
記住我



The aged immune system contributes to systemic aging, serving as a driver of morbidity and mortality in the elderly. In a recent Cell Research paper, Wang et al. identified both resilient and dysfunctional subsets of old hematopoietic stem cells (HSCs), of which selective reduction of dysfunctional old HSCs ameliorates aging phenotypes and extends lifespan in mice.
Identifying therapeutic targets of systemic aging could begin to address the growing burden of age-related diseases and morbidity in old age. Blood, and the components therein, are posited drivers of aging1,2 and therapeutic targets for rejuvenation.3 Previous work in aging mouse models demonstrated that heterochronic transplantation of bone marrow or peripheral immune cells restore healthspan and extend lifespan3,4 — pointing to the potential of targeting hematopoietic stem cells (HSCs) themselves. HSCs are characterized by self-renewal capacity and the ability to give rise to all cells of the blood. Although the HSC reservoir expands with age, HSCs undergo major age-related changes that decrease regenerative potential, bias lineage potential in favor of myeloid cells, and reduce niche-homing efficiency. The age-related shift in differentiation towards myeloid cells at the expense of lymphoid cells results in a maladaptive immune system, and targeting myeloid-biased HSCs can rejuvenate the aged immune system.5 However, whether targeting subsets of HSCs impacts systemic aging has not been assessed.
Given recent studies showing the rejuvenating potential of a young hematopoietic system and increased HSC heterogeneity during aging, Wang and colleagues sought to identify subpopulations of HSCs with more resilient molecular profiles that could mitigate aging phenotypes (Fig. 1). The authors established functional differences between young and old HSCs in mice, confirming with competitive transplant assays that young HSCs exhibit superior engraftment and reconstitution of the immune system. This observed age-related difference is maintained in secondary transplants, further supporting proposed cell-intrinsic aging mechanisms. Functionally, recipient mice with young donor HSCs exhibited attenuated aging phenotypes, as previously observed with heterochronic bone marrow transplants.4,6
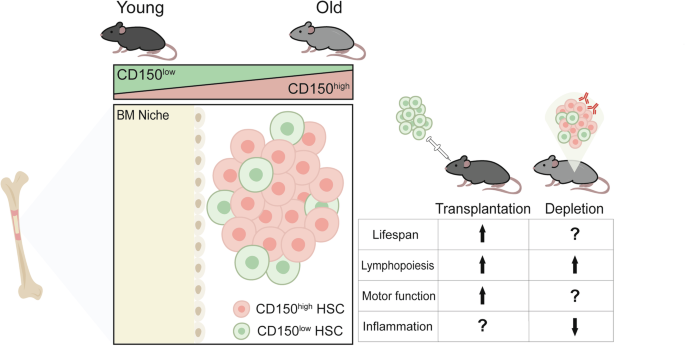
Fig. 1: CD150low HSCs in old mice possess youthful molecular profiles and mitigate aging phenotypes.
Two molecularly distinct subpopulations of HSCs, distinguished by variable CD150 expression, reside in the aging bone marrow (BM) niche. In aged mice, enrichment of CD150low HSCs (green) by CD150low HSC transplantation or antibody-based CD150high HSC (red) depletion results in increased lifespan, lymphopoiesis, motor function, and decreased inflammation.
At the molecular level, scRNA-seq analysis revealed distinct transcriptional signatures between young and old HSCs with old quiescent HSCs possessing more heterogeneity, while young quiescent HSCs displayed uniform molecular profiles. Three distinct clusters emerged from old HSCs with two clusters expressing transcriptional signatures of aging, while the third cluster expressed a transcriptional signature reminiscent of young HSCs, mostly related to cell proliferation processes.
To detect this more youth-resembling HSC subpopulation, Wang et al. identified genes highly upregulated with age that display cell surface expression, of which CD150 emerged as a promising candidate. Previous work identified CD150 as a marker of myeloid-biased HSCs, which clonally expand with age.7 To validate CD150 as an HSC aging marker, FACS-based approaches were used to divide HSCs into four groups according to CD150 levels. CD150 level positively correlated with previously described HSC aging markers and negatively correlated with highly expressed genes in young HSCs. Functionally, CD150low HSCs derived from old donor mice showed superior engraftment in primary and secondary competitive transplantation assays compared to CD150high HSCs, displaying a more resilient HSC phenotype.
To characterize the proliferative and differentiation capacity of CD150low HSCs, the authors conducted serial transplantation of CD150low HSCs and CD150high HSCs at varying ratios. Even in recipients where CD150low HSCs were transplanted in lower proportions, this subset demonstrated superior engraftment and reconstitution of B cells and naïve T cells. Additionally, a higher frequency of myeloid cells was observed in the peripheral blood following transplantation of CD150high HSCs, consistent with the fact that old HSCs exhibit myeloid-biased potential. Most strikingly, CD150low recipient mice outperform their CD150high counterparts in lifespan and various well-being measures of health, such as motor function, suggesting that removal of these CD150high HSCs in favor of CD150low HSCs may attenuate age-related changes at old age.
While these data excitingly point toward targeting defective CD150high HSCs as a promising approach to counter aging in the elderly, further investigations will help determine the extent of its therapeutic potential. Transplantation experiments were performed in middle-aged recipient mice. The results showed that reducing the CD150high HSC population can mitigate, or perhaps delay, age-related phenotypes. However, it remains an open question whether this treatment can rejuvenate an aged animal late in life (i.e., turn back the clock). Performing a broader assessment of aging-related changes — including systemic inflammation and dysfunction in various tissues, such as brain aging — will help determine how robust the therapeutic potential is of targeting CD150high aged HSCs. Studies are also necessary to understand how altering subpopulations of HSCs impacts systemic aging. Specifically, what are the lineage cell types that are promoting age-related phenotypes and how do those cells communicate with tissues to impact their function with age?
Identifying approaches to target dysfunctional aged HSCs in humans will be critical for the translational potential of these findings. Whether CD150low HSCs are resilient to age-related changes in humans has not been established. Moreover, work is needed to identify the best strategy to remove dysfunctional aged HSCs. A recent study targeted myeloid-biased HSCs in mice for immune system rejuvenation using an antibody cocktail, including anti-CD150,5 raising the possibility of leveraging this antibody cocktail approach to mitigate systemic aging. Of note, targeting CD150-positive cells with this antibody-based approach is not without potential adverse effects, as CD150 was initially identified as a general marker of LT-HSCs.8 In addition to CD150, other candidate markers of dysfunctional aged HSCs (e.g., Selp and Clca3a1) have been identified.9,10 As such, combinatorial approaches targeting multiple cell surface proteins in concert should be explored to selectively target dysfunctional aged HSCs. Addressing these remaining questions will be critical for realizing the exciting prospect of systemic rejuvenation in the elderly by targeted removal of dysfunctional HSCs.
留言 (0)