
記住我




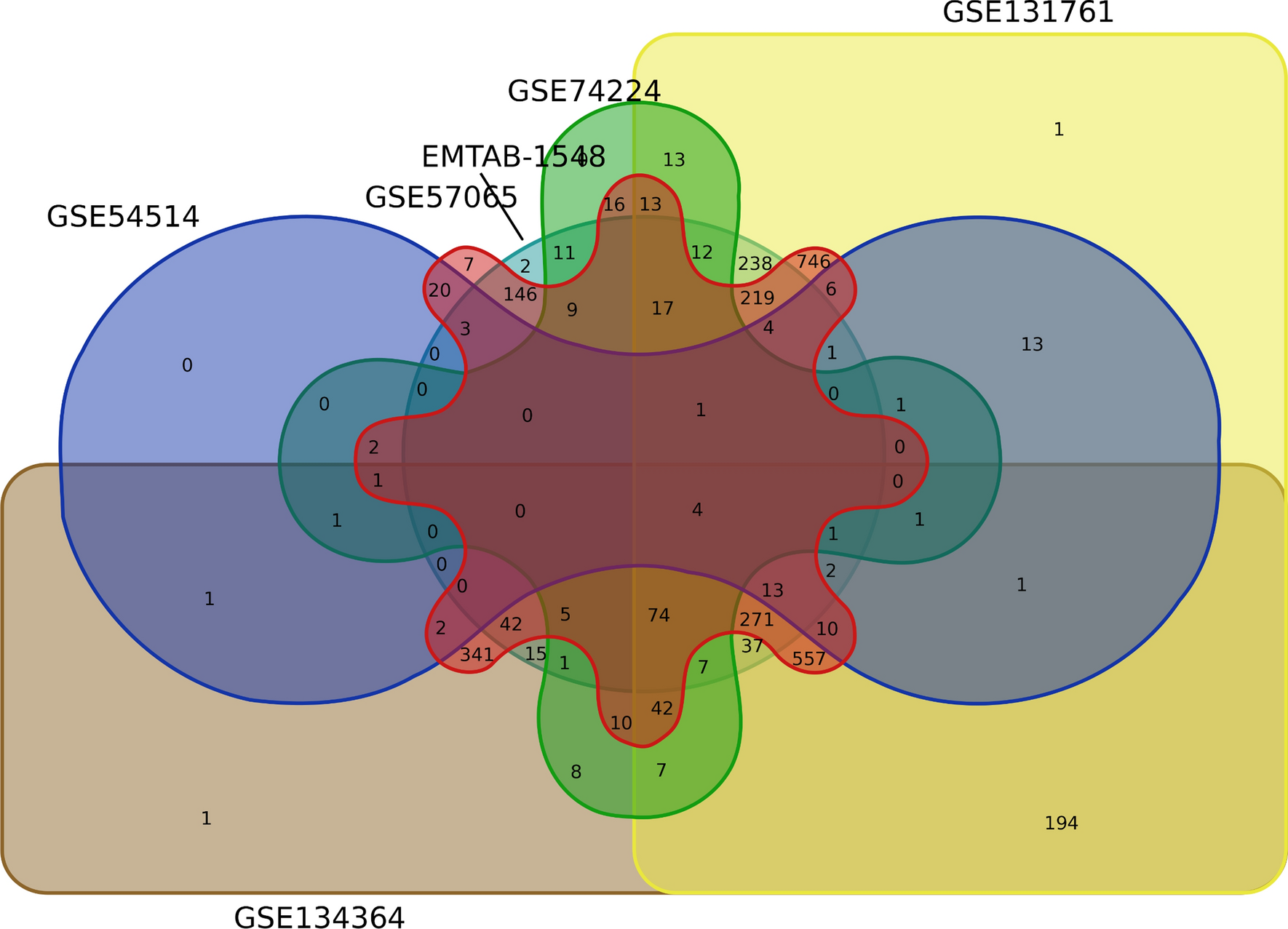

Sepsis is found to be one of the fatal conditions with high morbidity and mortality in susceptible age groups. It is associated with inflammation [15], immunosuppression [16], immune dysfunction [17] and increased mortality [18]. The pathophysiology of the sepsis is not well established, and the literature survey has disclosed vast heterogeneity among the studies in relation to gene expression in sepsis. Heterogeneity in transcriptome is known fact while comparing different sample types, for instance PBMC vs Whole blood. Hence our meta-analysis was limited to transcriptome data derived solely from whole blood. From the Venn diagram (Fig. 1), the analysis showed a significant number of common genes with four, 74, 271, 557, and 746 genes in six, five, four, three, and two datasets, respectively. Lesser common genes were observed upon analysis of six or five datasets due to low DEGs in two datasets (GSE54514 and GSE74224). The other four studies showed a significant number of common genes resulting in less heterogeneity.
