POI refers to the cessation of ovarian function before the age of 40. According to a recently published meta-analysis, the global prevalence of this disease is estimated at 3.7% [22]. Currently, treating POI with recognized and reliable methods remains challenging. In this field, Chinese herbal medicines, such as HSYLF, are gaining increasing popularity.
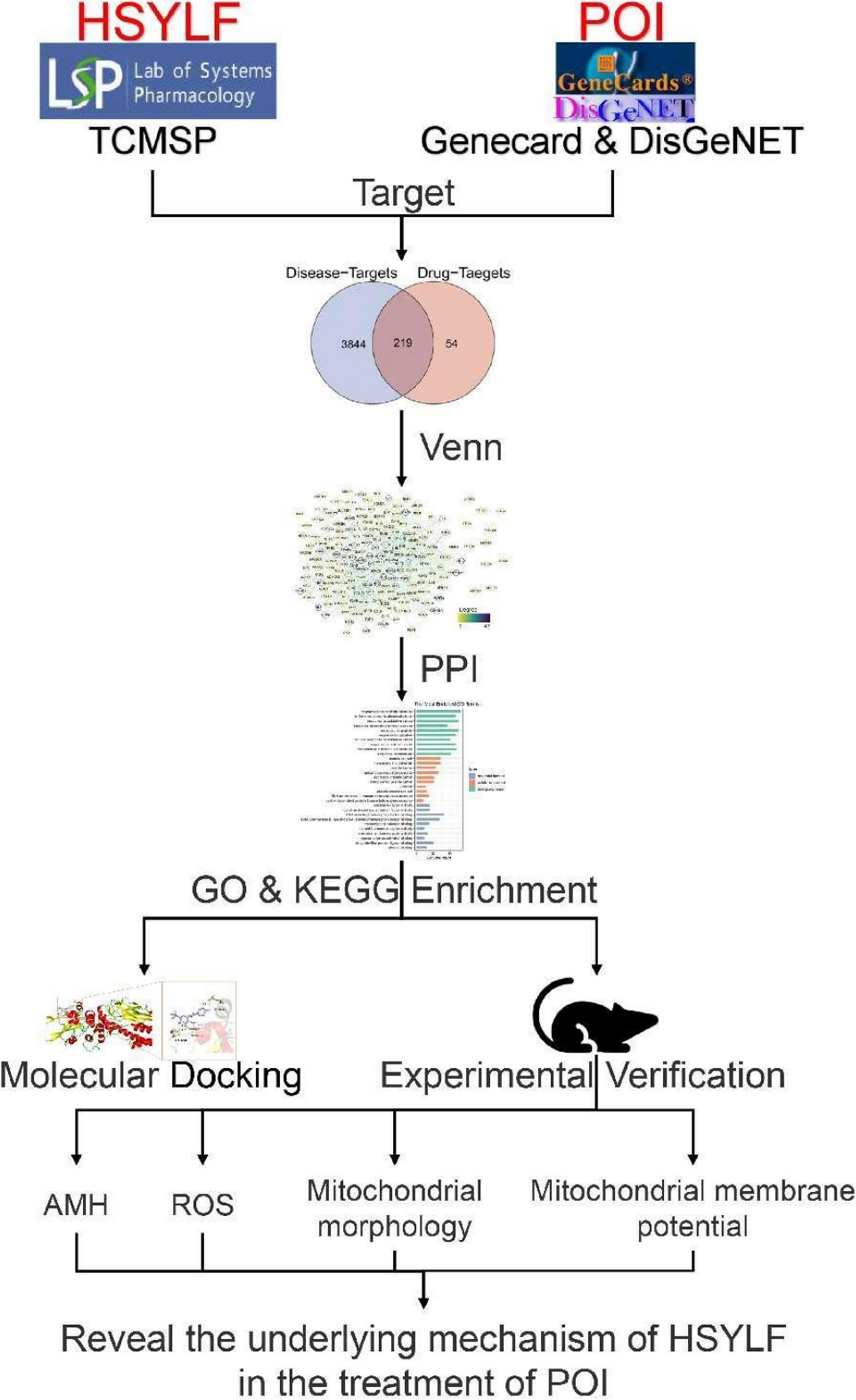
The mechanism of Yulin Formula in treating cyclophosphamide-induced diminished ovarian reserve has been reported. This mechanism highlights that YLF significantly reduces ovarian cell apoptosis and improves ovarian function by regulating the phosphatidylinositol 3-kinase/AKT signaling pathway, leading to an increase in AMH levels close to normal. Such a result provides theoretical support for the application of YLF in the field of reproductive health [23]. In this study, special attention is given to investigating the effects of HSYLF on POI. To better understand the mechanisms of HSYLF in treating POI, mice were used as animal models in pharmacodynamic experiments, and network pharmacology and molecular docking techniques were employed to investigate the chemical components and targets of HSYLF. Through network pharmacology analysis, 204 active components and 219 key genes identified in HSYLF were associated with POI. Herbs in HSYLF with higher active components included formononetin, naringenin, licochalcone A, and quercetin. Among them, formononetin, a phytoestrogen, can bind to estrogen receptors due to its structural similarity to estrogen [24]. As an estrogen-like compound, formononetin can enhance estrogen-like functions by increasing the expression of estrogen receptor proteins [25]. Additionally, formononetin can reduce the ROS levels by activating the phosphatidylinositol 3-kinase/AKT and nuclear factor erythroid 2-related factor 2 signal pathways and increasing the antioxidant gene expression [26]. Naringenin exhibits good corrosion inhibition performance and acts as a chemical group and a scavenger (superoxide dismutase). Additionally, it functions as a free radical scavenger and inhibits the activities of oxygen promoting enzymes, such as nicotinamide, adenine dinucleotide phosphate oxidase, cyclooxygenase, lipoxygenase, xanthine oxidase, and metal ion chelation [27]. Consequently, naringenin can inhibit inflammatory