
記住我





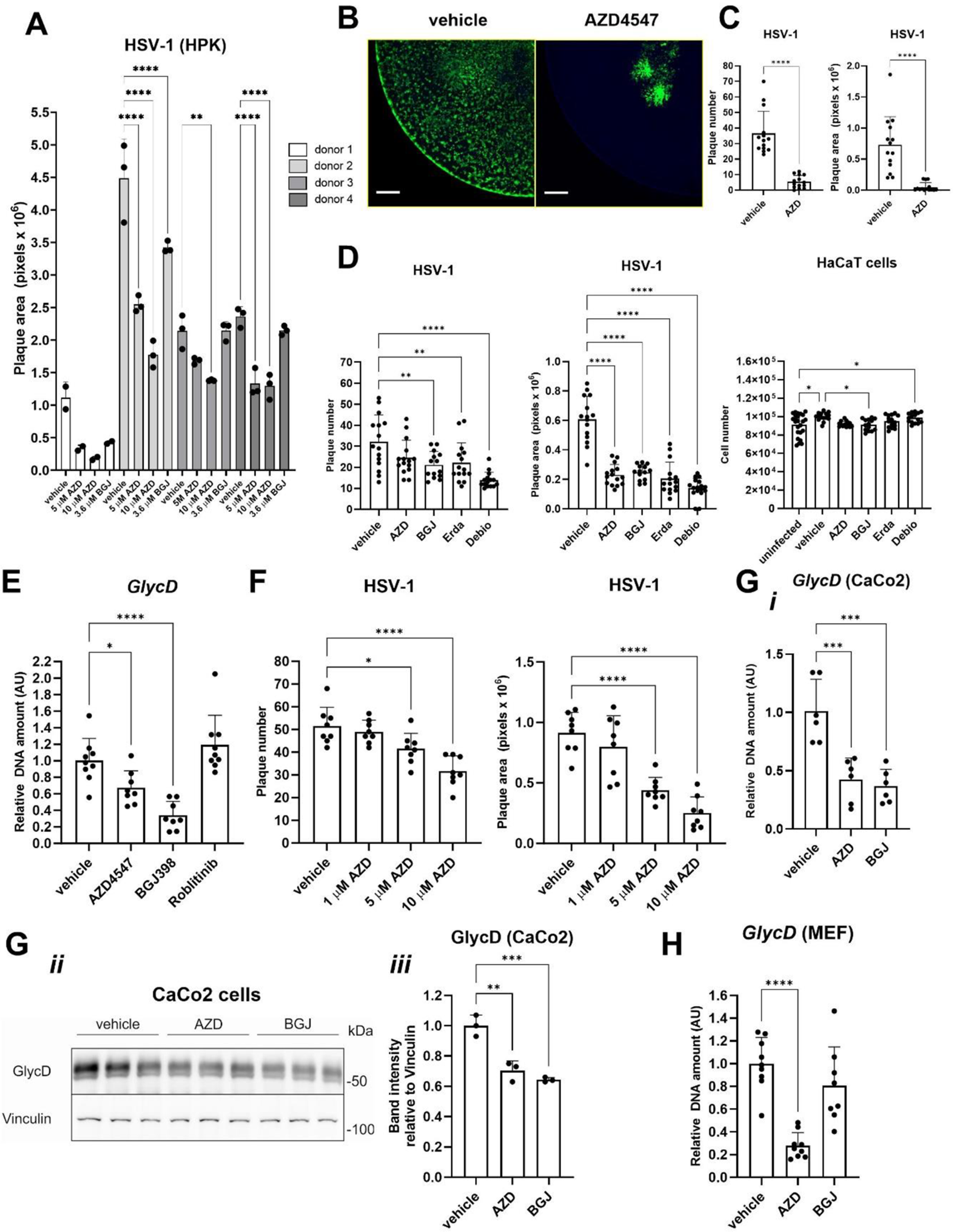
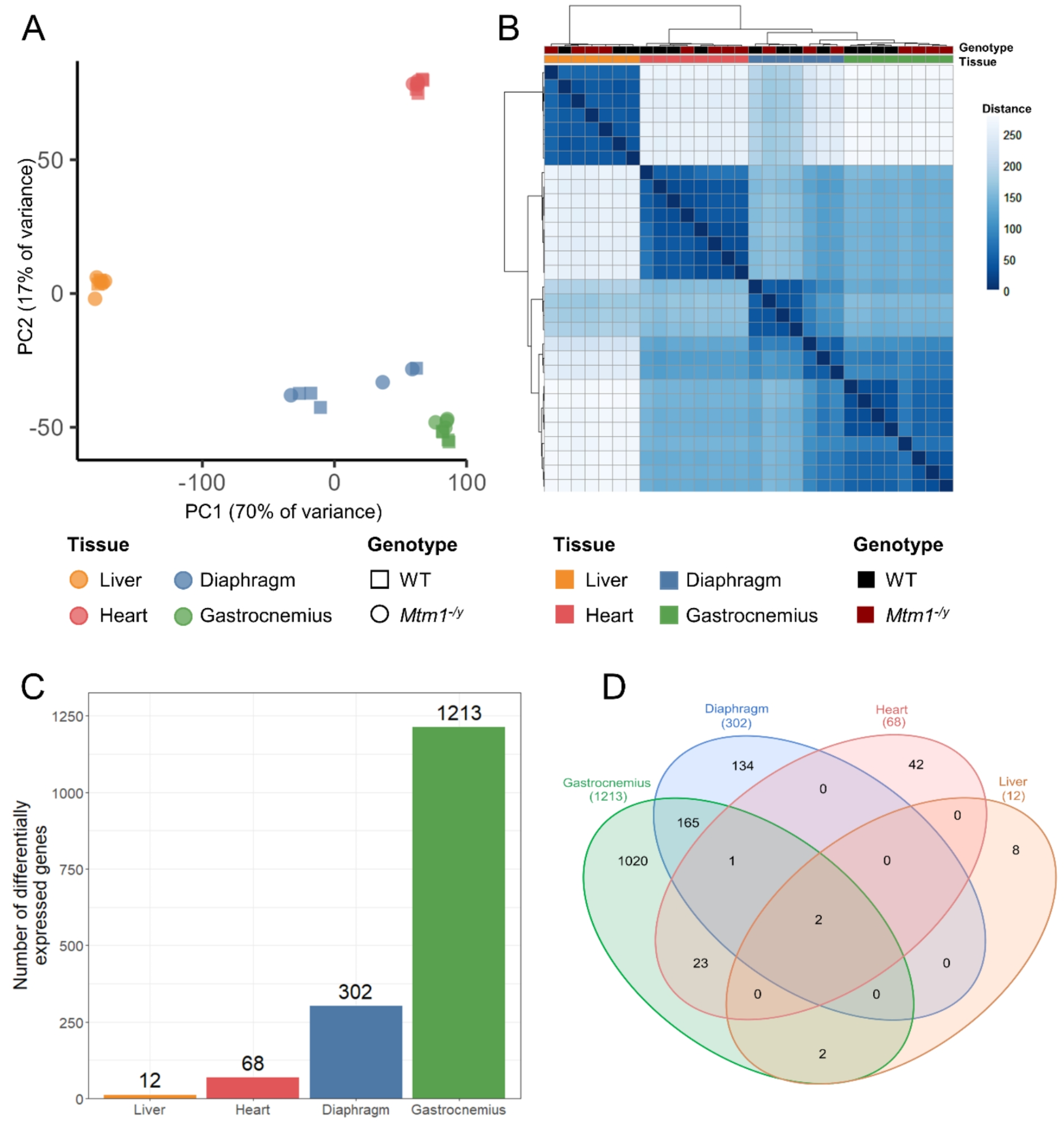
To assess the impact of XLCNM on gene expression in different muscle and non-muscle tissues, we performed RNA-seq in Mtm1−/y mice and their wild-type (WT) littermates at 5 weeks, a time point where Mtm1−/y mice display muscle weakness without been moribund. Gene expression was quantified in the heart, the liver, and two skeletal muscles: the gastrocnemius and the diaphragm. Exploratory data analysis revealed that clusters of samples derived from gene expression accurately recapitulated organ identity (Fig. 1A-B). Indeed, three clusters which correspond to skeletal muscles, heart, and liver samples were identified. However, the WT and Mtm1−/y samples were not clearly separated within each cluster. When considering all organs together, genotype separation only appeared in the gastrocnemius samples.
