
記住我




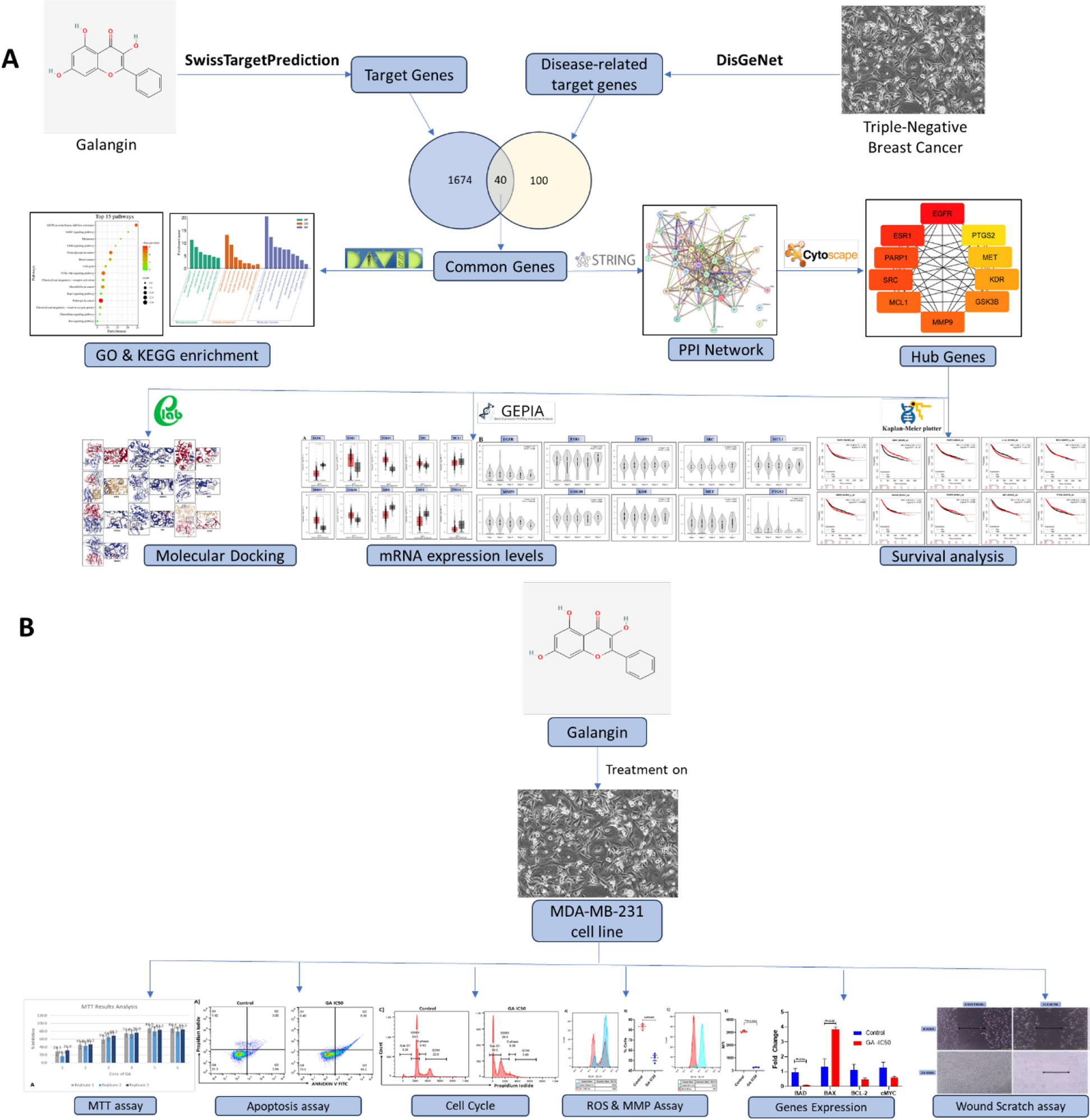


It follows from our findings that Galangin follows Lipinski's Rule of Five (RO5). Adhering to Lipinski's Rule of Five, a molecule with medicinal potential must have the following characteristics: a polar surface area (PSA) of 140 Ų or smaller, a molecular weight (MW) below 500 g/mol, and a computed octanol/water partition coefficient (XLogP3) below 5. For the best drug-ligand interactions, these three qualities are crucial. In addition, Galangin has a maximum of 10 hydrogen bond acceptors and no more than 5 hydrogen bond donors, with less than 10 rotatable bonds. Galangin possesses beneficial drug-like properties since its chemical characteristics meet the RO5 requirements.
