
記住我



Reactive oxygen species (ROS) are molecules derived from the reduction of molecular oxygen (O2). After oxidizing carbon in biomolecules, this process generates a range of oxidant species, including superoxide anion, hydroxyl radical, peroxyl radical, and peroxide [1]. In cancer cells, ROS play a crucial role in fundamental processes by interacting with different signaling pathways involved in cellular survival, angiogenesis, tumor progression, and metastasis [2,3,4,5]. ROS levels in cancer cells are higher compared to non-tumor cells, primarily due to their elevated metabolic rates. The main sources of ROS in cancer cells are mitochondria and the enzymatic activity of NADPH oxidase proteins (NOXs). The heightened presence of ROS contributes to the dynamic cellular environment in cancer, influencing key processes that drive malignancy [4]. However, the exact contributions of ROS from specific sources, such as mitochondria versus NOXs, in cancer progression remain poorly studied.
The NOX protein family is considered one of the most crucial sources of ROS production in eukaryotic cells. This family comprises seven members known as NOX1-5 and DUOX1-2, each participating in several cellular processes, including hormone production, modification of the extracellular matrix, host defense for immune cells, and redox signaling [6,7,8]. NOXs, membrane proteins with a conserved catalytic core of six transmembrane alpha helices, display structural similarity but vary in cellular distribution, activation mechanisms, and regulatory systems [9, 10]. Recent studies have underscored the significant importance of NOX-derived ROS in cancer cells, showcasing their crucial influence on cell survival, tumor development, and progression [11, 12]. While there is substantial evidence supporting the role of NOX-derived ROS in cancer, the molecular mechanisms linking NOX activity to tumor progression remain incompletely understood.
This involvement occurs via the activation of different signaling pathways related to the expression and secretion of matrix metalloproteinases (MMPs) [13, 14], the regulation of epithelial-mesenchymal transition, and the generation of invadopodia [15]. This literature review aims to compile relevant background information on the influence of the NOX protein family expression and function on the formation and activity of invadopodia, as well as their role in cell invasion.
Cell invasion and invadopodiaThe fundamental process enabling cancer cells to migrate or invade, involves the acquisition of mesenchymal characteristics, which occurs through epithelial-mesenchymal transition (EMT). EMT is characterized by the loss of epithelial features, such as cellular polarity and cell–cell junctions, leading to the acquisition of a mesenchymal phenotype associated with increased cell motility [16]. This transition involves the loss of molecular epithelial markers and the acquisition of mesenchymal markers such as N-cadherin, Vimentin, and fibronectin and the loss of E-cadherin [17]. Migration in cancer cells is mediated by the coordinated action of cytoskeletal components and proteins associated with the formation of protrusions, such as lamellipodia and filopodia [18]. The EMT process and the acquisition of increased cell motility are crucial for tumor cells to invade healthy tissues.
The cell invasion is the capacity through which tumor cells can penetrate and invade surrounding tissues until they reach the lymphatic and blood vessels, facilitating dissemination to more distant organs [19,20,21]. This process represents the initial step of metastasis and is mediated by various biochemical and physical factors in the tumor microenvironment, inducing a remodeling of the cytoskeleton and extracellular matrix (ECM) [22]. To initiate the invasion process, tumor cells must penetrate the ECM through the formation of specialized protruding structures known as invadopodia [23]. Invadopodia are composed of actin and surrounded by several adhesion proteins such as integrins, paxillins, and talins [24, 25]. The initiation of invadopodia formation involves the creation of precursor complexes enriched with actin-regulating proteins like cortactin, cofilin, WASP, and Arp2/3. These complexes enable cells to create protrusions by assembling actin filaments, which gradually become organized and anchored to the cell's surface, this, together with the recruitment of several proteins, ultimately forming functional structures [24, 26]. This activity is primarily mediated by metalloproteinases (MMPs), a family of zymogen proteases involved in the degradation and remodeling of the ECM [24,25,26,27]. MMPs can be anchored to the plasma membrane or secreted into the extracellular medium [27]. Studies have demonstrated that NOX-dependent ROS play a key role in this process. However, much of what is known about invadopodia formation is based on simplified in vitro models, which may not fully reflect the complexity of ROS dynamics and cellular interactions in vivo.
