
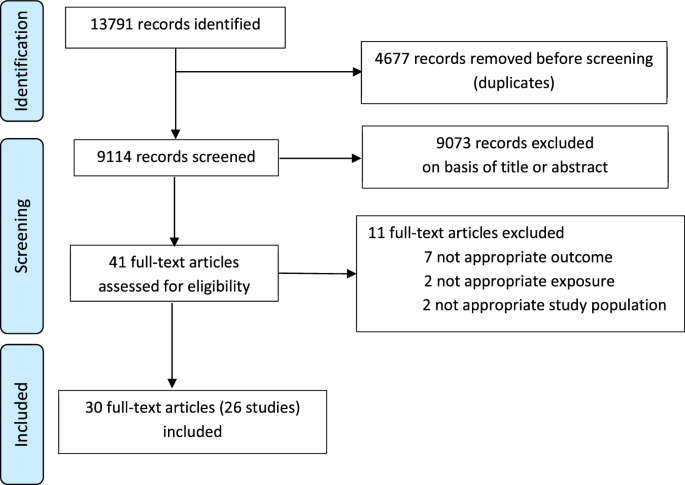
記住我

Ab1456 and Ab1271 were isolated from NHPs after sequential immunizations with engineered or wildtype SOSIP-based immunogens designed to target the V3-glycan patch on the gp120 subunit of Env and characterized as weak, but heterologously-neutralizing mAbs14,15. NHPs were primed with RC1-4fill, a low affinity V3-glycan patch germline-targeting immunogen conjugated to virus-like particles (VLPs) using the SpyCatcher-SpyTag system17,18. RC1-4fill is a modification of the clade A BG505-based 11MUTB SOSIP immunogen19, in which the N156gp120 glycan was removed (N156Q) and potential N-linked glycosylation sites (PNGSs) to block BG505 strain-specific responses to an immunodominant glycan hole in the vicinity of residue 241gp12020 were added. A series of boosts consisting of VLPs presenting 11MUTB-4fill15, a clade B B4121 or B41-5MUT15, a mosaic of a clade B (AMC011)22 and clade C (Du422)23, and a mosaic of consensus Envs from Group M and clade C (ConM/ConC)24,25 were given to try to shepherd antibody responses towards broader reactivities and avoid strain-specific responses (Fig. 1a).
Fig. 1: Characterization of Ab1456 and Ab1271.
a Schematic describing the sequential immunization of NHPs that gave rise to Ab1456 (Regimen 1 in boost 2; R1) and Ab1271 (Regimen 2 in boost 2; R2)15. Ab1456 was isolated from NHP DGJI, and Ab1271 was isolated from NHP T15. b Alignments of Ab1456 and Ab1271 to their presumptive germline VH gene precursors, as identified by IMGT/V-QUEST67,68. CDRs are defined according to Kabat69.
Using single B-cell cloning techniques, Ab1456 and Ab1271 were isolated after the third boost in the same two NHPs as the Ab1573 and Ab1303 CD4bs mAbs15 (Fig. 1a). In contrast to Ab1573 and Ab1303 competition results, Ab1456 and Ab1271 each competed with the V3 bNAb 10-1074, suggesting on-target binding specificities for these mAbs15. However, structural characterizations of Ab1456 and Ab1271 remained elusive. Both mAbs displayed heterologous neutralization when tested against a panel of 19 pseudoviruses including the 12-strain global HIV-1 panel26 and two SHIVs, neutralizing 6 of 19 (Ab1456) or 14 of 19 (Ab1271) HIV-1 pseudoviruses with IC50 values < 100 µg/mL15. While neutralization potencies were generally weak, Ab1271 in particular exhibited breadth, neutralizing all viruses tested at IC50 values < 500 µg/mL. Ab1456 was derived from macaque IGHV5-43*02 and IGLV1-81*01 germline V gene segments, exhibiting 14.3% (heavy chain; HC) and 7.1% (light chain; LC) amino acid changes due to somatic hypermutation. Ab1271, derived from the IGHV4-92*01 and IGLV1-85*01 germline V gene segments, exhibited 15.5% (HC) and 8.2% (LC) changes from somatic hypermutation (Fig. 1b). Of note, there is a one-residue deletion in the HC framework region 1 (FWRH1) of Ab1271.