To evaluate the tissue-specific differences between WT and Mtm1−/y mice, we performed differential expression analysis (Fig. 1C). The threshold used to define differentially expressed genes (DEGs) was set at absolute log2-fold change > 1 and adjusted P value < 0.05. Gastrocnemius was the tissue with most dysregulation (1213 DEGs), followed by diaphragm (302 DEGs). Gene expression was less affected in heart (68 DEGs) and liver (12 DEGs). Among these dysregulated genes, only two were common to all tissues: Mtm1 and Arrdc2, which were both down-regulated (Fig. 1D, Supplementary Datasets S1-4). The observed downregulation of Mtm1 in all tissues validated the methodology. The downregulation of Arrdc2 ranged from a log2FC of -1.25 in the heart to -1.70 in the liver, with a log2FC of -1.45 and − 1.46 in the gastrocnemius and the diaphragm. Arrdc2 is a mechanosensitive gene, which is downregulated during anabolic stimuli and upregulated during catabolic stimuli in skeletal muscle, its induction leading to thinner myotubes [41, 42]. However, its function in liver and heart is unknown. No dysregulation of Dnm2 was observed, similarly to previous studies in TA [43]. Apart from these two genes, the liver had no dysregulated genes in common with heart or diaphragm, and only two additional dysregulated genes in common with the gastrocnemius. Similarly, the heart had few dysregulated genes in common with the skeletal muscles (gastrocnemius and diaphragm). On the other hand, skeletal muscles share 168 DEGs, which represents 55% of the dysregulated genes found in diaphragm.
Overall, these data indicate that the effect of XLMTM on the transcriptome is highly organ-specific, both in the number of differentially expressed genes and in the specific genes that undergo alteration. To gain a better understanding of the impact of the disease, we further studied each organ separately.
The Mtm1 -/y mouse model shows no liver defectsWe investigated whether the Mtm1−/y mouse model of XLMTM exhibited hepatic dysfunction similarly to what has been reported in patients. We first studied liver histology following hematoxylin and eosin (H&E) stain. Similar to their WT littermates, the Mtm1−/y mice did not exhibit excessive inflammation, fibrosis, steatosis, or necrosis (Fig. 2A). They presented a normal liver architecture into lobules with portal veins, hepatic artery and bile ducts at the periphery, and a central vein at the center. No accumulated bile was visible in the hepatocyte cytoplasm or canaliculi. Immunofluorescent staining of the bile salt export pump (BSEP) showed normal localization in the caniculi, excluding the cholestatic pattern of injury found in patients and mtm zebrafish characterized by a quasi-total loss of BSEP expression in the liver [16, 19] (Fig. 2B). Similarly, no liver defects were observed in the histology and BSEP immunofluorescence of 8-month-old Mtm1−/+ female mice (Supplementary Figure S1).
Fig. 4
Myogenic development is impaired in the skeletal muscles of Mtm1−/ymice. A Myh3 transcript relative expression. B Representative immunofluorescent staining images of the embryonic myosin (MYH3, green) and WGA (pink) in the tibialis anterior and the gastrocnemius (left), and MYH3 signal intensity quantification (right). Scale bars 50 μm. C Mstn transcript relative expression. D Fst transcript relative expression. For panels A, C, and D, transcript expression levels were obtained by RT-qPCR from tibialis anterior (2 weeks and 7 weeks), gastrocnemius and diaphragm (5 weeks) Mtm1−/y and WT samples, and are shown as relative expression compared to the average of the WT. ns P > 0.05, *P < 0.05, **P < 0.01, ***P < 0.001
As the morphology of the liver of Mtm1−/y mice was normal, we proceeded to assess hepatic function by measuring liver enzyme levels in the blood. The levels of bilirubin, aspartate aminotransferase and alanine aminotransferase were within normal ranges in all mice, and no statistically significant differences were found between Mtm1−/y and WT mice (Fig. 2C).
Finally, as no defects were detected at the morphological and functional levels, we analyzed RNA-seq data obtained from liver samples. Principal component analysis showed that the first 6 principal components (cumulatively, 99% of variance explained) did not separate the genotypes, which indicated that the WT and Mtm1−/y liver samples were highly similar. Indeed, differential gene expression analysis revealed little to no dysregulation in Mtm1−/y samples compared to WT. Only 12 differentially expressed genes were identified: Mtm1, Arrdc2, Myl2, Hhex, Myh6, Tmc7, Jaml, Setdb2, Pim3, Zfp36, 4632427E13Rik, and Phf11c (Fig. 2D; Supplementary Dataset S1). Due to the low number of DEGs, we then conducted gene set enrichment analysis (GSEA) in order to detect modified biological pathways. Only a small number of gene sets displayed significant enrichment, and none of these were pertinent to hepatic function (Supplementary Figure S2). Quantification of the protein level of DNM2 showed no difference between Mtm1−/y and WT livers (Supplementary Figure S3).