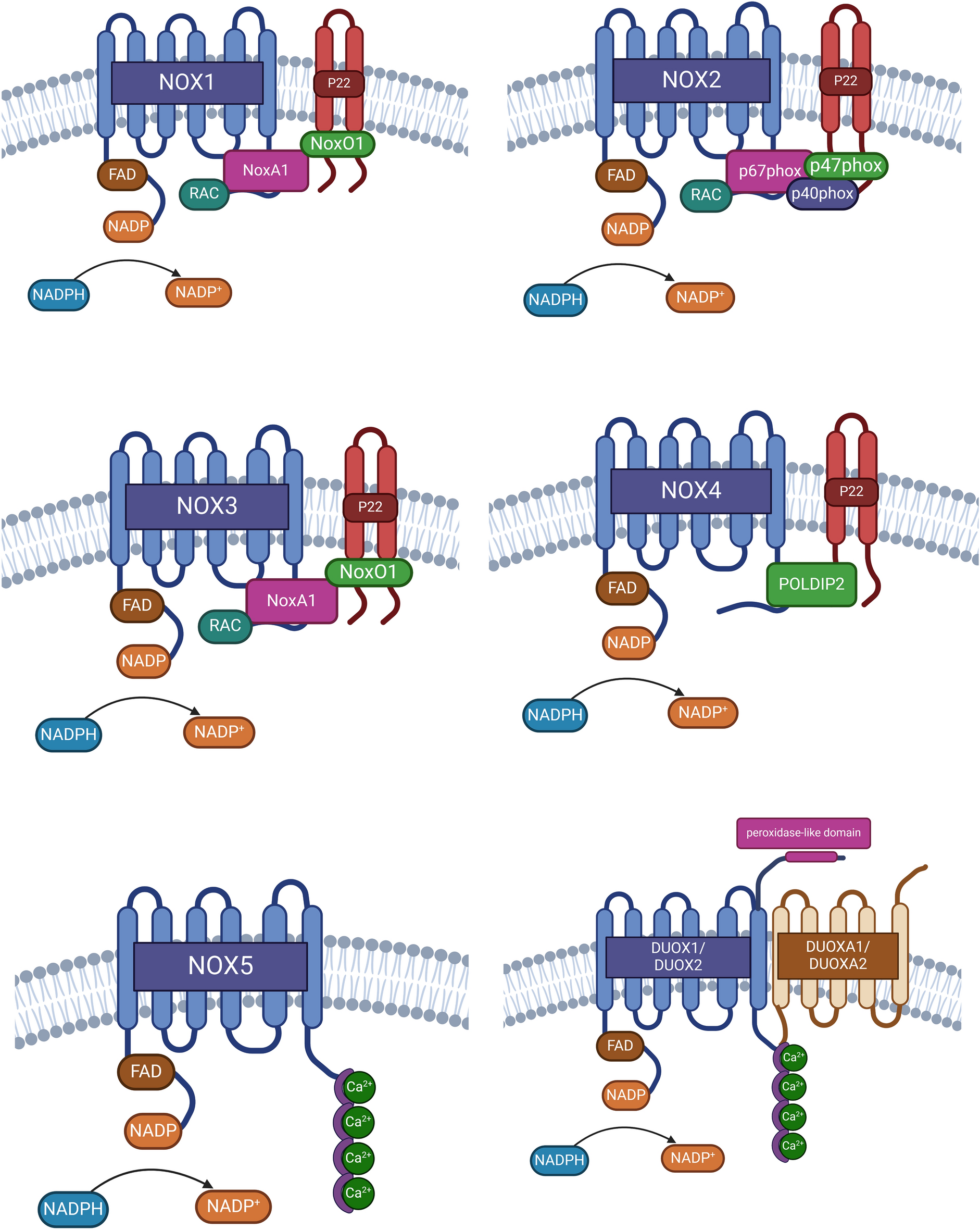
Structural features of NADPH oxidases and their role in ROS productionNADPH oxidases (NOX) constitute a family of membrane proteins and serve as one of the main sources of reactive oxygen species (ROS) in eukaryotic cells [28]. NOX proteins can be classified based on the type of ROS production. Specifically, NOX1-3 and NOX5 are known as superoxide anion (O₂⁻) producers, while NOX4 and DUOX1-2 are mainly related with the production of H₂O₂ [29]. ROS generation by NOXs usually involves the oxidation of NADPH molecules, wherein the electron from NADPH is transferred to a molecule of oxygen, resulting in the production of O₂⁻ and NADP + [30,31,32,33,34]. O₂⁻ is rapidly converted to hydrogen peroxide (H₂O₂) by the action of the antioxidant enzyme Superoxide Dismutase (SOD) through a two-step dismutation reaction. In the first step, O₂⁻ binds to the active site of SOD, where it transfers an electron to the SOD metal cofactor, leading to its reduction. This electron transfer disrupts the bonds between the metal cofactor and nearby histidine residues, inducing protonation of histidines and facilitating the release of molecular oxygen as the first product. In the second step, a new O₂⁻ binds to SOD's active site, receiving the electron from the previously reduced metal cofactor. This electron transfer promotes the protonation of the new O₂⁻, ultimately generating H₂O₂ as the final product and restoring the bonds between the metal cofactor and histidines in SOD [30, 31]. On the other hand, NOX4 directly produces H₂O₂, utilizing 90% of the electron flow, while the remaining 10% of the electron flow is used for O₂⁻ production [35]. For H₂O₂ formation, two electrons are sequentially transferred to an O₂ molecule, resulting in its double reduction. In the first reduction step of O₂ generates an O₂⁻ molecule, which interacts with the heme group of NOX4, forming a stable intermediate between the heme group and O₂⁻. This interaction facilitates a second reduction of O₂⁻, followed by protonation, leading to the formation of H₂O₂ [35, 36]. It has been proposed that H₂O₂ generation by DUOX1 and DUOX2 proteins occurs through the reduction of two O2 molecules, producing two O₂⁻ molecules. These O₂⁻ then undergo a dismutation reaction, leading to the formation of H₂O2 [37]. Despite the well-established role of NOX proteins in ROS production, the exact biochemical and molecular processes leading to their activation remain poorly understood, particularly with respect to isoform-specific