Target identification and analysisThis study used the DisGeNet database to find targets related to triple-negative breast cancer. The search keyword used was "Triple-negative breast cancer." There were 1,674 targets associated with triple-negative breast cancer that were found during the screening. One hundred Galangin targets were found by searching the Swiss Target Prediction database. Forty genes were found to be shared between the Galangin targets and the triple-negative breast cancer targets (Fig. 2).
Fig. 2
Number of genes common between triple-negative breast cancer and Galangin
The establishment of a PPI network and the identification of top hub genesTo examine the interplay between the designated targets' proteins, the study made use of the STRING Database. Clusterone in Cytoscape 3.10.1 was used to conduct further analysis on the PPI network in order to identify modules. The network, which includes 40 shared genes, showed a p-value of less than 1.0e-16, indicating that the proteins in this network interact more strongly with each other than a randomly selected set of proteins from the genome with similar size and degree distribution. The proteins are shown to be biologically connected as a cohesive unit in Fig. 3, which is supported by this enrichment. Three distinct clusters were identified in the investigation. The first cluster exhibited a p-value of less than 0.05 (p-value – 1.0e-16), the second cluster showed a p-value of 6.46e-11, and the third cluster noted a p-value of 0.000233. The p-values indicate that the target proteins within each cluster are significantly connected and likely share similar biological functions. Cluster 1 (Fig. 3) was associated with the positive control of cellular proliferation, apoptosis, cell migration, and the MAPK signaling cascade. In contrast, Cluster 2 (Fig. 3) was linked to cell division, the positive control of the G2/M transition in the mitotic cell cycle, and the negative regulation of the G1/S transition in the mitotic cell cycle.
Fig. 3
Protein–protein interaction network comprising 40 genes: cluster 1 has 26 proteins (nodes) interconnected by 155 edges, with a PPI enrichment p-value of 1.0e − 16. Cluster 2 has 8 proteins (nodes) interconnected by 24 edges, with a PPI enrichment p-value of 6.46e − 11. Cluster 3 has 4 proteins (nodes) and their interconnections (four edges), (PPI enrichment p-value: 0.000233)
Using a variety of techniques, the study determined that EGFR, ESR1, PARP1, SRC, MCL1, MMP9, GSK3B, KDR, MET, and PTGS2 were the ten main hub genes. The most highly active gene among these hub genes was found to be EGFR. The study identified ten key hub genes: EGFR, ESR1, PARP1, SRC, MCL1, MMP9, GSK3B, KDR, MET, and PTGS2. It used a variety of methods to reach this conclusion. The EGFR gene was determined to be the most active of these hub genes (Table 1, Fig. 4).
Table 1 Various algorithms used to determine the top 10 hub genes based on CytoHubbaFig. 4
Figure showing the top 10 hub genes from common 40 genes
Functional enrichment analysisApproximately twenty-five GO terms were discovered in an analysis of the forty genes that were found to be common. Angiogenesis, biological rhythms, mitosis, and lipid transport are all biological processes (BP) that these targets are involved in, according to the research. The kinetochore, centromere, chromosome, and cytoskeleton were all part of the cellular component (CC) findings. When it comes to molecular function (MF), the targets primarily participate in activities such as tyrosine-protein kinase, translocase, viral entry mediated by host cell receptors, and serine/threonine-protein kinase (Fig. 5).
Fig. 5
GO enrichment analysis was conducted by selecting 7 biological processes (BP), 8 molecular functions (MF), and 10 cellular components (CC) from the DAVID Database for the targets associated with Galangin in the treatment of breast cancer
KEGG enrichment analysisThe KEGG pathway analysis yielded 75 pathways in total. Further analysis was limited to the top fifteen pathways. Pathways associated to cancer, microRNAs in cancer, proteoglycans in cancer, resistance to EGFR tyrosine kinase inhibitors, and the PI3K-Akt signaling pathway were among the many that the ten main hub genes showed substantial associations with, according to the analysis (Fig. 6).