Analysis of six datasets with GEO2R resulted in 233 common DEGs; 147 genes were up-regulated and 87 were downregulated with p-value < 0.05 and log2 FC ≥ 1.5. The DEGs were subjected to network and functional enrichment analysis to identify the hub genes, GO terms, and KEGG pathways. It was observed that several genes involved in adaptive and innate immune response were dysregulated, therefore interfering with normal immune function. Based on the Venn diagram (Fig. 1) four genes HK3, ITGA2B, MPO, and TLR5 were found to be up-regulated in all six studies. Hexokinase-3(HK3) and myeloperoxidase (MPO) are involved in neutrophil activation, regulation and degranulation. MPO is an enzyme present in the granules of polymorph nuclear (PMN) leucocytes associated with the formation of reactive oxygen species (ROS) and oxidation of biological material. Elevated neutrophil levels during sepsis increases MPO in the plasma, which could be used as a biomarker of neutrophil activity and inflammatory response during sepsis[19, 20]. HK3 is found in the HIF-1 signaling pathway that mediates glycolysis in neutrophils, also influencing increased PMNs chemotaxis and phagocytosis function [21]. ITGA2B is involved in regulation of hemopoiesis, hemostasis, coagulation, leukocyte migration, platelet activation and degranulation. Increased expression of ITGA2B in circulating platelets during sepsis is associated with increased mortality, which could be used as a potential marker to predict mortality [22]. TLR5 is associated with defense response to bacterium, positive regulation of IL-8 and other cytokines. It is a core member of the Toll -like receptor family that induces the inflammatory cytokine production and immune response [23].
In our study we found five MCODE clusters, in which three clusters (Fig. 4a–c) consisting of 45 genes in total were found to be significant. Cluster-1 (20 genes) is mainly involved in regulation of adaptive immune system, T cell differentiation and development. All genes in cluster-1 were down-regulated resulting in suppression of T cell function, suggesting immunosuppression during sepsis. Cluster-2 (10 genes) and 3 (15 genes) participate in innate immune responses, pathogen defense, antigen processing and presentation, neutrophil regulation and NET formation. Interestingly, all except three genes (CD74, HLA-DMA, and HLA-DMB) were up-regulated in cluster 3. The highest scoring genes KLRB1 (killer cell lectin-like receptor B1), DEFA4 (defensin 4), and LTF (lactoferrin) in clusters one, two, and three, respectively. KLRB1 plays a role in innate immune response, mediates cytotoxicity, and secrete cytokines upon stimulation [24]. DEFN4 found abundant in neutrophil granules, exhibits antimicrobial activity against gram-negative bacteria [25]. LTF is reported as the marker for sepsis in several studies, it plays a key role in innate immune response and pro-inflammatory cytokine production through the TLR4 signaling pathway [26]. Through network and functional enrichment analysis, 20 hub genes were identified that are involved in defense response against microbes, adaptive and innate immune response, regulation of Tcell and neutrophil differentiation and activation. Also these genes have significantly higher fold change and were found to be part of enriched GO terms (Fig. S1a) and KEGG pathways (Fig. S1b), which could be identified as key genes associated with sepsis conditions. Meta-analysis revealed that the adaptive immune response genes (CD3G, CD247, CD3E, CX3CR1, IL7R, CCR7, CD2, CXCR3, LCK, CD8A, and CD40LG) that regulate T cell functions were down-regulated, which could be possible cause of suppressed immune response. The genes involved in innate immune response and neutrophil function (CD177, S100A12, ARG1, FCER1G, ELANE, S100A8, S100A9, ANX1, CTSG, IL18RAP, and HP) were up-regulated during sepsis. Targeting the cluster genes and corresponding pathways could lead to discovery of biomarkers associated with pathology of sepsis.