Ab1456 recognizes CD4-bound open Env trimer conformationsFor structural studies of Ab1456 recognition of Env, we formed Fab-Env complexes using a chimeric SOSIP Env containing a gp120 derived from JRCSF.JB, an HIV-1 strain that was potently neutralized by Ab145615, and a gp41 derived from BG50527. Fab-SOSIP complexes isolated by size-exclusion chromatography (SEC) were used for EM analysis. Initial processing in cryoSPARC28 yielded a 6.6 Å resolution structure that showed targeting of the trimer apex of an open Env conformation with an apparent stoichiometry of one Fab per trimer (Fig. 2a, b). In this structure, two of three protomers in the trimer appeared to adopt a CD4-bound open conformation as indicated by rearranged V1V2 densities, while the third protomer exhibited an outward rotation, but lacked the V1V2 rearrangement29. Overall, this state of the Env trimer resembled the conformation of HT2, a SOSIP heterotrimer in complex with two, rather than three, copies of soluble CD46; hence, we refer to this Ab1456-bound Env conformation as HT2-like, noting, however, that the Ab1456-Env structure was determined in the absence of CD4 and with a homotrimeric SOSIP (Fig. 2a, b). Unexplained density was present at the trimer apex (Supplementary Fig. 1a), suggesting that this consensus structure included particles from distinct 3D classes. Indeed, 3D classification in RELION30 revealed extensive heterogeneity. Structural classes were identified that differed in the conformational state of the trimer, the number of bound Fabs per trimer, the relative positioning of the bound protomers, and the approach angles of the Fabs (Fig. 2). Sorting of approximately 80,000 particles allowed us to determine eight structural classes with resolutions ranging from 8.8 Å to 14 Å (Supplementary Fig. 2). Given the high degree of heterogeneity, additional states could exist. In addition, imperfect separation of particles may bias some of the reported structures.
Fig. 2: Ab1456 Fab binds open forms of HIV-1 Env.
a Schematic of top-down views of Env trimers showing a potential pathway from a closed (left) to a fully-open (right) trimer. The three open Env states were identified in structural classes with bound Ab1456 Fab(s). b Results of a non-uniform refinement of Ab1456 Fab-bound JRCSF SOSIP particles prior to 3D classification. A schematic representation of the top-down view of the trimer conformation is shown in this and subsequent panels with the Ab1456-bound protomer(s) indicated by a black dotted pattern and displaced V1V2 loop(s) indicated by orange dot(s). c Left and middle: Two structural classes showing three Ab1456 Fabs bound. Right: An overlay of the two structural classes. d Three structural classes showing two Ab1456 Fab-bound trimers. e Three structural classes showing one Ab1456 Fab-bound trimers. f Analysis of the Ab1456 epitope. Protomer A from PDB 8FYJ (HT2 Env heterotrimer complexed with two CD4s) and 3MLX (human mAb 3074 complexed with a V3 peptide; ordered residues 305-320) were independently docked into the density corresponding to protomer A and Ab1456 Fab/V3 peptide in the consensus map, respectively. g Mass photometry of JRCSF SOSIP (blue) and JRCSF SOSIP in complex with Ab1456 Fab (gold).
Two different three Fab-bound HT2-like trimer classes were identified. In these classes, the Fab bound to protomer B was wedged either in front, or behind, of the Fab bound to protomer C, resulting in distinct angles of approach (Classes 1 and 2) (Fig. 2c).