responses, protect mitochondrial function, and inhibit apoptosis [28,29,30,31]. Moreover, due to the antioxidant properties of quercetin, it can reduce DNA damage, mitochondrial damage, endoplasmic reticulum damage, and cellular damage caused by reactive oxygen free radicals [32]. The molecular docking results indicated that these active components have potential binding affinities with therapeutic targets for POI, indicating a moderate potential for binding. Hydrogen bonding interactions were observed between residues Asn-647, Lys-658, and Gly-656 with licochalcone A molecules; residue DG-308 with formononetin molecules; residues ASP-184, LYS-131, MET-125, and ASP-123 with naringenin molecules; residue ASN-106 with 1,6-dihydroxy-5-methoxy-2-(methoxymethyl)anthracene-9,10-dione molecules; and residues GLU-326, GLU-349, and ASN-345 residues with quercetin molecules. The shorter hydrogen bonding distances imply stronger binding affinities. Additionally, the more stable binding conformation was associated with the lower required binding energy. Binding energies of ≤ − 4.25 kcal/mol, ≤ − 5.0 kcal/mol, and ≤ − 7.0 kcal/mol indicate weak, good, and strong binding activities, respectively. The molecular docking analysis in this study identified JunB, c-Jun, STAT3, and MAPK3/1 as target proteins due to their critical roles in the pathophysiology of POI. JunB and c-Jun could regulate autophagy, with their expression responding to autophagy-inducing signals. Autophagy is essential for maintaining mitochondrial health and removing damaged mitochondria, contributing to alleviating POI [33]. STAT3 is involved in autophagy through various pathways, exerting a protective effect on the ovaries tissue, including both oocytes and their surrounding cumulus cells within cumulus-oocyte complexes (COCs) [34]. The MAPK3/1 pathway is associated with growth hormone-mediated reduction of aneuploidy in aged oocytes, thereby improving the quality of denuded oocytes (DOs) following cumulus cell removal [35]. These target proteins were selected to better understand their mechanisms in POI and to explore their potential as therapeutic targets at the molecular level. Overall, these data suggest that 1,6-dihydroxy-5-methoxy-2-(methoxymethyl)anthracene-9,10-dione molecules, formononetin and naringenin may be the key active components of HSYLF in the treatment of POI.
