treatment with BBR, the MoCDR1 deletion led to changes in the expression levels of MGG_12612, MGG_07375, MGG_06203, MGG_01862, MGG_08710, MGG_05190, and the TmCDR1 deletion resulted in changes in the expression levels of TMEN_8641, TMEN_3047, TMEN_2052, TMEN_6869, TMEN_9261. Treatment with CMZ, the MoCDR1 deletion led to changes in the expression levels of MGG_12612, MGG_03341, MGG_10508, MGG_08710, and the TmCDR1 deletion resulted in changes in the expression levels of TMEN_8641, TMEN_6844, TMEN_2052, TMEN_6869, TMEN_9261 (Fig S9). These results assume that MoCDR1 or TmCDR1 is directly involved in drug transport, or may regulate membrane transport processes by influencing the activity of other transporters, so that the deletions of MoCDR1 or TmCDR1 affect the membrane transport of drugs.
留言 (0)