Multiple structural classes of Env trimers with two bound Fabs were also found. Two distinct classes of two Fab-bound HT2-like trimers were identified with the Ab1456 Fabs binding to different configurations of the Env A, B, and C protomers. In one configuration, both Fab-bound protomers (protomers A and B) adopted the CD4-bound open conformation (Class 3) (Fig. 2d). In another structural class, both the protomer that lacked apparent V1V2 rearrangements (protomer C) and an adjacent protomer in the CD4-bound open conformation (protomer A) exhibited bound Ab1456 Fabs (Class 4) (Fig. 2d). Another structural class showed two Ab1456 Fabs bound to a fully-open trimer in which all three protomers displayed V1V2 rearrangements (Class 5) (Fig. 2d).
In Env structures exhibiting a single bound Fab, three different trimer conformational states were identified (Fig. 2a, e). In one state, only the Fab-bound protomer adopted a CD4-bound open conformation (as defined by a V1V2 rearrangement), and the other protomers exhibited neither an outward gp120 rotation nor V1V2 rearrangement (Class 6) (Fig. 2e). Another class was found in which the Env trimer adopted an HT2-like state, and only protomer A was bound (Class 7) (Fig. 2e). Finally, a structure of a single Ab1456 Fab bound to a trimer in which all three protomers adopted a CD4-bound open conformation was identified (Class 8) (Fig. 2e).
At the resolutions of our EM structures, we are mostly limited to analyzing the Ab1456 epitope through docking of previously-determined EM and X-ray structures. Docking of protomer A from the HT2 trimer6 (Env heterotrimer bound by 2 copies of soluble CD4; PDB 8FYJ) into the consensus structure revealed qualitative agreement with protomer A density in the EM map (Supplementary Fig. 1b). The docked protomer showed apparent Ab1456 targeting of V3 residues that were not built in the HT2 structure as a consequence of being disordered in PDB 8FYJ. To account for additional Ab1456 and Env V3 density in the Ab1456-JRCSF Env structure, we docked a crystal structure of the human mAb 3074 in complex with a V3 peptide spanning gp120 residues 301-324 (PDB 3MLX; residues 305-320 were ordered in the crystal structure). We chose this peptide/V3-antibody structure because 3074 neutralizes viruses in common with Ab1456 (e.g., both neutralized JRCSF, 6535.3, and X1632 to a greater extent than other strains15,31) and preferentially binds Env in the presence of CD432. While these properties may indicate that 3074 and Ab1456 make similar contacts with Env, such interactions could be mediated by different antibody features (e.g., different complementarity-determining regions, different specific Fab-Env interactions, different Fab binding orientations, etc.). The docked Fab-bound V3 peptide fit the EM density well, providing support for the interpretation that Ab1456 and mAb 3074 contact similar V3 residues that are exposed on open states of the trimer (Fig. 2f). Ab1456 targeting of the V3 epitope is consistent with it competing for Env binding with the human V3-directed bNAb 10-107415.
To further characterize the stoichiometry of Ab1456 Fab binding to Env trimer, we performed mass photometry, a technique that detects binding interactions in solution via mass measurements of individual molecules33,34. Unlike the Ab1456-JRCSF SOSIP complex used for cryo-EM, the Fab-Env sample for mass photometry was not purified by SEC, and mass photometry was performed at a more dilute final concentration (diluted from ~1 mg/mL to <1 µg/mL for measurement) than what was imaged by cryo-EM (~1.1 mg/mL). By mass photometry, the JRCSF trimer appeared at ~300 kDa, whereas a second population of approximately one third the mass (experimentally measured as ~80 kDa) is assumed to be JRCSF monomer, which was also observed in the SEC profile of this SOSIP (Supplementary Fig. 3). In agreement with observations by cryo-EM, the Ab1456-JRCSF sample showed particles with increased mass relative to the trimer alone control (Fig. 2g). Although distinct populations could not be unambiguously identified, the masses of the complexes were consistent with a mixture of Fab-Env particles containing either 0, 1, 2, or 3 bound Fabs per trimer (Fig. 2g).
