
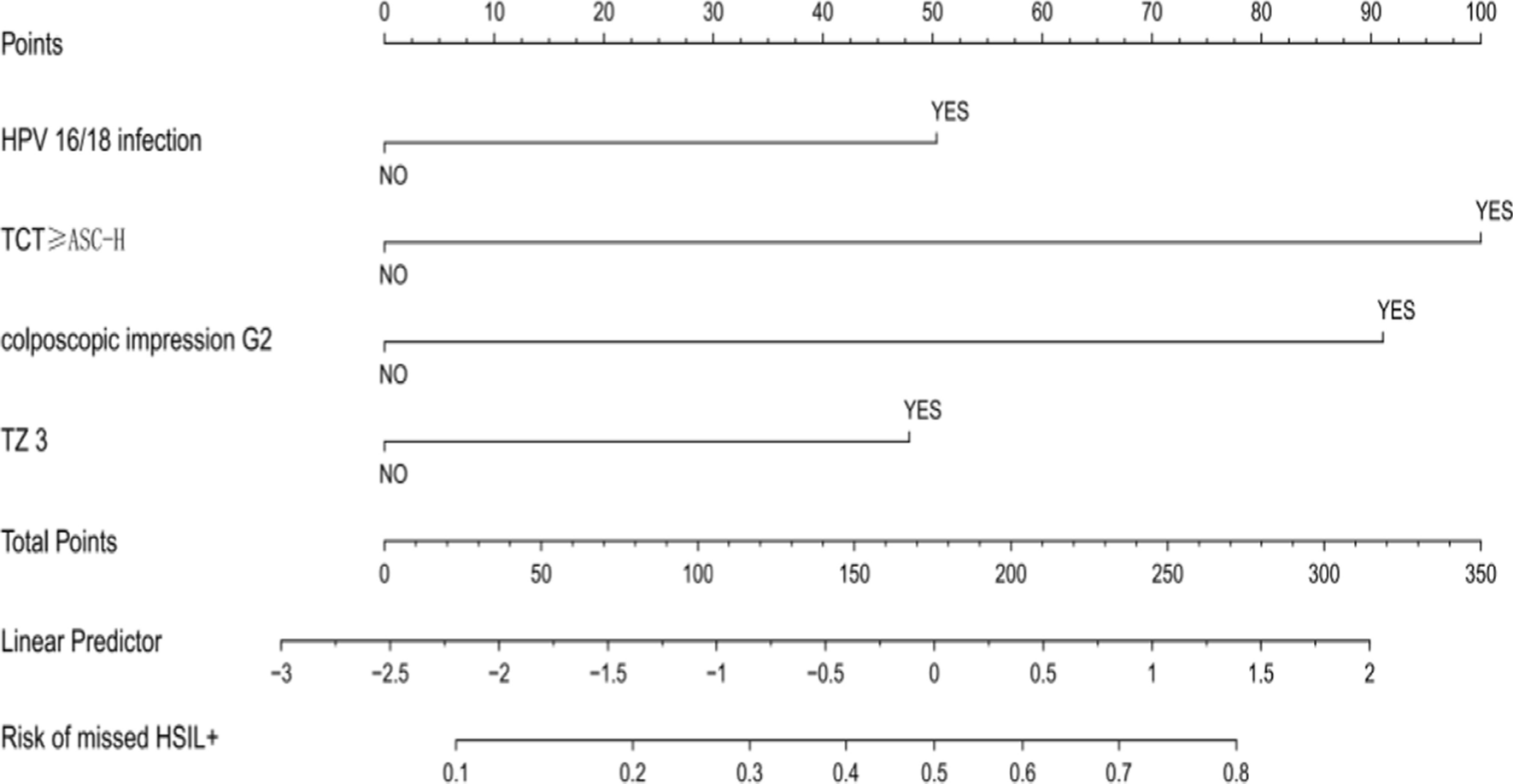
記住我



In the current study, G-negative bacteria were the most prevalent bacteria in the obtained microbiome with the rate of 74.87% (146/195 isolates, Table 1). Furthermore, data in Table 1 showed that the general ranking of the most common pathogens among G-negative microbiota of oncology patients was Esherichia sp. (n = 36, 24.66%) > Acinetobacter sp. (n = 34, 23.29%) > Stenotrophomonas sp. (n = 29, 19.86%) > Pseudomonas sp. (n = 26, 17.81%) > Serratia sp. (n = 21, 14.38%).