PPI network analysis showed that the hub genes of HSYLF in treating POI included heat shock protein 90 alpha family class A member 1, Jun, MAPK3, STAT3, and TP53. MAPK3, a member of the MAPK family, regulates a variety of cellular processes, including proliferation, differentiation, and cell cycle progression, in response to various extracellular signals. The activity of MAPK3 during oocyte maturation is crucial for granulosa cell function [36]. STAT3, a member of the STAT family, plays an essential role in processes such as cell proliferation, differentiation, and apoptosis. STAT3 has been reported to be capable of entering mitochondria, where it regulates the activity of electron transport chain complex enzymes, thereby affecting mitochondrial function [37]. Additionally, TP53 is closely related to ovarian granulosa cell apoptosis and proliferation [38]. GO and KEGG enrichment analyses indicated that the anti-POI targets of HSYLF mainly regulated response to xenobiotic stimulus, cellular response to chemical stress, and response to oxidative stress, with the main manifestation observed in responses to oxidative stress and response to ROS.
Highly correlated signaling pathways included the endocrine resistance, TNF signaling pathway, interleukin (IL)-17 signaling pathway, chemical carcinogenesis—receptor activation, and AGE-RAGE signaling pathway in diabetic complications. Therefore, HSYLF might play an important role in treating POI by regulating these signaling pathways.
TNF-α activates the Toll-like receptor 4 signaling pathway, further triggering the nuclear factor-κB (NF-κB) signaling pathway. Such a process promotes the phosphorylation and nuclear translocation of NF-κB, leading to an increased expression of pro-inflammatory factors, such as IL-6 and TNF-α in ovarian granulosa cells. During ovulation, TNF-α exhibits dual functions. On the one hand, TNF-α promotes the growth of preantral granulosa cells, inhibits differentiation, and enhances vascularization around the follicle, thus facilitating follicular rupture and ovulation. On the other hand, TNF-α can promote the apoptosis of luteinizing granulosa cells after ovulation, inhibit luteogenesis, and facilitate luteolysis. These results indicated that the mechanism of TNF-α is complex. Additionally, TNF-α can induce inflammatory responses, promote cell proliferation and survival, and trigger cell differentiation and apoptosis. The activation of TNF-α also triggers the phosphorylation and degradation of IκB proteins, releasing NF-κB (p65/p50 complex) for translocation to the nucleus. In the nucleus, NF-κB binds to specific DNA sequences, regulating the expression of genes associated with cell proliferation, survival, and apoptosis [39,40,41]. This regulatory mechanism may lead to increased proliferation of ovarian granulosa cells and potential induction of apoptosis, thereby significantly impacting ovarian physiological function.
IL-17 activates MAPK signaling pathways, including extracellular signal-regulated kinase, c-Jun N-terminal kinase, and p38 MAPK, which are crucial for cell proliferation, differentiation, and survival. By increasing the phosphorylation of proteins in the MAPK pathway, IL-17 promotes the growth of ovarian granulosa cells. Additionally, IL-17 can enhance the expression of pro-inflammatory factors, regulate the ovarian environment, and promote angiogenic factors such as vascular endothelial growth factor, increasing follicular vascularization and ensuring adequate nutrient and oxygen supply. Therefore, IL-17 facilitates the growth and vascularization of granulosa cells, influencing follicular development and ovarian function [42].
Oxidative and carbonyl stress can be mediated by the AGE-RAGE signaling pathway, and the accumulation of AGEs leads to accelerated ovarian aging [43]. AGEs binding to RAGE can initiate a cascade of intracellular signaling, activating various inflammatory pathways, including NF-κB. This activation promotes the expression of pro-inflammatory cytokines, resulting in a chronic inflammatory state in the ovarian microenvironment. Under the condition of ovarian aging, the accumulation of AGEs exacerbates oxidative stress, leading to mitochondrial dysfunction and elevated levels of ROS. Such results can cause cellular damage, including ovarian cell apoptosis, thereby reducing ovarian reserve and impacting overall reproductive function [44].
Additionally, AGEs accelerate the activation of primordial follicles through the AGE-RAGE pathway; however, premature activation can result in the early depletion of the primordial follicle pool. Moreover, AGEs can also promote the proliferation of ovarian granulosa cells and oocytes, accelerating the activation of primordial follicles in the ovary, which may further accelerate ovarian aging and lead to ROS [45].
