More than 50 years ago, loss of direct intercellular communication was first associated with cancer development and progression [9]. Since then, many studies have supported the concept that Cxs represent tumor suppressors [12]. In PCa, Cxs research has shown contradictory evidence regarding the biological role of some of the isoforms in the early and more advanced stages of the disease. Our study primarily analyzed the expression of Cx26, Cx32 and Cx43, because these isoforms have been described to be expressed in benign prostate epithelium and during prostate cancer progression [26, 27]. In addition, we also analyzed the expression of Cx37, Cx40 and Cx45 because these Cxs have been reported to be expressed in other cancer cell models [38,39,40,41,42], but have not been analyzed in PCa.
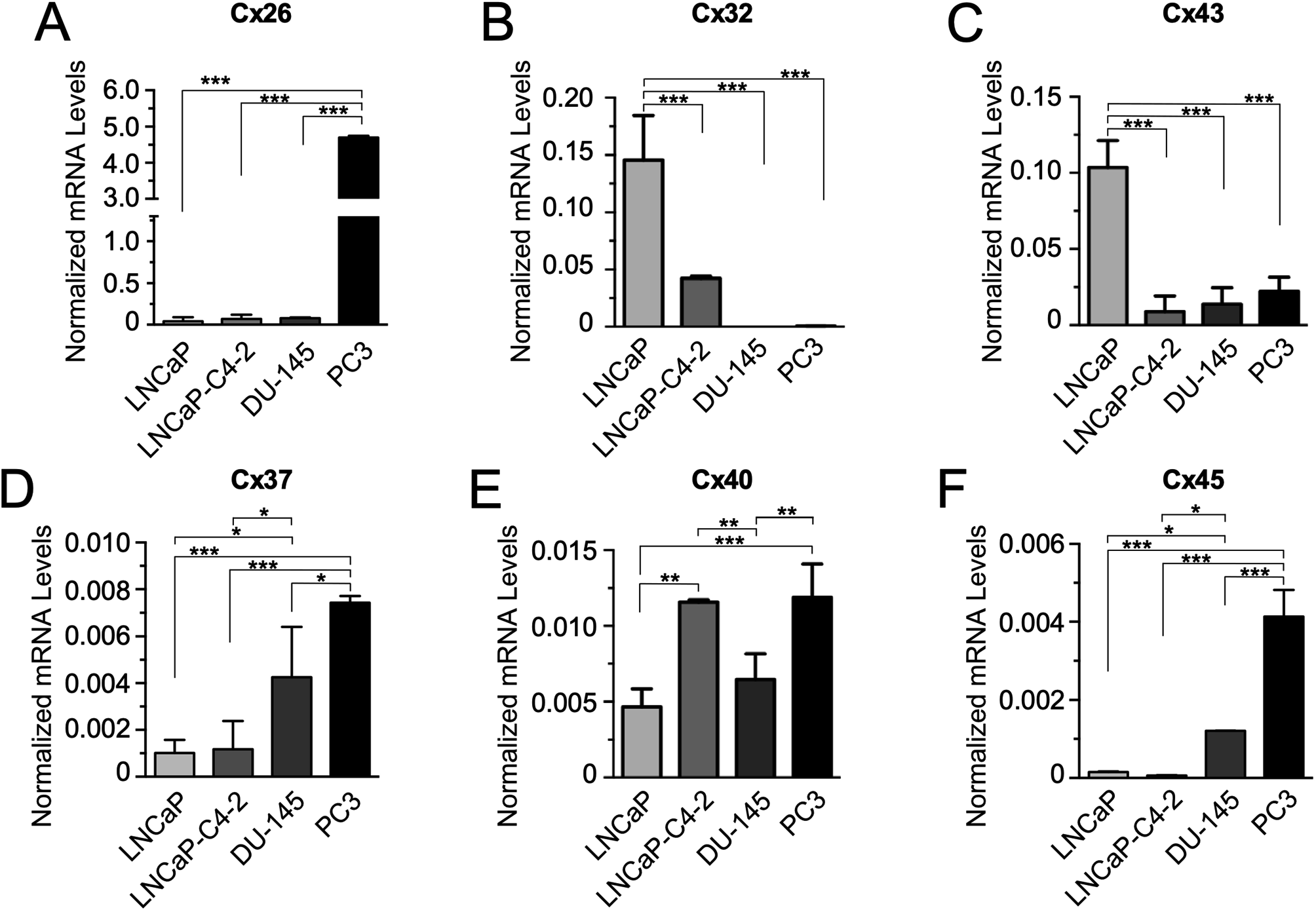
Our qRT-PCR results indicated that Cx26 mRNA is expressed at the highest level in the highly aggressive PC-3 cell line, when compared to the other PCa cell lines. This result might initially suggest that Cx26 could play a role in the advanced stages of PCa. However, when Cx26 protein levels were analyzed in the corresponding PCa cell lines, an opposite relationship was observed. In addition, opposite results were also observed when comparing Cx37 and Cx45 mRNA and protein expression levels in PCa cells (Figs. 1 and 2).