Table 1 Multidrug resistance prevalence and MICs of various antibiotics against G-negative bacterial strains derived from the cancer mirobiomeThe prevalence of antibiotics resistance to G-negative bacteria obtained from cancer microbiomeThe antibiotic resistance prevalence analysis in Table 1 showed that the obtained G-negative bacteria derived from the cancer microbiome exhibited entirely multidrug resistance to penicillin antibiotics including ampicillin, ticarcillin, oxacillin, piperacillin and carbenicillin (MIC; 128—> 1024, 560—> 1024, 256—> 1024, 512—> 1024 and 440—> 1024 μg/mL), respectively as well as the first-generation cephalosporins including cephalothin, cefazolin and cefoxitin (MIC; 256—> 1024, 360—> 1024 and 480—1024 μg/mL), respectively. Moreover, resistance prevalence to other cephams including ceftazidime, cefepime, ceftriaxone and cefotaxime reached 100% in Stenotrophomonas and Acinetobacter isolates with MIC ranged from 128 to ˃ 1024 μg/mL (Table 1). Additionally, resistance rate equal to 96.15, 76.92, 65.39 and 57.69% with MIC 400–664, 256–512, 128–512 and 64–512 μg/mL, respectively was recorded in Pseudomonas isolates (n = 26) compared to 95.24, 76.19, 66.67 and 61.91% with MIC 512–720, 256–640, 128–560 and 64–512 μg/mL, respectively in Serratia sp. isolates (n = 21 isolates) while 88.89, 80.56, 63.89 and 63.89% of Esherichia isolates (n = 36 isolates) were resist to ceftazidime, cefepime, ceftriaxone and cefotaxime with MIC 256–512, 128–800, 64–480, and 128–256 μg/mL, respectively (Table 1). Then the cancer microbiota under study can be considered as extended-spectrum β-lactamase producers (ESBL).
The highest resistance rates toward imipenem (73.08% with MIC; 128–512 μg/mL) and doripenem (76.92% with MIC; 264–472 μg/mL) was recorded among Pseudomonas species but the highest resistant against meropenem (75.86% with MIC; 256–600 μg/mL) and ertapenem (79.41% with MIC; 400–648 μg/mL) were occurred among Stenotrophomonas and Acinetobacter species, respectively (Table 1). On the other hand, when ceftazidime, imipenem and meropenem were used together with the β-lactamase inhibitors avibactam, cilastatin and vaborbactam, respectively the rates of resistance in the total number of G-negative bacteria decreased significantly from 95.89, 62.33 and 68.49% to 39.73, 25.34 and 29.45%, respectively (Table 1).
