
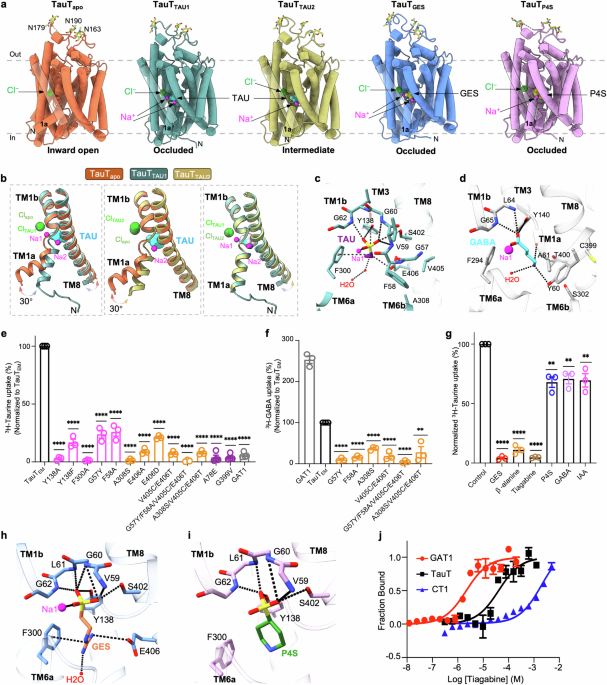
記住我
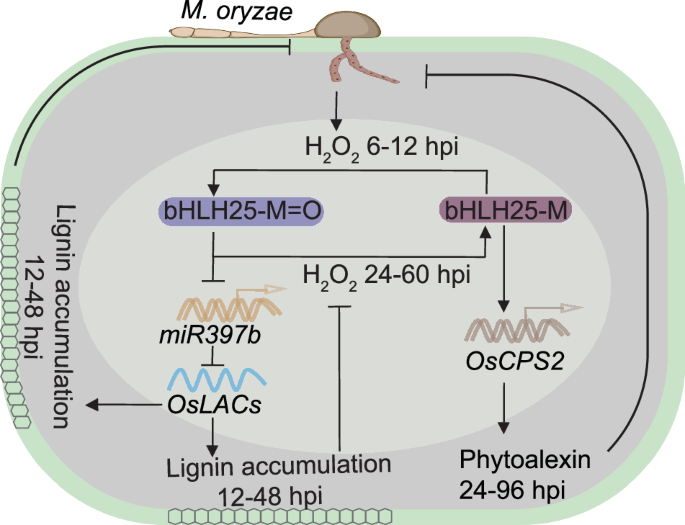



A recent study by Liao et al. reveals a sophisticated mechanism in rice where the transcription factor bHLH25 senses H₂O₂, orchestrating two independent defense pathways: lignin biosynthesis for physical reinforcement and phytoalexin production for antimicrobial action.
Hydrogen peroxide (H₂O₂) plays a critical multifaceted role in plant defense mechanisms in response to biotic and abiotic stresses.1,2 Pathogen attacks induce a rapid accumulation of reactive oxygen species (ROS), including H₂O₂ which serves as a frontline defense mechanism that can restrict pathogen growth through two primary pathways: directly damaging pathogen cellular components, such as DNA, proteins, and membranes,3 and indirectly reinforcing plant defenses by strengthening cell walls through cross-linking cell wall proteins.4 Furthermore, H₂O₂ acts as a secondary messenger in the activation of systemic acquired resistance (SAR)5 after a localized immune response mediated by the nucleotide-binding leucine-rich repeat (NLR) immune receptors,4 amplifying the plant’s defensive signaling network.
In this recent study, Liao et al.6 demonstrated that bHLH25, a basic/helix-loop-helix transcription factor in rice, senses H₂O₂ through the oxidation of a conserved methionine residue (M256). This oxidation triggers dual defensive responses: lignin biosynthesis via suppressing transcription of miR397b, a microRNA that specifically targets and negatively regulates the mRNA levels of the laccase enzyme (LAC) genes, OsLAC7, OsLAC28, and OsLAC29, and phytoalexin production through activation of Copalyl Diphosphate Synthase 2 (CPS2) (Fig. 1). In the context of response to the rice blast pathogen Magnaporthe oryzae, inoculation induced H₂O₂ production, leading to increased M256 oxidation of bHLH25. This oxidation suppresses miR397b, leading to increased expression in OsLAC7, OsLAC28, and OsLAC29 crucial for lignin polymerization. The resulting lignin accumulation reinforces plant cell walls, preventing pathogen entry and enhancing disease resistance.7,8 Notably, H₂O₂-mediated disease resistance through bHLH25 is only partially dependent on miR397b because overexpression of bHLH25 in rice led to a cell death phenotype even in the absence of pathogen attack, a response not observed in the miR397b knockout plants. Beyond the lignin biosynthesis, bHLH25 was shown to enhance disease resistance by promoting CPS2-mediated biosynthesis of phytocassane, a phytoalexin known to inhibit M. oryzae spore germination and trigger cell death.9 Furthermore, the rice CPS2 promoter contains an N-box-like motif, which is a binding site for bHLH25. The study confirmed that bHLH25 directly activates rice CPS2 expression, driving phytoalexin accumulation and further strengthening disease resistance.