Overall, we characterized hepatic morphology, function, and gene expression in Mtm1−/y mice, and found no significant differences compared to their WT littermates. This indicates that the unchallenged Mtm1−/y mouse model of XLMTM does not replicate the liver defects observed in patients.
Comparison of 3 skeletal muscles underlines a common disease signatureUnlike the liver, skeletal muscles are severely affected in the Mtm1−/y mouse [8, 22, 23] (Supplementary Figure S4). Previous studies have extensively characterized gene expression dysregulation in tibialis anterior [23, 24], a fast—contracting skeletal muscle of the lower leg, but transcriptomic studies of other skeletal muscles are lacking. Therefore, we compared the RNA-seq data we obtained in the gastrocnemius and the diaphragm at 5 weeks with previous RNA-seq data obtained in the tibialis anterior at 2 and 7 weeks [23] to evaluate how MTM1 loss affects different skeletal muscles. Principal component analysis of the cardiac and skeletal muscles samples showed that the tibialis anterior samples clustered properly with the other hindlimb muscle without requiring batch-effect correction (Supplementary Figure S5). By comparing different ages, we aimed to define a fundamental and time-independent common signature.
Principal component analysis revealed a separation according to the different skeletal muscles on the first principal component (45% of variance) and the age on the second PC (16% of variance). Separation of the genotypes appeared when combining the second and third principal components, and was stronger for the tibialis anterior and the gastrocnemius than for the diaphragm (Fig. 3A). Compared to other tissues, the three skeletal muscles showed a high number of DEGs, ranging between 302 and 1722 (Fig. 3B, Supplementary Datasets S3-6). GO term over-representation analysis performed on each muscle identified 238 to 1428 enriched terms with an overlap of 104 terms enriched in all skeletal muscles and related to skeletal muscle development and function (Supplementary Figure S6, Supplementary Datasets S7-10).
In total, there were 132 common DEGs affected in the three studied skeletal muscles at any age (Fig. 3C, Supplementary Dataset S11). Enrichment analysis revealed that these genes were associated to GO terms previously identified in the tibialis anterior such as skeletal muscle development and function, extracellular matrix, and inflammation, but also revealed new affected pathways: calcium homeostasis and protein kinase B (AKT1) signaling, and negative regulation of cell development (Fig. 3D, Supplementary Dataset S12). Investigation of the cell development pathways revealed an upregulation of the fetal myosin Myh3, which was confirmed by RT-qPCR and immunofluorescence microscopy (Fig. 4A-B). Moreover, the upregulation of Fst and downregulation of Mstn were identified on the RNA-seq data and confirmed by RT-qPCR (Fig. 4C-D).
Fig. 5
The Mtm1−/y mouse heart shows no structural defects but is affected at the transcriptomic level. A Representative heart histology of WT and Mtm1−/y mice (H&E). Scale bars 100 μm. B Representative heart electron microscopy of WT and Mtm1−/y mice (H&E). Scale bars 2 μm. C PCA plot of heart samples, PC1 and PC4 are shown. D Volcano plot showing the 68 DEGs in heart. E Subset of the GSEA results based on Molecular Signature Database Hallmarks, Reactome pathways, and GO terms gene sets in cardiac and skeletal muscles. Red and blue respectively indicate statistically significant positive and negative normalized enrichment scores (NES)
To evaluate the robustness of this common disease signature, we compared it to the list of differentially expressed genes found in XLMTM dogs [44]. Out of 132 common DEGs, 25 were also differentially expressed in the vastus lateralis or biceps femoris of the canine model of XLMTM, including Mstn, Cilp, and Tiam2 (Supplementary Dataset S13).
To investigate further the functional consequences of gene dysregulation caused by XLMTM in skeletal muscles, we performed gene set enrichment analysis (GSEA) on a list of 50 curated gene sets representing well-defined biological states and processes and obtained from the Molecular Signature Database (MSigDB). From these gene sets, we were able to identify pathways that were significantly affected in all skeletal muscles at all time points (Fig. 3E). In particular, the inflammatory response, the TNFα signaling via NFκB, and the apoptosis gene sets were positively enriched, while the fatty acid metabolism and oxidative phosphorylation gene sets had a significant negative enrichment score when comparing to WT. We then performed GSEA on 1251 gene sets obtained from the Reactome database to acquire a more refined understanding of the impacted pathways. Over 10 pathways related to mitochondrial function were depleted in at least 3 groups, which reinforced the hypothesis that oxidative phosphorylation is impaired in skeletal muscles. Additionally, several pathways related to extracellular matrix organization were enriched in the gastrocnemius, the diaphragm, and the tibialis anterior at 2 weeks (Supplementary Figure S7).