HSYLF is speculated to significantly improve ovarian function in patients with POI, considering that mitochondria are the site of ROS production and the primary target of oxidative damage. It is inferred that HSYLF may improve hormone levels and oxidative stress response by modulating its fluorescence intensity, distribution, morphology, quantity, and activity levels.
Pan et al. discovered [46] that massive accumulations of free radicals and ROS can exert detrimental effects on cells, such as increased mitochondrial membrane permeability, destruction of cristae structure, and reduced numbers of mitochondria debris. [47, 48], contributing to mitochondrial dysfunction [49]. Excessive ROS generation can destroy the ovarian follicular granulosa cells, luteinize follicles, and induce granulosa cell apoptosis [50]. When granulosa cells undergo apoptosis, oocytes lose the signals and energy required for physiological activities, resulting in halted development. This can lead to follicular atresia or even failure, further causing the decrease in ovarian reserve function. Moreover, this decrease severely impacts the dynamic characteristics of mitochondrial biogenesis of surrounding granulosa cells, forming a vicious cycle, and ultimately promoting the progress of POI [51,52,53].
Through the animal experiments, HSYLF might improve ovarian function in POI mice by decreasing mitochondrial ROS, increasing membrane potential, and improving mitochondrial function. These experimental results confirmed that the AMH levels in the model blank group of mice were significantly reduced, while the levels in the high-dose HSYLF group were markedly higher compared to the model blank group and approached those of the normal control group, indicating that HSYLF could improve ovarian function. Intragastric administration of HSYLF reduced ROS levels, with greater benefits observed in the TCM-treated group relative to the Western medicine group. Microscopy and fluorescence staining images of mitochondria revealed stronger fluorescence in the high-dose HSYLF group, with improved distribution, morphology, and quantity of mitochondria. According to the mitochondrial fluorescence staining images and electron microscopy, the number and morphology of mitochondria were significantly improved in the HSYLF treatment groups, indicating restored mitochondrial function. The membrane potential was notably increased, suggesting a significant effect of HSYLF on mitochondrial activity. The presence of HSYLF could modify hormone levels and oxidative stress responses in POI mice by altering fluorescence intensity, distribution, morphology, quantity, and activity level of mitochondria in oocytes, thereby improving oocyte quality and ovarian function.
In this study, we explored the methods for selecting disease targets using the DisGeNET and GeneCards databases and acknowledged the significant advantages of the Gene Expression Omnibus dataset in terms of accuracy for gene expression analysis. The DisGeNET and GeneCards databases were the initial choice due to their extensive coverage. However, in future research, the use of the Gene Expression Omnibus dataset will be prioritized to enhance data accuracy and improve the reliability of target selection. This study primarily focused on the mitochondrial regulatory effects of HSYLF on ovarian function, and thus expression validation for specific targets, such as STAT3, Jun, and TP53, was not conducted. Nevertheless, these targets might play critical roles in the pathogenesis of POI. Future research will involve more in-depth validation of these target expressions to reveal the potential multi-target regulatory mechanisms of HSYLF, providing a more comprehensive scientific basis for its mechanisms of action. Furthermore, based on the current animal experiment results, the potential of HSYLF in treating POI is recognized. However, animal experiment results may not fully reflect the physiological and pathological conditions in humans. Therefore, future research should include systematic clinical trials in a clinical setting to validate the safety and efficacy of HSYLF in humans. These trials will contribute comprehensively to assessing the therapeutic potential of HSYLF, further elucidating its regulatory effects on mitochondrial function and oxidative stress responses across multiple signaling pathways. This study provides definite evidence for the molecular mechanisms of HSYLF in treating POI by decreasing mitochondrial ROS, increasing membrane potential, and improving mitochondrial function. The treatment of POI with HSYLF may affect the cellular response to oxidative stress and ROS, thereby affecting the oocyte mitochondrial function and maintaining the normal function of ovarian granulosa cells. Findings on active components and related key targets of HSYLF also confirmed its characteristics of multi-component, multi-target, multi-pathway, and holistic regulatory effects against POI. The study establishes a theoretical basis for further research on the treatment of POI with HSYLF. These results need to be generalized with further mechanistic studies, and the deep clinical value of HSYLF should be investigated in the future.




留言 (0)