Fig. 1: The working model.
M. oryzae infection in rice leaves results in the production of H2O2 at 6‒12 hpi, which oxidizes bHLH25 at M256, promoting OsLAC7/28/29 expression and lignin accumulation to prevent pathogen penetration through repression of the microRNA, miR397b. Subsequently, the increased lignin decreases H2O2 level at 24‒60 hpi, which inhibits oxidation of bHLH25, and non-oxidized bHLH25 promotes accumulation of phytoalexin to further increase rice defense through increased expression of OsCPS2.
Temporally, following M. oryzae inoculation, H₂O₂ level increased at 6‒12 h post inoculation (hpi), resulting in the oxidation of bHLH25 at 24 hpi. This oxidation suppressed miR397b and subsequently induced the expression of genes, OsLAC7, OsLAC28, and OsLAC29, promoting lignin accumulation at 12‒60 hpi as part of the early defense response. As the lignin biosynthesis progressed, H₂O₂ levels gradually declined after 24 hpi. This reduction in H₂O₂ allowed the accumulation of non-oxidized bHLH25, enabling it to activate CPS2 expression at 24‒96 hpi, which enhanced phytoalexin accumulation. Furthermore, H₂O₂-mediated disease resistance is primarily dependent on the repression of miR397 rather than the activation of CPS2. This conclusion is supported by observations that H₂O₂-induced disease resistance was significantly compromised in miR397b-overexpression rice plants, whereas it remained unaffected in the CPS2 knockout or overexpression plants.
Additionally, the methionine residue M256 is highly conserved among bHLH25 orthologs across many plant species. The in vitro study demonstrated that the conserved methionine in Arabidopsis AtbHLH25 can also be oxidized by H₂O₂. In rice, bHLH25-mediated disease resistance is localized to pathogen infection sites, which showed high H₂O₂ accumulation. However, under conditions of weak H₂O₂ induction, such as salt or high-temperature stress, the oxidation level of bHLH25 remains unchanged, resulting in no significant effect on rice resistance. Beyond its role in defense against M. oryzae, bHLH25 also confers rice resistance to other pathogens, including Rhizoctonia solani and Xanthomonas oryzae pv. Oryzae, underscoring its broader role in plant immunity.
The discovery of bHLH25 as a H₂O₂ sensor that integrates dual defense pathways represents a major advancement in plant immunity research. By linking lignin biosynthesis and phytoalexin production to pathogen-induced oxidative signals, bHLH25 provides crucial insights into how plants mount localized defense responses. However, the finding that bHLH25 knockout rice plants exhibit compromised but not completely abolished resistance suggests that additional H₂O₂-mediated defense mechanisms remain to be uncovered. Given that H₂O₂ can oxidize amino acids such as cysteine, methionine, tyrosine, and tryptophan,10 it would be intriguing to explore whether residues beyond M256 also play roles in bHLH25-mediated defense. Furthermore, as methionine oxidation can be induced by various molecules, including ROS and reactive nitrogen species (RNS), determining whether H₂O₂ directly oxidizes bHLH25 in planta or acts indirectly through downstream molecules remains an exciting avenue for future research.
Notably, prior studies have identified CCA1 Hiking Expedition/TCP21 (CHE) as another transcription factor that senses H₂O₂, but only in systemic tissues after pathogen attack.5 CHE’s conserved cysteine residue is sulfenylated by H₂O₂, enhancing its binding to the salicylic acid (SA) synthesis gene ICS1 to promote SAR. This raises the compelling question of whether bHLH25, in addition to its roles in lignin and phytoalexin pathways, may also contribute to defense via the SA signaling pathway. Another intriguing area for exploration is how plants regulate H₂O₂ production to coordinate defense responses across different tissues. While bHLH25 detects high H₂O₂ concentrations in pathogen-infected regions, CHE responds to lower H₂O₂ levels in uninfected systemic tissues. Investigating the mechanisms underlying this spatial regulation of H₂O₂ could shed light on how plants balance localized and systemic defense responses. Finally, both the bHLH25 and CHE studies highlight the pivotal role of NADPH oxidases (RBOHs) in pathogen-induced H₂O₂ production and defense signaling. The functional diversity of RBOHs in both biotic and abiotic stress responses deserves further investigation to better understand their contributions to plant resilience under diverse environmental conditions.
留言 (0)