functions in cancer cell invasion. Structurally, NOX proteins are composed of four transmembrane domains (TM) and an intracellular dehydrogenase domain (DH) [9, 29]. The TM consists of six transmembrane alpha helices connected by intra- and extracellular loops. Additionally, this domain is associated with two heme groups responsible for electron transfer to oxygen [10]. The DH contains binding sites for NADPH substrate, and FAD, facilitating electron transfer to the heme groups at TM domain [10, 29]. However, current research often overlooks how differences in the cellular localization and regulation of NOXs might influence their contributions to ROS production in different cellular contexts. Notably, the activation of NOXs at the membrane requires the formation of distinct complexes specific to each NOX isoform, as illustrated in Fig. 1. NOX1-4 proteins are associated with a membrane-associated protein, p22phox, which acts as a scaffold protein for the maturation and folding of the active NOX protein. Furthermore, for NOX1-3, p22phox functions as a platform for the binding of cytosolic activator proteins [10, 38]. In the case of NOX5, this variant does not form a complex with p22phox, and conversely, for DUOX1 and DUOX2, the presence of scaffold proteins DUOXa1 and DUOXa2, respectively, is imperative for their activation [38]. For NOXs requiring the recruitment of cytosolic proteins, the specific proteins involved vary depending on the type of NOX, in the case of NOX1/p22phox, the adapter protein NoxO1 is recruited, facilitating the binding of the activator protein of NOX1 (NoxA1) and Rac, which then activate the catalytic domain of NOX1. Conversely, NOX2/p22phox utilizes the adapter protein p47phox, enabling the binding of activator proteins p67phox and Rac [10, 38]. For NOX3, its low expression has hindered the determination of interacting proteins for activation, but it is suggested that its activation depends on NoxO1 and NoxA1, similar to NOX1 [24, 38]. Finally, while NOX4/p22phox acts constitutively, its activity is potentiated through the binding of the protein POLDIP2 [38, 39]. The formation of regulatory complexes controlling NOX activity at the membrane level generates localized ROS, for example, at invadopodia, thereby linking to the invasive capacity of tumor cells. Despite extensive studies on NOX activators, the complexities of their interactions with different subunits and activators have not been fully addressed, leaving gaps in our understanding of their precise regulatory mechanisms.