Fig. 6
A Bubble plot illustrating the enrichment of the top 15 signaling pathways related to oral cancer was created. The X-axis denotes the enrichment factor of the genes, and the Y-axis signifies the various pathways. The circles in the graphic are color-coded and sized according to the Log10 (p-value), with red representing pathways containing the highest gene counts and light green denoting pathways with fewer genes
The genes EGFR, PIK3R1, MET, GSK3B, SRC, and CDK6 were consistently found to be involved in the 10 major pathways. These genes showed significant associations with the PI3K-Akt signaling system, which is important in regulating a variety of functions in cells that influence carcinogenesis, such as metabolic pathways, cell survival, cell proliferation, gene expression, and protein synthesis [33]. Table 2 shows that Galangin may play a role in breast cancer treatment, with the PI3K-Akt signaling pathway as a key focus and apoptosis as the predominant cellular mechanism.
Table 2 Top 10 pathways showing the genes involvedPhosphoinositide 3-kinase-Akt signaling pathway PIK3CA is a constituent of the PI3K signaling pathway, often dysregulated in numerous cancer types, and is implicated in cell growth, proliferation, differentiation, protein synthesis, and apoptosis [34]. Somatic mutations in PIK3CA are deemed responsible for the enhancement of its kinase activity, leading to cellular transformation [35]. Phosphoinositide 3-kinases (PI3Ks) are enzymes crucial for cellular development and differentiation. This is initiated by receptor tyrosine kinases (RTKs) such as epithelial growth factor receptor (EGFR). Protein kinase B (PKB or AKT) is phosphorylated and activated subsequent to the activation of PI3K, followed by its translocation to the cell membrane [36]. AKT can activate the mammalian Target of Rapamycin (mTOR) complexes (mTORC1 and mTORC2), leading to the transcription of PI3K pathway signaling molecules, such as serine/threonine protein kinase SGK146. This consequently results in enhanced protein synthesis and cellular proliferation [37]. The PI3K/AKT pathway is enhanced by IGF-1, EGF, and sonic hedgehog (SHH) [38], and is suppressed by several factors, including phosphatase and tensin homolog (PTEN). Alterations and mutations in the PTEN/PI3K/AKT/mTOR pathway enhance cellular proliferation and diminish apoptosis [39]. These mutations have been associated with drug resistance in the management of oral cancer. Drug resistance is believed to result from modified activation of the MDR-1 gene and extended cellular viability. The predominant mutational sites in oral cancer are located in exons 9 and 20 of the PIK3CA gene. E542K, E545K, H1047R, H1047Y, and H1048Q are prevalent variants in this gene [40].
Hub target confirmation using molecular dockingTo assess the reliability of the drug-target interactions, ten hub genes were selected as targets for molecular docking studies. The structure of Galangin was submitted to CB-DOCK to evaluate its docking affinity with EGFR, ESR1, PARP1, SRC, MCL1, MMP9, GSK3B, KDR, MET, and PTGS2. A reduced energy value signifies a more stable conformation of the ligand binding to the receptor, implying an increased probability of contact. This investigation revealed that the binding energies between Galangin and the core target proteins were below − 5.0, signifying a robust binding affinity between Galangin and the core targets. The binding energies are detailed in Table 3, while the docking schematic representations of the interactions between the target proteins and Galangin are shown in Fig. 7.
Table 3 Molecular docking scores of Galangin and hub target proteinsFig. 7
Sketch and matching diagrams of molecular docking of Galangin and top 10 hub genes (target proteins)
External validation of hub genesmRNA expression levels of hub genesWe employed the GEPIA database to analyze the differential expression of hub genes in breast cancer and normal tissues. Our study demonstrated that mRNA levels of ESR1, PARP1, GSK3B, and MMP9 were considerably increased in breast cancer tissues relative to normal samples (p < 0.01) (Fig. 8A). Additionally, we examined the correlation between mRNA levels of the hub genes and the pathological stages of breast cancer. The findings demonstrated that the expression levels of EGFR, MMP9, ESR1, MET, and PTGS2 showed considerable variations throughout various clinical phases. Notably, EGFR, MMP9, ESR1, and PTGS2 exhibited significant elevations in stage II (Fig. 8B). The results indicate that the expression levels of these five genes may correlate with the advancement of breast cancer.