Almansa et al., reported a similar pattern where expression of significant 55 genes involved in adaptive and innate immune response are associated with organ failure and mortality in sepsis [12]. In relation to this the genes associated with markedly enriched GO terms—innate immune response, defense response to bacteria and fungus, neutrophil activation and NET formation were up-regulated. Adaptive immune response, antigen processing and presentation, Immunoglobulin production, T cell differentiation, selection, activation and regulation were found to be down-regulated. Similar results were observed in KEGG pathway analysis, where innate immune response related pathways are up-regulated and pathways related to adaptive immune response were down-regulated (Fig. 5).
Fig. 5
Image credits: Shruthi T N; Dr Anbazhagan K
Altered Innate and Adaptive Immune System during Sepsis: The interaction between host and pathogen (PAMP-PRR interaction) leads to the activation of innate immune cells such as neutrophils, macrophages and recruit to the site of infection. Activated macrophages and neutrophils release the pro-inflammatory cytokines (IL1R1, IL6, INFG, TNF etc.…) and chemokines to the site of infection leading to cytokine storm. The neutrophils show decreased apoptosis and enhanced mobilization leading to increased ROS and NET formation, which further aggravates the inflammatory mediators resulting in tissue damage and hyper-inflammatory response. Activated innate immune cells further activate the adaptive immune cells like T cells, B cells and NK cells. The T cells show decreased activation, proliferation, antigen presentation and increased apoptosis during sepsis due to suppressed anti-apoptotic genes and enhanced apoptotic gene expression resulting in immunosuppression. Together these hyper inflammatory responses and Immune suppression may lead to multiple organ failure or death. Down arrow in green indicates the suppressed function; up arrows with red indicate enhanced function. Genes in red represent increased expression; genes in blue represent decreased expression.
Among all immune cells neutrophils play a crucial role in innate immunity, acting as the first line of defense in the clearance of pathogenic bacteria during infection. Neutrophils play a key role in controlling infection during normal conditions, but their antimicrobial activity is impaired and immune responses are dysregulated during sepsis [27]. They participate in chemotaxis, phagocytosis, oxidative burst, and neutrophil extracellular traps. Upon pathogen encounter the host recognizes the PAMPs by pathogen recognition patterns (PRRs-TLR2, TLR4, TLR5, TLR8), which induce cytokine storm. The pro-inflammatory and anti-inflammatory cytokines, chemokines, interleukins and TNF released from macrophage (IL1R1, IL1R2, IL18R1, and IL18RAP), B cells and T cells acts on itself or other cell types like neutrophils and endothelial cells. These cytokines further activate the NFKB pathways and JAK/STAT signaling to recruit more pro-inflammatory cytokines to the site of infection to clear the pathogen. But in case of sepsis the dysregulated immune system results in release of a large pool of pro-inflammatory cytokines and adhesion molecules which leads to hyper-activation of neutrophils further resulting in hyper-inflammatory response. Increased circulation of neutrophils is observed in sepsis condition, which further infiltrates and accumulates in tissues leading to tissue damage and multi-organ failure [28, 29].
In addition neutrophils exert their function through the formation of NETs, which is a mechanism to capture and kill the pathogen extracellularly. During sepsis excessive NETosis is shown to be detrimental to the host. This NETosis induces histone dependent vascular damage leading to tissue damage and multiple organ dysfunction [30]. The neutrophil genes related to NETosis were found to be highly up-regulated signifying their association with increased inflammation in sepsis. The life span of circulating human neutrophil is estimated to be 5.4 days approx. But, in case of sepsis, the cell count is increased due to delayed apoptosis of neutrophils, thus increasing the lifespan and altering the phenotype and functional properties of neutrophils. The suppression of neutrophil apoptosis may be due to alteration in the apoptotic pathways and altered expression of caspase (Caspase4, Caspase3, and caspase 5) activity as observed in the meta-analysis. Thus the altered neutrophil function like phagocytosis, chemotaxis and apoptosis results in hyper-inflammatory response.
