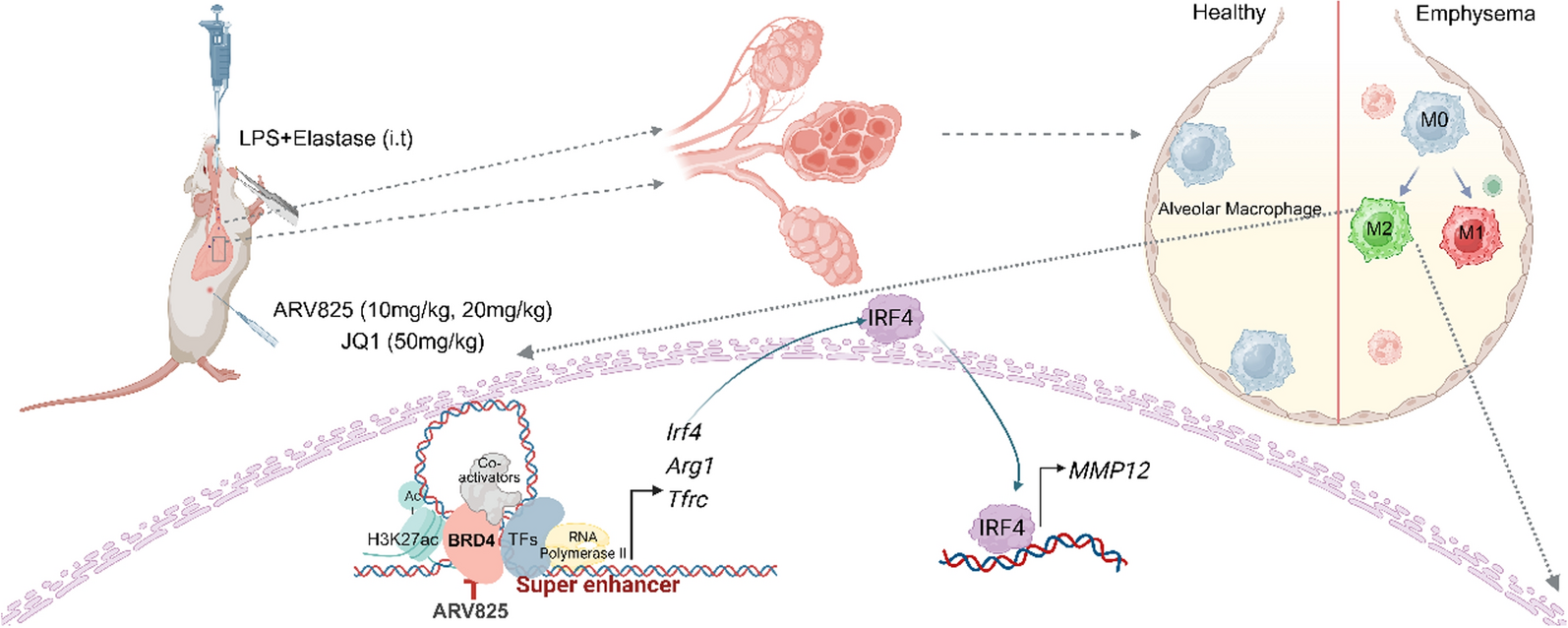
Increased MMP-12 in COPD
To determine the proteolytic activity in COPD airways, we measured concentrations of MMP-9, MMP-12, and TIMP-1 in BAL fluid from COPD subjects and controls. In line with many previous studies, we found that MMP-12 was significantly increased in COPD, but for MMP-9 and TIMP-1, no such associations were found.
Several mechanisms might contribute to increased levels of MMP-12 in COPD airways. For example, microbial colonization and impaired phagocytosis can lead to persistent activation of immune cells and production of pro-inflammatory cytokines. These cytokines, in particular TNF-α and IL-1β [11, 12], stimulate macrophages and other cell types to produce MMP-12. Interestingly, based on the same study population as the current, we have previously reported that NK cells, capable of producing both TNF-α and IL-1β, were increased in COPD subjects [13]. Dysfunctional mitochondria are another well-described feature of COPD [14] and one that can further contribute to increased airway MMP-12. Leaky mitochondrial membranes generate reactive oxygen species that activate the NF-κB pathway, contributing to increased expression of MMP-12 [15].