Fig. 8
The mRNA expression levels of hub genes in The Cancer Genome Atlas (TCGA) and Genotype-Tissue Expression (GTEx) datasets. A mRNA expression levels in the GEPIA database (Boxplot of hub genes). Red represents breast cancer tissue, whereas grey signifies normal breast tissue. B mRNA expression levels and disease stages in the GEPIA database. (Stage plot of hub genes)
Survival analysis of the hub genesA survival analysis was carried out on the ten hub genes, which are as follows: EGFR, ESR1, PARP1, SRC, MCL1, MMP9, GSK3B, KDR, MET, and PTGS2. Four thousand nine hundred and twenty-nine breast cancer patients from the TCGA database were used in the investigation. The results of the study demonstrated that every hub gene exhibited a significant association with a poor prognosis or outcome (p < 0.05, Fig. 9).
Fig. 9
Kaplan–Meier overall survival analyses of patients with breast cancer based on the expression of the ten hub genes. HR, hazard ratio ("http://kmplot.com/analysis/index.php?p=service&cancer=pancancer_rnaseq")
Anticancer activity of Galangin on MDA-MB231 cell lineCytotoxic effect of Galangin on MDA-MB231 cellsVarious dosages of Galangin (5, 10, 20, 40, 60, and 80 µg/ml) and Carboplatin (10, 20, 40, and 80 µg/ml) exhibited a significant cytotoxic effect on the MDAMB231 cell line in a dose-dependent manner, with an IC50 value of 17.7 ± 5.1 µg/ml. Carboplatin had an IC50 value of 31.6 ± 8.1 µg/ml. This concentration of Galangin (IC50 17.7 ± 5.1) was utilized for all subsequent tests.
Apoptosis Regulation and the Role of GalanginA 24-hour apoptosis investigation was conducted on MDA-MB231 cells using Galangin at its IC50 concentration. Figure 10A shows dot plots that illustrate how Galangin induces cell death. Figure 10B shows that compared to control cells, there is a significant decrease in viable cells (73.95 ± 1.2%) and an increase in early apoptotic cells (17.75 ± 0.6%) and late apoptotic cells (7.72 ± 0.8%). Galangin mostly causes cell death by inducing early apoptosis, according to these studies.
Fig. 10
Effect of Galangin treatment on apoptosis and cell cycle in MDA-MB231cells. A Dot plot shows apoptosis ratios of MDA-MB231 cells in control (left) and after treatment of Galangin (right) for 24 h using propidium iodide and Annexin-V FITC. B Bar graph shows the percentages of live, early apoptotic, late apoptotic, and necrotic cells in control (blue) and after treatment ofGalangin (red) for 24 h. C Histogram represents different stages of the cell cycle in MDA MB231 cells in control (left) and after treatment of Galangin (right) for 24 h using propidium iodide. D Bar graph shows the percentages of cells in sub-G1, G0/G1, S, and G2/M phases in control (blue) and after treatment of Galangin (red) for 24 h. Error bar shows mean with standard deviation. *p < 0.05,**p < 0.01, ***p < 0.001
The impact of Galangin on the regulation of the cell cycle in MDA-MB231 cellsWe performed an additional examination into the effects of Galangin therapy on the cell cycle of MDA-MB231 cells. Figure 10C displays sample graphs depicting different phases of the cell cycle in control cells compared to those treated with Galangin at dosages corresponding to its IC50 for 24 hours. Treatment with Galangin resulted in a significant increase in the proportion of cells in the Sub G1 phase (58.05 ± 0.6%), accompanied by a marked decrease in cells in the G0/G1 phase (28.85 ± 0.4%) and the G2/M phase (3.63 ± 0.2%), while the S-phase cell count remained stable (8.36 ± 0.03%), compared to control cells (Sub G1: 2.8 ± 0.5%, G0/G1: 64.55 ± 2.0%, S-phase: 9.25 ± 0.2%, G2/M: 22 ± 1.1%) (Fig. 10D). This study highlights the specific impact of Galangin in inhibiting cell cycle progression during the Sub G1 phase, suggesting its potential significance for therapeutic strategies aimed at apoptosis.