Similar to what was described by Tate et al. [28], we detected high levels of Cx26 protein in LNCaP cells. The fact that in our study LNCaP and LNCaP-C4-2 cell lines express high amounts of Cx26 protein, despite having low amounts of mRNA, could be explained by specific regulation of Cx26 expression at the post-transcriptional level. Protein synthesis is determined by the balance between mRNA degradation and stability, which is controlled by binding proteins or non-coding RNA to specific sequences within the mRNA [43]. In this case, it could be favoring Cx26 mRNA stabilization in these cell lines and, therefore, allowing a greater protein synthesis. Another explanation for this controversy might be related to the efficiency of Cx26 translation in PCa cell lines, which may result in high amounts of Cx26 protein despite having low amounts of Cx26 mRNA. Most mRNAs are translated by a cap-dependent canonical mechanism. However, some mRNAs might contain internal ribosome entry sites (IRES) that allow direct binding to the ribosome and their cap-independent translation. This mechanism has been reported in cancer cells under stress, apoptosis, among other conditions [13, 43]. Moreover, previous studies indicated that Cx26 mRNA contains IRES elements that induce its cap-independent translation in human pancreatic cancer cells [44].
At the protein level, we observed a clear inverse relationship between the expression of Cx32 and the aggressiveness of PCa cell lines. This finding aligns with prior research in human samples where Cx32 loss was associated with more advanced stages of prostate cancer [24]. Similarly, in hepatic cancer animal models treated with MitoQ [45], and in liver cancer, Cx32 has been shown to negatively regulate the expansion of liver cancer stem cells and, as expected, was positively correlated with patient survival [46]. Furthermore, in two breast cancer cell lines (MCF-7 and Hs578T), the enhancement of Cx32 expression resulted in reduced migration and invasion potentials [47]. Collectively, these examples underscore Cx32 as a broad-spectrum tumor suppressor, and PCa is no exception.
In the case of Cx43 mRNA and protein expression, our results indicated that the less aggressive PCa cell lines, LNCaP and LNCaP-C4-2, showed the highest expression of the mRNA and mRNA and protein, respectively, when compared to the more aggressive PCa cell lines, Du-145 and PC-3. These results contradict the results reported by Zhang et al. [15], which showed that the expression of Cx43, at the mRNA and protein levels, was positively associated with an increase in PCa malignancy. This controversy might be explained by differences in experimental conditions between both groups, such as using cells at different cell culture passages and/or confluences. In our case, our cell line models were used at very low passages (passage 1–5) after been commercially obtained from ATCC and all experiments were conducted with cells at no more than 80% confluence, though results were consistent even under higher confluence conditions (data not shown).
In addition, we focus on the analysis on the amount of mRNA and protein levels of Cx37, Cx40 and Cx45, which have not been studied in PCa, but have been described in other types of human neoplasms. A previous study showed that the C1019T polymorphism in the Cx37 gene correlates with the occurrence of gastric positive infection with Helicobacter Pylori. Subsequently, it was demonstrated that, by using a siRNA against Cx37 in subcutaneous gastric tumors grown in mice, tumor cell apoptosis was promoted [39, 48]. Other groups have found overexpression of Cx45 in clinical specimens of patients with acute myeloid leukemia (AML) [36], and oral squamous cell carcinoma (OSCC) [49]. We observed a positive relationship between the amount of Cx37 and Cx45 mRNAs and the aggressiveness of PCa cell lines (Figs. 1 and 2). However, Cx40 mRNA showed no clear relationship with the level of aggressiveness in PCa cell lines; LNCaP-C4-2 and PC-3 cell lines showed the highest level of expression of Cx40 mRNA. Once again, these differences observed at the mRNA level were not replicated at the protein level, observing a non-significant association between the expression level of the Cx37, Cx40 and Cx45 proteins and the degree of aggressiveness of PCa cell lines. These results show the need to unify criteria for the analysis of the expression of these proteins, considering aspects directly related to the cell types and their culture conditions.
Subsequently, we evaluated the ability of the different PCa cell lines to form functional GJCs. Interestingly, only the less aggressive PCa cell line, LNCaP, were able to moderately transfer the LY, which indicates that these cells possess some functional GJCs. Our results indicate that PCa cell lines that show greater tumor aggressiveness lose their intercellular communication mediated by GJs, which is in accordance with most of the functional studies carried out in cancer cells. In fact, Mehta et al. [27] analyzed cell-to-cell communication through GJCs in Du-145 and PC-3 cell lines, using the LY dye, and found similar results.