In addition to these effects of chronic disease processes, in current smokers with COPD, the secretion of MMP-12 may also be influenced by acute effects of smoking. For example, tobacco smoke has been shown to cause leakage of serum containing plasminogen and prothrombin into the alveoli. After being activated, these proteins can bind to alveolar macrophage PAR-1 receptors [16], which leads to secretion and activation of MMP-12. Other mechanisms include upregulation of granulocyte–macrophage colony-stimulating factor (GM-CSF) and tachykinins [1], e.g., substance P and neurokinin, both of which have been suggested to increase secretion of macrophage MMP-12. In our study, smoking intensity was however not a significant predictor of MMP-12 concentrations.
The chicken or the egg?
The above-described mechanisms relate to MMP-12 in manifest COPD. However, increasing evidence of early-life events and genetics as major factors for the development of COPD raises the question of what comes first: manifest COPD or increased activity of MMP-12 in the airways? (Or even a causal relationship in both directions?).
It is widely recognized that in utero and early life tobacco smoke exposure leads to long-term adverse effects on lung function and increased risk for COPD. In mice models, in utero secondhand smoke exposure causes upregulation of MMP-12 at gene and protein levels and leads to significantly enlarged airspaces in the offspring [17]. MMPs are vital for normal lung development in mice as well as humans, with MMP-12 being expressed in the later stages of development [18]. During childhood, tobacco smoke exposure, lower respiratory tract infections, and asthma are risk factors for the later development of COPD [19]. In a longitudinal study from Tasmania, 75% of individuals who developed COPD in their 50 s had exhibited impaired lung function already during childhood [20].
As for genetics, it has been found in several cohorts that the minor allele of an SNP in the promoter of the MMP-12 gene (substitution A-to-G at position − 82; rs2276109) is associated both with a reduced risk of developing COPD and with a beneficial effect on lung function in children with asthma [21]. This functional polymorphism impairs AP-1 transcription factor binding, resulting in a lower expression of MMP-12. In another study, haplotypes of two SNPs in MMP-12 (rs652438 and rs2276109) were associated with severe to very severe COPD [22].
Based on our findings, the potential mechanisms described above, as well as the large body of evidence from previous studies supporting a major role for MMP-12 in COPD pathogenesis, we decided to investigate this matrix metalloprotease further in a pEWAS.
Association between MMP-12 and DNA methylation
To the best of our knowledge, this was the first time the role of DNAm in airway MMP-12 regulation was explored. Our pEWAS identified 34 loci where levels of DNAm in BAL cells were significantly associated with concentrations of MMP-12 in BAL. This model specifically estimated the relationship in a non-COPD context, highlighting shared regulatory mechanisms of MMP-12 across healthy and diseased states. Among genes annotated to the 10 pQTMs with the lowest p values, two are well-known to be associated with MMP-12 and ECM remodelling: TGFBR2 (Transforming Growth Factor Beta Receptor 2) and THBS4 (Thrombospondin 4).
TGFBR2 encodes a transmembrane protein acting as a receptor for the TGF-β signalling pathway, which can stimulate the expression of ECM proteins, the activity of MMPs in general, and MMP-12 in particular [11]. TGFBR2 is also implicated in interactions with other molecules associated with ECM remodelling, such as integrins, focal adhesion kinases, and thrombospondins.
THBS4 (also known as TSP-4) is an ECM protein and a member of the thrombospondin family whose expression is closely associated with TGF-β signalling [23]. Thrombospondins function to guide tissue remodelling and extracellular matrix synthesis through interactions with other ECM proteins and MMPs. THBS4 has been associated with interstitial lung disease [24], a group of conditions in which MMP-12 is often increased and plays a pro-fibrotic role [25].