Moreover, data in Table 1 showed that the rates of resistance toward fluoroquinolones antibiotics include ofloxacin, ciprofloxacin levofloxacin and moxifloxacin as well as trimethoprim/sulfamethoxazole (folate synthesis pathway inhibitors) were ≥ 61.91% ≤ 80.95%; ≥ 41.67% ≤ 75.00%; ≥ 69.23% ≤ 80.77%; ≥ 75.86% ≤ 93.10%, and ≥ 73.53% to 100.00% with Serratia sp., Esherichia sp., Pseudomonas sp., Stenotrophomonas sp. and Acinetobacter sp., respectively. Because aminoglycosides remain, a significant class of agents when microorganisms of interest are resistant to other classes, especially β-lactams and fluoroquinolones as in the current study, resistance to aminoglycosides among G-negative microbiome under study were evaluated (Table 1). High resistance potency was recorded against nitrofurantoin (n = 93, 63.70%) followed by tobramycin (n = 90, 61.64%), netilmicin (n = 85, 58.22%), gentamicin (n = 82, 56.16%), amikacin (n = 69, 47.26%) and vancomycin (n = 57, 39.04%) (Table 1). Therefore, the current spread of resistance to aminoglycosides amongst already MDR microorganisms is an unwelcome event. The data in Table 1 indicated that the cancer microbiome resistance rates under study were estimated to be 63.70, 42.47 and 34.93% with MIC; 240–1000, 128–500, and 32–280 μg/mL against the tetracycline antibiotics (doxycycline, minocycline and tigecycline), respectively but 79.45, 74.66 and 65.07% of G-negative isolates were resistant to clarithromycin, zithromycin (macrolides) and chloramphenicol, respectively with MIC ranged between 128 and > 1024 μg/mL. Of the results presented in Table 1, the highest sensitivity profile was recorded against colistin followed by imipenem-cilastatin, meropenem-vaborbactam, tigecycline, vancomycin and ceftazidime-avibactam (78.77, 74.66, 70.55, 65.07, 60.96 and 60.27%), respectively (Table 1). The isolates Serratia sp. Esraa 1, Stenotrophomonas sp. Esraa 2, Acinetobacter sp. Esraa 3, Escherichia sp. Esraa 4 and Pseudomonas sp. Esraa 5, which showed resistance against all the antibiotics under study and then characterized extensive drug-resistant (XDR) strains with the highest MIC values, were selected for the further studies.
Carcinogenic activities of bacterial extractsAfter oral ingestion of microbial extracts, under study by the five animals’ groups, the values of serum cancer markers (AFP, CEA, and LDH) increased significantly in all treated animals’ groups comprised to the healthy animals’ group (Table 2). Serum AFP levels was significantly increased from 0.52 ± 0.019 ng/mL (healthy group) to 2.08 ± 0.07 ng/mL (300%) after treating with Acinetobacter sp. Esraa 3 followed by Pseudomonas sp. Esraa 5, Stenotrophomonas sp. Esraa 2, Escherichia sp. Esraa 4, and Serratia sp. Esraa 1 extracts (1.92 ± 0.41; 269.2%, 0.85 ± 0.04; 63.5%, 0.73 ± 0.04; 40.4%, and 0.57 ± 0.02 ng/mL; 9.6%), respectively (Table 2). Furthermore, serum CEA level increased in descending order in animals’ groups handled by extracts of Pseudomonas sp. Esraa 5, Stenotrophomonas sp. Esraa 2, Escherichia sp. Esraa 4, Acinetobacter sp. Esraa 3, and Serratia sp. Esraa 1 (07.69 ± 0.71; 311.2%, 6.08 ± 0.13; 225.1%, 4.41 ± 0.79; 135.8%, 3.53 ± 0.07; 88.8%, and 1.99 ± 0.14 ng/mL; 6.4%), respectively compared with the control group (1.87 ± 0.04 ng/mL), indicating the carcinogenic activity of the studied G-negative bacteria (Table 2). Moreover, activity of serum lactate dehydrogenase (LDH) significantly increased to 2248 ± 81.1, 1851 ± 73.4, 789 ± 7.49, 623 ± 18.5, and 402 ± 11.6 IU/L (464.8, 365.1, 98.2, 56.5, and 1.1%) in rats after oral ingestion of the extracts of strains Esraa 3, Esraa 5, Esraa 2, Esraa 4, and Esraa 1, respectively comprised with 398 ± 3.74 IU/L in healthy animals’ group (Table 2). For instant, the higher carcinogenic activity with Escherichia strain was achieved by subcutaneous-injection while Serratia sp. Esraa 1 or Pseudomonas sp. Esraa 5 extracts markedly increased serum level of CEA in the case of oral ingestion. However, there is no significant difference in the effect of Stenotrophomonas sp. Esraa 2 extract whether administered orally or by injection while Acinetobacter sp. Esraa 3 extract increased serum LDH activity by 171.5 and 98.2% post subcutaneous-injection and oral administration, respectively.