Ab1271 recognizes a conformation distinct from pre-fusion closed Env trimersWe next focused on Ab1271, which exhibits a broader neutralization profile than Ab145615. For these structural studies, we formed Fab-Env complexes using a chimeric SOSIP with a gp120 derived from 6535.3, a tier 1 virus that was potently neutralized by Ab127115, and a gp41 derived from BG50527. Cryo-EM analysis revealed trimers that were not complexed with Ab1271 Fab, yielding a 4.6 Å structure of the unliganded 6535.3 SOSIP trimer (Fig. 3a, Supplementary Fig. 4). Although the 6535.3 Env conformation generally resembled a typical closed prefusion trimer in that gp120s were not outwardly rotated and the V1V2 regions were not displaced to the trimer sides as seen in the CD4-bound open Env conformation11,29,35, the trimer apex in 6535.3 Env differed from those of BG505 and other SOSIP Env structures. Docking of a closed BG505 SOSIP (PDB 6UDJ) into the 6535.3 SOSIP density showed differences in the presumptive locations of the V1V2 regions at the trimer apex for two of the three 6535.3 protomers, and density for V1V2 on the third protomer was not observed (Fig. 3a).
Fig. 3: Ab1271 recognizes a closed 6535.3 Env trimer with an altered apex.
a Side and top-down views of a structural class of unliganded 6535.3 SOSIP. BG505 SOSIP (PDB 6UDJ) (tan cartoon representation) was docked into 6535.3 density. Presumptive V1V2 density for two of the three 6535.3 protomers (pink and purple) is shifted relative to the BG505 V1V2, as indicated by white arrows in the top-down view on the right. b A structural class of Ab1271-Env complex with a single copy of Ab1271 Fab bound to the 6535.3 SOSIP. c Left: Ab1271-6535.3 complex. Middle: Close-up of Fab-Env interaction showing apparent targeting of Ab1271 Fab towards the V3 GDIR motif (G324gp120 – R327gp120). Right: Close-up of Fab-Env interaction showing apparent clash with V1V2 residues of the docked BG505 SOSIP. d Mass photometry of 6535.3 SOSIP (blue) and 6535.3 SOSIP in complex with Ab1271 Fab (gold).
In addition to the unbound 6535.3 trimer structure, we determined a 6.3 Å structure of a 6535.3 SOSIP bound by a single Ab1271 Fab (Fig. 3b). In this structure, the Env portion of the density closely matched the density for the unbound 6535.3 SOSIP (Fig. 3a, b), with Ab1271 interacting with the V3 region of the protomer lacking apparent V1V2 density. The apex of a docked closed BG505 trimer (PDB 6UDJ) clashed with density corresponding to the Ab1271 Fab, suggesting that V1V2 must be in a position distinct from its position in BG505 in order to accommodate Ab1271 binding (Fig. 3c). Unlike the occluded-open Env conformations found in the Ab1303-Env and Ab1573-Env complex structures12 or the CD4-bound open structures with Ab1456 (Fig. 2), the Env trimer in the Ab1271-Env complex did not exhibit an outward rotation of its gp120s, in common with conventional pre-fusion closed SOSIP trimer structures3. However, as the form of the Env trimer in the Ab1271-6535.3 SOSIP complex is distinct from previously-determined closed trimer structures3, we refer to its conformation as apex-altered closed. While low resolution limited our analysis of antibody epitope details, the docked structure revealed apparent Ab1271 targeting at or near the conserved GDIR motif within the V3 loop of gp12036 (gp120324-327) (Fig. 3c).