To evaluate whether these results were consistent with other transcriptomic studies in mice, independently of the genetic background and breeding/sequencing conditions, we compared them to TA and quadriceps datasets previously published [24, 25]. After removal of batch effects with the limma package, principal component analysis revealed that PC3 (11% of variance) separated the ages while genotype separation occurred with PC4 (6% of variance), regardless of the skeletal muscle or genetic background (Supplementary Figure S8A-B). GSEA revealed common dysregulated pathways in symptomatic muscles, such as oxidative phosphorylation, TNFα signaling via NFκB and epithelial mesenchymal transition, which were in agreement with the results of our study (Supplementary Figure S8C).
Overall, these results identified a disease signature common to several skeletal muscles. By considering only these 132 DEGs that were dysregulated in all muscles, we were able to confirm previous results obtained in the tibialis anterior and underline new pathways of interest.
The Mtm1 -/y mouse heart shows a mild phenotypeMyopathies can be associated to cardiac phenotypes, and some genes are implicated in both myopathy and cardiomyopathy, such as TTN and SPEG [45,46,47]. Even though skeletal muscles are severely affected in XLMTM, the cardiac function of Mtm1−/y mice has never been extensively studied. We thus performed functional phenotyping, histological and ultrastructural analyses, and gene expression quantification in Mtm1−/y mice.
First, we used noninvasive blood pressure measurement, electrocardiography (ECG), and echography to obtain a functional phenotyping of the heart at 5–6 weeks. No significant changes in heart rate and systolic blood pressure were observed in Mtm1−/y mice compared to WT (Table 2). On the ECG, a mild increase in ST segment elevation was observed in the Mtm1−/y mice, but no statistically significant differences were detected for the RR, PR, QRS, ST, and QT intervals durations, nor for the P-wave duration and R wave-amplitude (Table 1). Finally, heart echography revealed mild decreases in aortic artery diameter, stroke volume and tricuspid valve velocity time integral, but no statistically significant differences in all other measured parameters (Table 3).
Table 2 Electrocardiographic dataTable 3 Echocardiographic dataWe thus investigated further heart morphology and ultrastructure. Heart slices were stained with hematoxylin and eosin (Fig. 5A). Compared to their WT littermates, the Mtm1−/y mice did not exhibit any striking abnormalities in terms of inflammation, necrosis, fibrosis, or cardiomyocytes hypertrophy. To study subcellular structure and organization, we then conducted transmission electron microscopy. This demonstrated a normal organization of sarcomere, contractile filaments, mitochondria and membrane structures (Fig. 5B).
Overall, while these data suggested the presence of a mild cardiac phenotype without obvious structural alterations once the myopathy is well established. Compared to the skeletal muscle defects that include strong muscle weakness, muscle and myofiber hypotrophy, and disorganization of sarcomeres and organelles [48] (Supplementary Figure S4), the cardiac phenotype is very mild.
To investigate the underlying reasons for the substantial difference in severity between cardiac (minor defects) and skeletal muscle (strong impairment and disorganization), we analyzed RNA-seq data obtained from cardiac muscle samples of Mtm1−/y and WT mice. Principal component analysis showed that PC1 and PC4 (68% cumulative variance) separated the genotypes (Fig. 5C). Differential gene expression analysis revealed 68 DEGs (Fig. 5D), and GSEA revealed a 664 significantly affected pathways (Fig. 5E).
Cardiac and skeletal muscles display an opposite dysregulation pattern at the transcriptome levelStrikingly, several affected pathways in the heart were inversely affected in the skeletal muscles (Fig. 5E; Supplementary Figure S9, Supplementary Dataset S14). For example, the common skeletal muscle signature identified in this study included upregulation of pathways related to the extracellular matrix and cell adhesion, and downregulation of pathways related to mitochondrial function (oxidative phosphorylation, complex I biogenesis, respiratory electron transport, fatty acid metabolism…), and the dysregulation of these pathways was inverted in the heart.