Table 2 Effect of oral ingestion of selected bacterial extracts on some tumor markers in treated albino rats’ groups compared to the normal groupHepatotoxic effect of bacterial extractsOral ingestion of the microbial extracts in treated animals stimulated liver dysfunction that was monitored through increased activities of liver enzymes. ALAT activity markedly increased from 23.1 ± 1.8 U/L (control group) to 27.1 ± 1.98, 85.3 ± 6.56, 27.7 ± 2.06, and 120.6 ± 3.2 U/L (17.3, 269.3, 19.9, and 422.1%) in the animal groups ingested with the extracts of strains Esraa 2, Esraa 3, Esraa 4, and Esraa 5, respectively; while it decreased by 7.8% post-handling with the extract of strain Esraa 1 (Table 3). Similarly, serum ASAT activity increased from 22.3 ± 0.3.6 U/L (control group) to 25.2 ± 4.1, 29.0 ± 3.8, 78.6 ± 9.6, 25.4 ± 4.12, and 101.7 ± 3.13 U/L (13.0, 30.0, 252.5, 13.9, and 356.1%); GGT increased from 45.6 ± 2.83 U/L (control group) to 52.1 ± 1.44, 64.8 ± 2.84, 142.1 ± 3.8, 48.7 ± 1.38, and 102.7 ± 13.19 U/L (14.3, 42.1, 211.6, 6.8, and 125.2%); as well as serum ALAP activity increased from 80.7 ± 4.38 (control group) to 93.9 ± 9.3, 138.3 ± 6.9, 375.4 ± 10.5, 89.7 ± 1.83, and 256.4 ± 7.43 U/L (16.4, 71.4, 365.2, 11.2, and 217.7%) after the oral ingestion of the extracts of Esraa 1, Esraa 2, Esraa 3, Esraa 4, and Esraa 5, respectively (Table 3). These results referring to sever hepatotoxic potential of these extracts; the highest hepatotoxic effect was exhibited by Esraa 3, and Esraa 5 (Table 3).
Table 3 Effect of oral ingestion of selected bacterial extracts on liver function enzymes in treated albino rats’ groups compared to normal animalsNephrotoxic effect of bacterial extractsThe extracts of cancer microbiota exhibited nephrotoxic effects on the kidneys, which were achieved through a respectable increase in the concentricity of urea and creatinine in the blood compared to normal rats. Esraa 3 extract performed the most nephrotoxic properties as it increased the serum levels of urea and creatinine to 110.4 ± 3.95 (253.9%), and 1.78 ± 0.42 mg/dl (114.5%), respectively; while the extracts of Esraa 1, and Esraa 2 exhibited the smallest effect as they performed the minimal increase in serum urea (34.7 ± 3.25 mg/dl, 11.2%) and creatinine (0.84 ± 0.07 mg/dl, 1.2%), respectively (Table 4). This study revealed that these tumors-associated microbial communities have some sort of influence on tumor development. Therefore, this correlation can be considered as demonstrated that it can be defined as an oncobiome.
Table 4 Effect of oral ingestion of selected bacterial extracts on kidney function tests and inflammatory cytokines in the treated albino rats’ groups compared to the normal animalsEffect of the bacterial extracts on the inflammatory cytokinesInflammatory cytokines (IL-1β and TNF-α) levels were markedly increased post-oral ingestion of each bacterial extracts under study. Pseudomonas sp. Esraa 5 extract induced the highest production of IL-1β followed by Esraa 2, Esraa 3, Esraa 4, and Esraa 1 extracts (30.2 ± 2.41; 297.4%, 24.5 ± 2.18; 222.4%, 18.6 ± 1.89; 144.7%, 15.3 ± 1.08; 101.3%, and 7.8 ± 1.44 ng/L; 2.6%), respectively compared to 7.6 ± 1.65 ng/L of control group (Table 4). Moreover, TNF-α level was increased by (11.08 ± 0.94; 6.5%, 29.8 ± 1.71; 186.5%, 27.5 ± 0.65; 164.4%, 19.1 ± 2.44; 83.7%, and 41.9 ± 5.32 ng/mL; 302.9%) after oral administration of Esraa 1, Esraa 2, Esraa 3, Esraa 4, and Esraa 5 extracts, respectively compared to 10.4 ± 0.24 ng/mL of control group (Table 4).
