
記住我





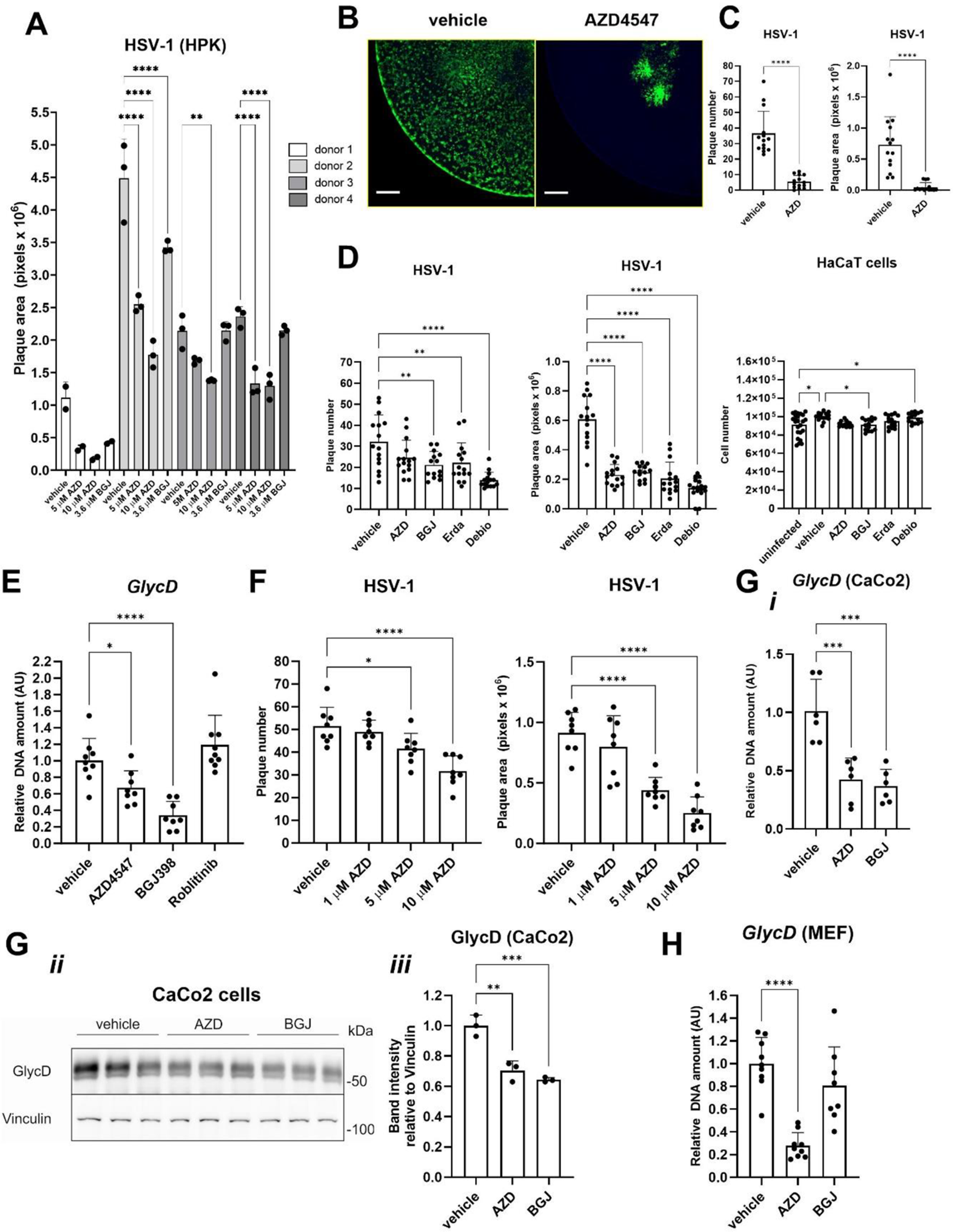
ROS have been linked to IVD degeneration [10,11,12]. To validate this association, we initially assessed the degenerative changes in the IVDs of mice at 8 weeks and 52 weeks of age. H&E staining showed no signs of degeneration in the IVDs of 8-week-old mice, whereas significant degeneration was observed in the IVDs of 52-week-old mice (Fig. 1A). Quantitative scoring of the H&E-stained sections revealed that the NP and AF tissues in 52-week-old mice had scores ranging from 3 to 5, indicating severe degeneration. In contrast, both NP and AF tissues in the 8-week-old mice received scores of 0 (Fig. 1B and C). MRI imaging of lumbar IVDs further supported these findings, showing no degeneration in 8-week-old mice, but severe degeneration in the 52-week-old group (Fig. 1D). Measurement of disc height indicated that the IVD disc height in 52-week-old mice was significantly lower than that in 8-week-old mice (Fig. 1E). Additionally, ROS levels in the lumbar IVDs of 52-week-old mice were approximately eight times higher than in the 8-week group (Fig. 1F).