Impaired mitochondrial function, in particular oxidative phosphorylation deficiency, is associated with an increased production of reactive oxygen species (ROS) and an increase in the level of the ROS detoxification enzyme SOD2, a superoxide dismutase located in the mitochondria [49]. Thus, we quantified ROS in skeletal and cardiac muscle with a dihydroethidium (DHE) assay (Fig. 6A), which showed elevated levels of ROS in TA and gastrocnemius but no difference in the heart. Then, we measured the levels SOD2 to evaluate whether antioxidant defenses against ROS varied in skeletal versus cardiac muscles. SOD2 was downregulated in tibialis anterior and gastrocnemius with no statistical differences in heart and diaphragm (Fig. 6B, Supplementary Figure S10A-D).
Fig. 6
Investigation of the pathways inversely dysregulated in the heart compared to the skeletal muscles. A ROS quantification with DHE assay in tibialis anterior (TA), gastrocnemius and heart. Representative fluorescent images (left) and ROS signal intensity quantification (right). Same scale for all images, scale bar 20 μm. B Protein level of SOD2 in TA, diaphragm, gastrocnemius and heart of WT and Mtm1−/y mice (n = 6) obtained by western blotting with standardization by Ponceau red staining. The fold difference from the average of the WT is shown. C Representative immunofluorescent staining images of laminin (left) and signal intensity quantification in a 2 μm-wide pericellular region (right) in tibialis anterior (TA), gastrocnemius and heart. Same scale for all images, scale bar 20 μm. D Representative immunofluorescent staining images of integrin β1 in TA and heart. Arrows indicate examples of fibers showing abnormal localization of integrin β1. Scale bars 20 μm. E Quantification of fibers with abnormal integrin β1 localization in TA and heart (n = 4). Mtm1−/y and WT littermates samples were taken at 7 weeks for all panels. Student’s t-test: ns P > 0.05, *P < 0.05, **P < 0.01, ***P < 0.001
Extracellular matrix (ECM) defects are well known in skeletal muscle affected by XLMTM, with smaller and rounder myofibers and increased inter-fiber space [3]. Therefore, we monitored laminin in cardiac and skeletal muscle with immunofluorescent assays, which revealed a thicker laminin signal around skeletal muscle fibers but not in the heart (Fig. 6C). Additionally, MTM1 has been shown to control focal adhesion and integrins recycling [3, 4]. We performed immunofluorescence staining to quantify the amount of cardiac and skeletal muscle fibers presenting abnormal β1 integrin localization. We confirmed abnormal β1 integrin internalization in tibialis anterior of Mtm1−/y mice, while no defects were detected in the heart (Fig. 6D-E).
The main substrate of MTM1 is PtdIns3P. PtdIns3P is composed of two fatty acid chains and one inositol molecule phosphorylated at position 3 of the ring [50]. In addition, PtdIns3P and MTM1 are main regulators of protein endocytosis and recycling [51], including integrins [52]. Therefore, we measured PtdIns3P levels in WT and Mtm1−/y cardiac and tibialis anterior muscles. While the level of PtdIns3P was significantly increased in Mtm1−/y tibialis anterior as expected, we observed no significant dysregulation in the heart (Fig. 7A). In addition, we assessed the level of DNM2, another protein mutated in CNM and a key regulator of membrane and protein trafficking. Western blot analysis revealed a 2.2-fold increase of DNM2 in tibialis anterior muscle and a normal level in heart in Mtm1−/y mice compared to WT mice (Fig. 7B; Supplementary Figure S10E-F).
Fig. 7
Investigation of PtdIns3P and DNM2 levels in the heart and TA. A PtdIns3P level in Mtm1−/y TA and heart as the fold difference from the average of the WT at 7 weeks. B Protein level of DNM2 in tibialis anterior (TA) and heart of WT and Mtm1−/y mice (n = 6) obtained by western blotting with standardization by Ponceau red staining in Mtm1−/y and WT mice at 7 weeks, represented as the fold difference from the average of the WT. Student’s t-test: *P < 0.05, **P < 0.01, ***P < 0.001
In conclusion, we found an inverted dysregulation of specific pathways in heart compared to skeletal muscle at the transcriptome level. We confirmed the strong alteration of these pathways in skeletal muscle at the biochemical level while they were unaffected in heart. These data, combined to the good preservation of heart function and morphology, suggest that the modulation of these pathways constitutes a molecular compensatory mechanism in heart.
留言 (0)