Histopathological studiesThe histopathological studies of the liver sections of untreated animals (group 1), and oral ingestion treated groups with Esraa 1 (group 2), Esraa 2 (group 3), Esraa 3 (group 4), Esraa 4 (group 5) and Esraa 5 (group 6) extracts, stained by Masson trichrom to examine the alterations in collagen and fibrous tissues are illustrated in Figs. 1 and 2. Liver section of the control group in Figs. 1 and 2 (section 1) showed branching cords of hepatocytes with vesiculated nuclei and intervening blood sinusoids lined with endothelial cells, hepatocytes as polygonal cells with acidophilic granular cytoplasm around the portal tract. The control group has the minimum amount of collagen or fibrous tissues compared to the collagen and fibrous tissues in all other orally treated groups. The hepatic sections of animals group 2 treated with the extract of Esraa 1 exhibited disrupted arrangement of hepatocytes around the portal tract (red arrow) together with severe cytoplasmic vacuolation and pyknotic nuclei (black arrow) but hepatocytes around the central vein are organized and nearly normal (blue arrow, Fig. 1; section 1P and 2; section 2). Treated animals by extract of Esraa 2 (group 3) exhibited disrupted arrangement of hepatocytes around the portal tract (pt) with vacuolization and pyknotic nuclei (arrow) but hepatocytes around the central vein are organized and nearly normal (inset) along with marked increase in connective tissue in liver capsule (right inset) were observed (Fig. 1; section 2P and 2; section 3). Radiating hepatocytes from the central vein, arranged, nearly normal and detached by blood sinusoid in addition to some inflammatory cells around the portal tract along with marked increase in connective tissue in liver capsule were observed in the liver sections of animals group 4 orally treated with Esraa 3 extract, (inset, Fig. 1; section 3P and 2; section 4). Data in Fig. 1 (section 4P) and 2 (section 5) illustrated the liver of animals group 5 with vacuolar and pyknosis in hepatocytes around the dilated and congested portal tract area with marked increase in normal arrangement of hepatic cells around the central vein (right inset) in connective tissue of liver capsule (left inset) after oral ingestion of Escherichia sp. Esraa 4 extract. Furthermore, the liver sections of the treated group with Esraa 5 extract (group 6) exhibited many hepatocytes with eosinophilic cytoplasm, pyknotic nuclei, massive increase in inflammatory cells around the dilated portal tract and central vein (left inset) and marked increase in connective tissue in liver capsule (right inset, Figs. 1; section 5P and 2; section 6).
Fig. 1
A section in liver of control and oral treated animals; (section 1; control) group 1 (Hx. & E. × 400), (section 1P) group 2 treated with Serratia sp. Esraa 1 extract (Hx. & E. × 400), (section 2P) group 3 oral treated with Stenotrophomonas sp. Esraa 2 extract (Inset × 400) (Hx. & E. × 400), (section 3P) group 4 treated with Acinetobacter sp. Esraa 3 extract (Hx. & E. × 300), (section 4P) group 5 treated with extract of Escherichia sp. Esraa 4 (Hx. & E. × 300), and (section 5P) group 6 oral treated with extract of Pseudomonas sp. Esraa 5 (Hx. & E. × 200)
Fig. 2
Photomicrographs of liver sections of 1; untreated and 2 −6; oral treated animal groups with extracts of Serratia sp. Esraa 1, Stenotrophomonas sp. Esraa 2, Acinetobacter sp. Esraa 3, Escherichia sp. Esraa 4 and Pseudomonas sp. Esraa 5 stained with Masson trichrom (Masson's trichrom reaction × 200), respectively
Frequency of micronuclei (MN) induced by the selected bacterial extracts in male ratsResults in Table 5 and Fig. 3a and b indicated that the ratio of micronuclei in poly chromatic erythrocytes from rat bone-marrow cells was statistically significant (p < 0.01) in all treated animals’ groups by oral ingestion compared to negative control. The numbers of MN cells in animals group that took orally the extract of Escherihia sp. Esraa 4 were markedly increased 10.32-fold (n = 392) followed by the animals’ groups that were administered orally extracts of Esraa 2, Esraa 5, Esraa 1, and Esraa 3 (8.45-fold; n = 321, 7.82-fold; n = 297, 7.79-fold; n = 296, and 5.97-fold; n = 227), respectively when compared to n = 38 in the untreated group (Table 5). Moreover, compared to untreated group (0.76 ± 0.48) the ratio of micro-nucleated polychromatic erythrocytes (MNPCEs) that specified by analyzing the number of MN cells from 1000 PCEs per animal was described to be 5.92 ± 0.58, 6.42 ± 0.50, 4.54 ± 0.63, 7.84 ± 0.68, and 5.94 ± 0.69 after oral ingestion by Esraa 1, Esraa 2, Esraa 3, Esraa 4, and Esraa 5 extracts in the current study (Table 5).