Impact of Galangin on intracellular reactive oxygen species (ROS) levels and mitochondrial membrane potential (ΔΨm) in MDA MB231 cellsSubsequently, we assessed the effect of Galangin on intracellular reactive oxygen species levels and mitochondrial membrane potential in MDA-MB231 cells. Figure 11A presents sample graphs demonstrating intracellular ROS levels prior to and following Galangin therapy, whereas Fig. 11C displays corresponding graphs for mitochondrial membrane potential. Post-Galangin therapy, a notable reduction in ROS levels (52.53 ± 4.82%) was recorded in contrast to the control (83.10 ± 3.31%) (Fig. 11B). Furthermore, Galangin administration resulted in a significant decrease in mitochondrial membrane potential (MFI: 256 ± 20.66) relative to the control (MFI: 3071 ± 96.46) (Fig. 11D).
Fig. 11
Impact of Galangin administration on reactive oxygen species production and mitochondrial membrane potential in MDA-MB231 cells. A The histogram illustrates the levels of reactive oxygen species (ROS) production prior to (blue) and following the administration of Galangin (red) in MDA MB231 cells. B The scatter dot plot illustrates the quantitative assessment of ROS generation prior to (red) and subsequent to (blue) the treatment with Galangin in MDA MB231 cells. C The histogram illustrates the mitochondrial membrane potential (ΔΨm) in MDA MB231 cells before (blue) and after treatment with Galangin (red). D The scatter dot plot illustrates the quantitative assessment of mitochondrial membrane potential in MDA MB231 cells prior to (red) and following (blue) Galangin therapy. The error bar represents the mean accompanied by the standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001
Activity of Galangin on Wound Scratch AssayFollowing cell seeding, a 24-h incubation period was allowed prior to administering IC50 doses of Galangin. A control group was also established. The wells were evaluated at 0 and 24 h to determine the impact of the treatments. The results demonstrated that the control group displayed cell proliferation and migration after 24 h, which was absent in the Galangin-treated group (Fig. 12A). The Galangin-treated group exhibited a substantial reduction of cell proliferation and migration at 24 h in comparison to the control group (Fig. 12B).
Fig. 12
Illustrative Image for scratch assay. A MDAMB231 cells were treated with Galangin at IC50 (17.21 µg/ml) after 24 h. The cellular scratch was evaluated at 0 and 24 h utilizing a Magnus INVI microscope at × 100 magnification. The migration of MDAMB231 cells was inhibited by Galangin IC50 after 24 h, as illustrated in the second vertical panel. The initial panel depicted the control group. B The data were presented as mean ± standard deviation (SD), and “two-way ANOVA” was employed for analysis, followed by “Tukey” post hoc tests at ****p < 0.0001; ***p < 0.001; **p < 0.01 in comparison to the control at the corresponding time point
In MDA MB231 cells, galangin causes apoptosis by upregulating the expression of the BAX gene and downregulating that of the BAD geneHaving identified Galangin-induced apoptosis in MDA MB231 cells, we continued to examine its effects on the expression levels of critical apoptosis-related genes: BAD, BAX, BCL-2, and cMYC. Our findings indicated a substantial reduction in BAD gene expression coupled with a marked elevation in BAX gene expression subsequent to Galangin administration. Moreover, Galangin decreased the expression levels of BCL-2 and cMYC relative to the control cells (Fig. 13). These findings indicate that Galangin may enhance apoptosis primarily by upregulating the BAX gene while downregulating the BCL-2 gene, thereby enhancing mitochondrial permeabilization. The IC50 value demonstrated a significant decrease in c-cMYC expression. This discovery indicates that Galangin may impede c-cMYC expression by attenuating TGF-β, therefore modulating cellular proliferation and death (Fig. 13). The IC50 value of Galangin significantly influenced the aforementioned gene expression, contributing to its anti-cancer efficacy against TNBC primarily as an apoptotic promoter and regulator of cellular proliferation.
Fig. 13
Effect of Galangin administration on the expression of several apoptosis-related genes. The bar graph illustrates the fold change in gene expression levels of BAD, BAX, BCL-2, and cMYC, prior to (blue) and subsequent to (red) the administration of Galangin in MDA-MB231 cells. The error bar represents the mean accompanied by the standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001
留言 (0)