Among the remaining top ten pQTM genes, four have previously been found to interact with MMP-12 or other MMPs in different settings (HOXD1, DIAPH3, TMEM17, and SORCS2); three to be involved in ECM interactions (ERO1A, GOLIM4, and TMEM17); while two have no previously known relationship with MMPs or ECM remodelling (LZTS1 Antisense RNA 1 and TRMT13). The biological importance of ECM-related gene regulation is underscored by the fact that six of our top ten pQTM genes are associated with either lung cancer or interstitial lung disease.
The effect of COPD on the MMP-12—DNA methylation relationship
Next, we sought to examine to what extent changes in DNAm might explain the increased MMP-12 in COPD airways. Here, we employed an interaction model in which COPD was included as a potential modifier. This analysis identified 66 interaction pQTMs, i.e., sites where the relationship between DNAm and MMP-12 levels was different in COPD compared to controls (e.g., slope of, variability around, or intercept of the regression line; see Figs. 4 and S1 for examples). For some interaction pQTMs, both mean methylation and MMP-12 concentrations were elevated in COPD subjects compared to controls. However, even when mean methylation was similar across groups, concentrations of MMP-12 were generally higher in those with COPD. This suggests that both DNAm and other mechanisms may contribute to the upregulation of MMP-12 in COPD airways.
Among the top 10 interaction pQTMs, eight were annotated to protein-coding genes, and all of those have previously been associated with either COPD or smoking (PRKAG2, ACSF3, IL6ST, CREBBP, KIF1A, SLURP1, CACNA2D2, and INSC), indicating biologically plausible results, and also a potential role for these genes in the dysregulation of MMP-12 in COPD. The remaining two interaction pQTM genes were long non-coding RNAs of which little or no mention can be found in the literature (LINC00378 and LOC643339).
PRKAG2 (Protein kinase AMP-activated non-catalytic subunit gamma 2) encodes for a sub-unit of the AMP-activated protein kinase (AMPK) enzyme, a central regulator of cellular energy metabolism implicated in COPD in several ways (e.g., compensation for mitochondrial dysfunction, regulation of oxidative stress and smooth muscle contraction). AMPK can be deactivated by MMP-12 proteolytic activity, leading to an increase of TNF-α [26]. Conversely, activation of AMPK seems to suppress the expression of at least some MMPs, although it is not clear if this applies to MMP-12 [27]. At the signalling level, this crosstalk between MMP and AMPK pathways might modulate inflammatory responses in the lung and has been suggested to play a role in emphysema development [28]. Based on machine learning analyses of gene expression, PRKAG2 has been proposed as a potential therapeutic target in COPD [29], it has also been found to be differentially methylated in several previous studies of the disease [30, 31].
ACSF3 (Acyl-CoA synthetase family member 3) is part of the acyl-CoA synthetase family, involved in fatty acid metabolism, and located exclusively in the mitochondrial matrix in humans [32], although its role in mitochondrial dysfunction in COPD specifically has not been examined. In an epigenetic study exploring the developmental origins of COPD, ACSF3 was found to be differentially methylated in smoke-exposed foetal lung samples compared to unexposed samples [33]. It has also been identified as a key regulator of COPD in an integrated analysis of lung tissue DNAm and gene expression [34].
Causal direction
Based on the currently available evidence, the causal direction of the above-described relationships is not fully elucidated, i.e., whether MMP-12 regulates DNAm or the other way around. For at least some MMPs, there is experimental support for the latter hypothesis; inhibition of DNAm has been shown to reduce methylation in MMP gene promoter regions, and this in turn to result in increased transcription and secretion of MMPs [35]. However, the alternative hypothesis, that airway MMP-12 could affect methylation of BAL cell DNA, is also biologically plausible. MMPs can alter cytokine activity in several ways—e.g., via interaction with neutrophils, activation of pro-cytokines through proteolytic cleavage, and ECM remodelling leading to the release of stored cytokines and activation of pro-inflammatory signalling pathways. Experimental evidence indicates that some of these cytokines (IL-1β and TNF-α in particular) can affect the activity of DNA methyltransferases and thus potentially DNAm [36, 37].