T cells are an essential part of the immune system that fight against the pathogen to protect the host. Several studies reported depletion of T cells leading to reduced adaptive immunity and prolonged lymphopenia in sepsis patients. A study reported a decreased number of CD4 + and CD8 + T cells in both adult and elderly sepsis patients. The CD28 expressing CD4 + T cells were reduced, which is known to be immunocompetent, while immunosuppressed PD-1 + T cells were increased [31]. From the analysis we found PD-L1 (CD274) was up-regulated in five different studies, which could be correlated to reduced T cell proliferation and increased apoptosis thus affecting their function. This could lead to increased secondary infections, multiple organ dysfunction and mortality in sepsis and septic shock [32, 33]. Patil et al., have reported that PD-L1 knockout mice showed improved function of T cells and survival [34], thus indicating its key role in T cell exhaustion and pathogenesis of sepsis.
Several studies have reported the expression of PD-L1 (CD274) on neutrophils in various disease conditions stimulated by interferon-gamma (IFN-G) and granulocyte macrophage-colony stimulating factor (GM-CSF) [35, 36]. Kleijn et al., reported the stimulation of neutrophils with IFN-G ex-vivo induces PD-L1 expression on neutrophils, which suppress the lymphocyte proliferation and induce the apoptosis of T cells leading to immunosuppression [37]. Upregulation of ARG1 on neutrophils in sepsis worsen the immunosuppression by suppressing T-lymphocytes. It impairs the function of T cells by down-regulating T cell receptor (TCR) associated CD3G and CD3E chains leading to immunosuppressive state [38].
IL7R is an important cytokine related gene found to be involved in hematopoietic cell lineage pathway that is down-regulated in sepsis, which is an important gene for maintaining the homeostasis of immune cells. It plays a predominant role in the growth, differentiation and survival of T cells. In addition to IL7R, CD27 is also involved in protecting T cells from apoptosis, thus contributing for survival and expansion of T cell [39, 40]. Reduced expression of IL7R and CD27 is observed in current analysis, and this could be associated with impaired differentiation of T cells. This is one of the causes for immunosuppression and enhanced susceptibility to infections during sepsis [41]. In addition to this, increased cell death during sepsis releases excess amounts of DAMPs, neutrophil extracellular traps (NETs) which trigger host immune response leading to increased inflammatory reaction. Hence DAMPs play a prominent role in sepsis progression [42]. S100 A8/A9/A12, types of DAMPs which are increased in sepsis condition and are associated with increased severity and death of sepsis patients [43]. The genes which are responsible for balanced functioning of T-cells (CD8A, IL7R, IL2 and CD28) and Tcell surface markers (CD3) are down-regulated. Increased expression of PD-L1 (CD274) and apoptotic genes could result in immune dysfunction, reduced functioning of T cells and increased cell death, which may lead to increased severity of sepsis and mortality.
The hyper-inflammation response during sepsis could be due to the suppression of apoptosis in circulating neutrophils, and their accumulation in tissues lead to irreversible tissue damage and multiple organ failure. Immune suppression is induced by depletion of immune cells, specifically T cells during sepsis. Overexpression of apoptotic genes and down-regulation of anti-apoptotic genes have potent effects on T cell apoptosis. Increased T cell apoptosis and neutrophil survival aggravates the immune suppression and inflammatory reaction, respectively. Exploring apoptotic and inflammatory genes could be potential targets to reverse immunosuppression and identify valuable biomarkers. In this study, through functional enrichment analysis, we found 20 candidate genes (Table 4) known to play a potential role in pathophysiology of sepsis. These genes include immune cell markers, secretory markers, interleukins and chemokines, further research and validation may result in understanding their expression at various stages of sepsis, which could be clinically useful as diagnostic and prognostic markers.
留言 (0)