Fig. 1
Structural components of active NOX and DUOX enzymes. Key elements that form the active NOX and DUOX enzymes, which are crucial for the generation of reactive oxygen species (ROS). NOX1, NOX2, and NOX3 are shown to share common structural features and rely on cytosolic subunits for their activation. In contrast, NOX4 is depicted as being constitutively active, although it has been described as further modulated by interaction with POLIDIP2. The figure also highlights that NOX5 and the DUOX isoforms respond to changes in intracellular calcium concentrations
NOX-derived ROS in invadopodia function and cancer cell invasionLocalized ROS production by NOX proteins is crucial in invadopodia formation [40, 41]. Studies in melanoma have shown an increase in localized ROS within invadopodia, suggesting that this rise is due to the activity of NOX proteins present in these cellular structures [42]. In colon cancer, the presence of the NOX1 protein in invadopodia has been observed, and its inhibition impair invadopodia formation, suggesting that NOX1 and ROS production are essential for the formation of the invasive phenotype in these cancer cells [43, 44]. The activation of NOX1 in colon cancer is mediated by the SRC-dependent phosphorylation of Tks4 and Tks5, which interacts with NoxA1 (Fig. 2) [43,44,45,46,47]. In the case of NOX2, the activation mechanism is different because Tks proteins do not interact with the activator protein p67phox [46, 47]. NOX4 has been detected in invadopodia alongside F-actin, although the constitutive regulation mechanism of NOX4 remains unclear [42, 48]. In lung cancer, it has been discovered that DUOX1 is epigenetically silenced through DNA hypermethylation in its promoter region, leading to an increased expression of molecular markers associated with EMT [49]. On the other hand, in colon cancer, DUOX2 has been observed to increase ROS production due to elevated DUOX2 protein levels, which, in turn, promotes the expression of EMT-associated markers [50]. In colon cancer, NOX5 participates in the regulation of integrin-linked kinase signaling pathways, which are involved in cell adhesion and movement, correlating with the motility of tumor cells [51]. In breast cancer, it was observed that the expression of NOX5 is regulated by the transcription factor STAT5A, and depletion of NOX5 leads to a reduction in the invasive capacity of tumor cells [52]. In prostate cancer, increased expression of NOX5 is associated with elevated ROS levels and enhanced invasive and proteolytic capacity through the activation of HIF-1α and an increase in MMP14 levels [53, 54]. The increase in ROS induced by NOX proteins has been observed to stabilize HIF1α [55], this is mediated by the oxidation of cysteine residues present in the prolyl hydroxylase domain-containing protein 2 (PHD2), which is responsible for inactivating HIF-1α [56]. In colon cancer, an increase in ROS levels due to elevated expression of NOX1, induced by NF-kB activation, contributes to the adhesive capacity of tumor cells [57]. Moreover, in colon cancer, increased expression of NOX2 is related to the negative regulation of the MAPK signaling pathway and an increase in proteolytic activity through elevated levels of MMP7 [58]. In gallbladder cancer have been reported an increased expression of NOX1 in cancer-associated fibroblasts, correlating with an invasive phenotype and poor prognosis [59]. In gastric cancer, an increase in NOX2 expression is linked to enhanced invasiveness of tumor cells [60]. In renal cell carcinoma subjected to hypoxic conditions, heightened NOX4 activity has been documented, contingent upon the downregulation of MAPK. This culminates in heightened production of interleukin-6 (IL-6) and interleukin-8 (IL-8), which enhance invasion of tumor cells [61]. Additionally, high expression of NOX4 in gastric cancer is associated with positive regulation of MMP7 and increased invasiveness of tumor cells [62]. The specific function of each NOX in different tumor models is detailed in Table 1. It is worth noting that the vast majority of these studies correlate NOX expression with aggressive characteristics; however, very little is known about the mechanism by which NOX-dependent ROS production modulates this malignancy. Moreover, the lack of data from different experimental conditions or patient samples makes it difficult to generalize these findings, and further studies are needed to validate these results across diverse cancer types.
Fig. 2
Activation and function of NOX1 in invadopodia formation. Activation mechanism of NOX1 within invadopodia and its contribution to the localized production of reactive oxygen species (ROS). The figure highlights the formation of the NOX1 complex, which includes the recruitment of the cytosolic activator proteins NoxO1, NoxA1, Rac, and the p22p
留言 (0)