Integration with GWAS
In contrast to the order of nucleobases in the DNA, methylation marks are dynamic—they accumulate over time and can change due to environmental cues. However, at some loci, DNAm is strongly associated with genetic variants, i.e., sites where DNA sequence somehow shapes methylation patterns. This phenomenon has been reported for genetic variants (SNPs) associated with many diseases, including COPD [38].
As genetic determinants are important for COPD pathogenesis, we tested whether the identified epigenetic variation related to both MMP-12 and COPD was colocalized with genetic variants linked to the disease. To do this, we integrated our interaction pQTM results with those from a recent large-scale GWAS of 35,735 cases and 222,076 control subjects [10]. We found that merely 1/66 interaction pQTMs were colocalized with COPD-associated SNPs. This may suggest that factors influencing MMP-12 overexpression in COPD airways are more strongly related to environmental influences and disease processes than to genetic predispositions. However, genetic effects on MMP-12 expression could still manifest through mechanisms not captured by colocalization analysis, such as transcriptional regulation, post-transcriptional modifications, or epigenetic changes in other cell types or tissues.
Potential mechanisms
The classic mechanism by which DNAm alters gene expression involves promoter methylation, which inhibits the binding of transcription factors and subsequently leads to decreased expression of the target gene. It might thus seem unexpected that MMP-12 was not in the list of interaction pQTM genes. However, the process from DNA sequence to protein expression is in reality more complex, and there are several potential mechanisms that could explain the effect of COPD and DNAm on airway MMP-12 levels and activity. For example, DNAm could affect (through cis- or trans-effects) the availability and expression of (i) proteinases involved in the extracellular cleavage of MMP-12 prodomains—since MMPs are most often secreted as inactive proenzymes [7]; (ii) transcription factors and/or endogenous small RNAs that affect MMP-12 expression [39]; and/or (iii) enzymes involved in the post-translational machinery, since MMP-12 secretion and activity may be altered by for example phosphorylation [40]. As discussed above, the causal relationship between DNAm and MMP-12 might also be bidirectional, i.e., some interaction pQTMs could represent sites where methylation is affected by MMP-12 and/or other COPD processes and not the other way around.
Strengths and limitations
Currently, no other dataset that contains measurements of MMPs and DNAm levels in BAL has been published. Consequently, our findings could not undergo validation in a suitable external dataset. While MMP/DNAm datasets based on peripheral blood exist, the marked differences in DNAm patterns and MMP-12 concentrations between compartments make replication in such a dataset inherently challenging to interpret. So, although our analyses indicate that we did capture a relevant signal (e.g., eight out of the top ten pQTM genes were previously associated with MMPs/ECM remodelling, and all top ten protein-coding interaction pQTM genes with COPD/smoking), availability of an external dataset enabling validation would have further reinforced our findings.
Another constraint is the limited number of participants included. While this limitation is common in studies relying on bronchoscopies, a larger study cohort would have been desirable. Furthermore, the ELISA kit used to measure proteins in BAL could not discriminate between active and inactive forms of MMPs.
The absence of genotype data in our cohort limits our ability to directly evaluate cis-meQTLs or explore SNP-methylation-protein interactions within the study population. For example, we were unable to test the potential impact of rs2276109, a SNP shown to affect both COPD risk and MMP-12 expression [21], on DNAm or protein levels. Integration with GWAS meta-analysis data provided an alternative approach to investigating the relationship between genetic and epigenetic variation, enabling us to contextualize our findings within a larger, well-characterized dataset.
One major strength is the well-characterized study population. Statistical adjustments are a powerful tool, but their effectiveness is entirely dependent on the quality of the control variable data. In the current study, all subjects were recruited from the longitudinal OLIN COPD study which provided detailed and reliable information on smoking habits, use of ICS, and other variables included as covariates in the models. Another strength is the use of BAL and not surrogate tissue such as peripheral blood. When trying to understand processes that take place in the lungs (e.g., expression and secretion of airway MMPs, in health and disease), examining target organ tissue is more likely to produce more relevant information than surrogates. Furthermore, integrating several layers of omics data allows for a more complete understanding of disease mechanisms and may provide more robust results, especially when sample size is limited [41].



留言 (0)