Table 5 Ratio of micronuclei in polychromatic erythrocyte and number of sperm defects stimulated in the treated male rats by selected bacterial extracts as compared to the normal rats groupFig. 3
Micronuclei in polychromatic erythrocyte prompted in rats’ bone marrow cells handled with the oral G-negative bacteria extracts; normal cell (a) and micronuclei cell (b)
Sperm-shape abnormalitiesVarious forms of head abnormalities as straight (n = , 34, 47, 40, 52, and 57), banana (n = 153, 161, 148, 157, and 191), amorphous (n = 41, 45, 37, 63, and 47) and without hook (n = 53, 39, 48, 61, and 31) in addition to the coiled tail (n = 20, 22, 18, 35, and 25) were discovered in groups treated by Esraa 1, Esraa 2, Esraa 3, Esraa 4, and Esraa 5 extracts, respectively compared to 27, 37, 3, 18, and 0, respectively in the untreated animals (Table 5 and Fig. 4a - d). The highest sperm abnormalities number in all groups was recorded for banana shape but the lowest was detected for coiled tail, that accompanied by male sterility (Table 5).
Fig. 4
Sperm defects stimulated in male rats treated by the oral G-negative bacteria extracts; normal sperm (a), amorphous (b), banana and without hook (c) and straight (d)
Assessment of anti-proliferative activities of the selected bacterial extracts against human normal cell linesAnti-proliferative activities of Esraa 1, Esraa 2, Esraa 3, Esraa 4, and Esraa 5 extracts individually against three different types of human normal cell lines including mammary epithelial (MCF10A), lung fibroblasts (WI38), and dermal fibroblasts (HDFs) are presented in Fig. 5. Total inhibition in the proliferation of WI38, MCF10A, and HDFs, when they were treated by the Serratia sp. Esraa 1 extract at concentrations of 60, 70, and 70 µg/mL and IC50 equal to 22.50, 26.80, and 32.28 µg/mL, respectively (Fig. 5). Stimulation of 100% death in WI38, MCF10A, and HDFs cells was noticed when they treated with Esraa 2 extract at doses of 50, 40, and 50 µg/mL with IC50 equal to 20.1, 12.80, and 11.20 µg/mL. Moreover, Escherichia sp. Esraa 4 extract at concentrations ranged between 70, and 80 µg/mL entirely inhibited the proliferation of WI38, MCF10A, and HDFs with IC50 ranged from 30 to 40 µg/mL, while their growth was totally inhibited at 40 µg/mL of Pseudomonas sp. Esraa 5 extract with 50% inhibition in their proliferation (IC50) achieved at doses of 20.0, 15.38, and 10.0 µg/mL, respectively. Interestingly, the extracted metabolites of Acinetobacter sp. Esraa 3 showed the highest cytotoxic effects against all human normal cells under study. Esraa 3 extract at a concentration of 30 µg/mL caused total inhibition in the growth of WI38, MCF10A, and HDFs cell lines with IC50 estimated to be 9.5, 14.12, 8.90 µg/mL, respectively (Fig. 5).
Fig. 5
Effect of the secondary metabolites extracted from the selected G-negative bacteria derived from cancer microbiota on the normal human cell lines MCF10A, WI38 and HDFs at different concentrations
留言 (0)