The moderate GJIC activity in the LNCaP cell line correlated with the high levels of expression of Cx26 and Cx43 observed in this cell line, which suggests that the intercellular channels could be constituted by one or both of these Cxs. Furthermore, the fact that only LY was transferred, can shed light on the type of Cx isoform involved in this communication. GJC have been proposed to exhibit Cx-type specific permeabilities to different substances. Ek-Vitorin et al. [50]. demonstrated that the negatively charged LY dye permeates through GJC composed of different Cxs types in the following the order of efficiency: Cx43 > Cx45 > Cx26 > Cx40, which suggests that Cx43 could be involved in GJIC in LNCaP cells. Our immunofluorescence analyses also suggest Cx43 as a potential mediator of the moderate GJIC activity observed in LNCaP cells. Even though we used two permeability tracers of different charges and sizes (LY: -2 and 443 kDa; Dapi: +1 and 279 kDa), adding more fluorescent tracers that differ in size or ionic charge, load, and stiffness to the functional study of GJs could help to solve this question with greater precision.
In the last decade, it has been found that hemichannels participate in various cellular processes, such as modulation of cell proliferation and survival [31]. However, their possible role in tumor growth has received less attention. Khalil et al. [30]. demonstrate that Cx43 hemichannels are functional and permit nucleotides and nucleosides release during the invasion process of breast cancer cells. In PCa cell models, the functionality of hemichannels formed by Cxs has not been evaluated. In our study, we measured the uptake of DAPI under basal conditions and in the presence of a medium without Ca+ 2 or Mg+ 2 as a stimulus to increase the open probability of Cx hemichannels [51, 52]. The fact that we did not observe a large amount of DAPI uptake under basal conditions is in accordance with the conservative concept that the hemichannels remain preferentially closed during cell-resting conditions [53]. In presence of medium lacking divalent cations, significant differences in dye uptake were observed in PCa cell lines. Du-145 and PC-3 cell lines showed low DAPI uptake, suggesting that the more aggressive PCa cell lines display low or no hemichannel activity. La+ 3 blocked DAPI uptake in the LNCaP PCa cell line by less than half. Nonetheless, in LNCaP-C4-2 cells, an effect of the La+ 3 was not visualized. Together, these results indicate that from all PCa cell lines analyzed, only LNCaP cells showed evident, but still moderate, hemichannel activity in vitro. This observation correlated with the findings at the level of GJIC and the expression patterns of Cx26 and Cx43 in LNCaP cells. More specifically, it matched the punctate localization of Cx43 at the plasma membrane of LNCaP cells, as determined by immunofluorescence. The fact that the highly aggressive, Du-145 and PC-3, PCa cell lines do not have functional GJC or hemichannel activity but do express certain Cxs such as Cx26, Cx43, Cx40 and Cx45 suggests that these isoforms could be fulfilling roles that could be independent of their capacity to form channels. Consequently, our immunofluorescence analyses revealed that Cx26 and Cx43 localized mostly at the cytoplasmic level in PC3 cells. Several studies have implicated a potential channel-independent activity of Cxs in supporting cell growth, differentiation, tumorigenesis, and apoptosis. Although, the mechanistic aspects of these actions remain poorly characterized [54, 55]. This channel-independent roles of Cxs have also been implicated in the regulation of migration and proliferation through direct Cx-protein interactions such as interactions of Cxs with the cytoskeleton. As an example, Cx43 induces migration of LNCaP cells that were exposed with osteoblasts-conditioned media, in a GJIC- and HC-independent manner [56].
Cxs are implicated in the activation or repression of the epithelial-mesenchymal transition (EMT) program in cancer cells [57, 58]. It is generally believed that connexins regulate EMT either as monomers or through GJs [59, 60]. For instance, Cx32 expression is downregulated in hepatocellular carcinoma (HCC) cells during carcinogenesis, and this decrease in Cx32 induces EMT by activating the Wnt/β-catenin/Snail pathway, leading to high invasion and metastasis characteristics in HCC cells [59]. Conversely, overexpression of Cx26 or Cx43 inhibits the proliferation of breast cancer cells in vitro and reduces tumorigenicity in vivo, promoting the transition from mesenchymal to epithelial states [61]. Additionally, overexpression of Cx43 increases the expression of E-cadherin and ZO-1 epithelial markers, while downregulation of Cx43 increases the expression of the N-cadherin mesenchymal marker in MDA-MB-231 cell lines [58, 62]. These findings indicate that connexins play a crucial role in regulating EMT in cancer progression and metastasis, highlighting their potential as biomarkers for cancer treatment and prognosis.
Even though most studies have focused on determining the channel-independent roles of Cx43 and 32 in cancer, our results also pointed out to Cx40 or Cx45 as potential candidates for modulating, in a GJC- and hemichannel-independent manner, the biology of highly aggressive PCa cell lines. Consequently, future studies should focus on evaluating the role of Cx40 and Cx45 in the progression of PCa and whether or not these proteins can affect cell proliferation and/or metastatic potential.




留言 (0)