To further investigate the stoichiometry of Ab1271 Fab binding to the Env trimer, we evaluated Ab1271 Fab-6535.3 SOSIP complex formation using mass photometry. Similar to the JRCSF SOSIP, the 6535.3 SOSIP also included multiple populations, predominantly corresponding to SOSIP trimers and protomers (Fig. 3d; Supplementary Fig. 3). Although a majority of the trimers remained unbound in the presence of Ab1271 Fab, a small shoulder with a mass ~50 kDa greater than the trimer alone control appeared in the experimental histogram of the Ab1271-6535.3 SOSIP sample, consistent with a population of one Fab-bound trimers (Fig. 3d). Additionally, excess Fab (~50 kDa) likely contributes to the large increase in the size of the peak experimentally measured at ~70 kDa. As mass photometry experiments were conducted at more dilute final concentrations (diluted from ~1 mg/mL to <1 µg/mL) compared to cryo-EM (~1.9 mg/mL), this result suggests a weak affinity and/or fast off-rate of Ab1271 Fab for the 6535.3 SOSIP trimer.
Antibody-virus pre-incubation is not necessary for in vitro neutralization by Ab1456Based on the cryo-EM structures of the weakly neutralizing NHP mAb Ab1456 that revealed recognition of open Env conformations, we reasoned that Ab1456 targeting could be limited by the conformational availability of the epitope on virion-bound Env trimers. Standard TZM-bl neutralization assays provide a time window, typically 1 hour, in which an antibody is incubated at 37 °C with the virus prior to the addition of target cells37,38. We hypothesized that this incubation could allow sampling of open trimer conformations, which could then be captured, permitting antibody binding to virion Envs to achieve neutralization. The antibody-virus co-incubation step is distinct from how antibodies neutralize HIV-1 in vivo, where antibodies and viruses are not pre-incubated in a small volume and where there might only be a limited time window for an antibody to recognize Env on a virus prior to encountering a target cell. We therefore reasoned that pre-incubation of antibody and virus might artificially inflate the neutralization potencies of antibodies that target an epitope on an open Env trimer.
To test this possibility, we compared the 50% inhibitory concentrations (IC50s) of Ab1456 and other weakly and broadly neutralizing mAbs in the standard TZMbl assay, in which virus and antibodies were preincubated for 1 hour37,38, and in a modified assay, in which antibodies were first added to the cells followed by virus addition in a separate step. Selecting a set of both sensitive and more resistant viruses15, we found that the neutralization potencies of Ab1456 were very similar regardless of whether the standard or modified assay was used (Fig. 4a). This was true for HEK293T-derived pseudoviruses and replication-competent simian-human immunodeficiency viruses (SHIVs) as well as a SHIV challenge stock that was propagated in rhesus macaque peripheral blood mononuclear cells (PBMCs)15,39. Preincubation also had no effect on the neutralization potencies of other antibodies known to target open Env conformations, such as the CD4-induced antibody 17b2 and the linear V3 mAb 307440 (Fig. 4b). The slopes of the 17b and 3074 neutralization curves, like those of Ab1456, were more shallow than the slopes of potent bNAbs, such as the V3 bNAb 10-107441 and the CD4bs bNAb VRC0142 that recognize closed Env conformations (Supplementary Fig. 5). Since shallow dose-response curves are associated with less favorable therapeutic potentials43, these results imply limited in vivo protection efficacy for Ab1456.
Fig. 4: Preincubation of antibody and virus during in vitro neutralization does not affect neutralization potency.
a Sensitivity of viruses expressing the DH12-V3AD8 (SHIVDH12-V3AD8; red), AD8-EO (SHIVAD8EO; green), and BG505.N332 (blue) Envs to neutralization by Ab1456 using a TZMbl assay with (left) and without (right) a 1 hour antibody and virus preincubation step. Neutralization curves are shown for pseudovirus (PV) and replication-competent forms of SHIVs derived either by HEK293T transfection (293T) or following propagation in rhesus macaque PBMCs (rh PBMC). Dotted lines indicate 50% reduction in virus infectivity. b Sensitivity of the viruses shown in a (listed on the left) to neutralization by other mAbs (listed on top) with (left panel) and without (right panel) a 1 hour antibody and virus preincubation step. 50% inhibitory concentrations (IC50) are shown in μg/mL (coloring indicates relative neutralization potency).
留言 (0)