Fig. 1
ROS accumulation resulted in IDD in mice. (A) Representative H&E stained sections of L1/L2 IVDs from mice aged 8 weeks and 52 weeks. Scale bars = 100 μm. (B) Quantitative scoring of the NP in degenerative IVDs shown in (A). (C) Quantitative scoring of the AF in degenerative IVDs illustrated in (A). (D) Representative MRI scans of lumbar IVDs ranging from L1 to L3 in mice aged 8 weeks and 52 weeks. (E) Measurement of disc height in L1/L2 IVDs comparing 8-week-old and 52-week-old mice. (F) Levels of ROS in lumbar IVDs of mice at 8 weeks and 52 weeks of age. (G) Schematic representation of H2O2 injection procedure near lumbar discs in mice. (H) Representative H&E staining images of lumbar IVDs (L1/L2) from mice administered with PBS and H2O2. Scale bars = 100 μm. (I) Quantified NP scores of IVDs demonstrating degeneration in mice treated as described in (H). (J) Quantified AF scores of IVDs showing degenerative changes in mice treated as indicated in (H). (K) Representative MRI scans of lumbar IVDs (from L1 to L3) in mice treated with PBS and H2O2. (L) Disc height measurements in L1/L2 IVDs from PBS and H2O2-treated mice. (M) ROS levels in lumbar IVDs of mice treated with PBS and H2O2. ***P < 0.001
To demonstrate that elevated levels of ROS can directly promote IDD, we administered subcutaneous injections of 50 µL of 2.5% H2O2 or PBS (control) near the lumbar discs of mice every five days for 12 weeks (Fig. 1G). At the end of the experiment, H&E staining revealed no degeneration in the discs of PBS-injected mice, whereas severe degeneration was observed in the discs of H2O2-injected mice (Fig. 1H). The H&E-stained sections were scored, and both NP and AF tissues in the H2O2-injected mice received scores of 3 to 5, indicating severe degeneration. In contrast, the NP and AF tissues in the PBS-injected mice scored 0 (Fig. 1I and J). MRI imaging of the lumbar IVDs further confirmed these findings, showing no signs of degeneration in the PBS group but significant degeneration in the H2O2 group (Fig. 1K). IVD disc height in H2O2 group mice was dramatically decreased than that in the PBS group (Fig. 1L). ROS levels in the lumbar IVDs of the H₂O₂ group were approximately 12 times higher than in the control group (Fig. 1M). These results suggest that the accumulation of ROS can indeed accelerate the degeneration of IVDs.
Identification of DEPs in IVDs from aged and H2O2-administrated miceTo investigate the molecular mechanisms by which ROS accumulation leads to IDD, we conducted iTRAQ protein profiling on two sets of IVDs: young vs. aged, and PBS- vs. H2O2-injected discs (n = 3 in each group) (Fig. 2A). The analysis revealed significant proteomic changes in the 52-week-old IVDs, identifying 1391 DEPs with fold changes of ≥ 1.5 and P ≤ 0.05 (Fig. 2B). Of these, 702 proteins were upregulated, while 689 were downregulated (Fig. 2B). Similarly, in the H2O2-injected mouse IVDs, 1216 proteins displayed significant alterations in expression, with 662 upregulated and 554 downregulated under the same criteria (Fig. 2C). Comparative analysis between the 52-week-old group and the H2O2 group revealed consistent upregulation of YTHDF2 and four matrix metalloproteinases (MMP1, MMP3, MMP7, and MMP9), alongside significant downregulation of the transcription factor FOXO3 and TIMP1 in both groups (Fig. 2D and Table S6). These consistent changes suggest that these proteins may play key roles in the pathogenesis of IDD induced by ROS accumulation.
Furthermore, we performed Gene Ontology (GO) and Clusters of Orthologous Groups (COG) analyses on the DEPs. In the 52-week group, the upregulated proteins were primarily involved in biological processes such as cellular response to ROS, ECM organization, MMP activity, negative regulation of cell proliferation, and transcription activation (Fig. S1). In the H2O2-treated group, the upregulated proteins predominantly participated in biological processes including cellular response to ROS, ECM organization, MMP activity, and oxidative phosphorylation (Fig. S2). Additionally, analyses of the cellular components and molecular functions of these DEPs were performed, with results presented in Fig. S1 and S2.
Fig. 2
Identification of DEPs in degenerative IVDs from aged and H2O2-injected mice. (A) Schematic of the experimental workflow for iTRAQ-based proteomic analysis conducted on degenerative IVDs from aged and H2O2-injected mice. (B and C) Volcano plots illustrating the log2 fold changes and statistical significance (− log10 p-value) of protein alterations. Red dots represent significantly upregulated proteins; green dots denote significantly downregulated proteins. (B) Comparison between 8-week-old and 52-week-old mice. (C) Comparison between PBS-treated and H2O2-injected mice. (D) Identification of proteins showing the most significant upregulation and downregulation in IVDs of 52-week-old mice and H2O2-injected mice. (E) Validation of protein expression levels for YTHDF2, FOXO3, TIMP1, MMP1, MMP3, MMP7, and MMP9 in IVDs from 8-week-old mice, 52-week-old mice, PBS-injected mice (20 weeks old), and H2O2-injected mice (20 weeks old). For each group, equal weights (0.05 g) of three independent IVDs were combined to generate homogenates, which were then analyzed via immunoblotting to assess protein levels
To validate the iTRAQ analysis results, we assessed the protein expression levels of YTHDF2, MMP1, MMP3, MMP7, MMP9, FOXO3, and TIMP1 in the lumbar IVDs of four groups of mice. The results were consistent with the iTRAQ findings. Specifically, the protein levels of YTHDF2, MMP1, MMP3, MMP7, and MMP9 was significantly elevated in both 52-week-old mice and H2O2-injected mice, while the expression of FOXO3 and TIMP1 was significantly reduced in these groups (Fig. 2E).
In vitro H2O2 treatment caused similar changes in protein levels of YTHDF2, FOXO3, TIMP1, and MMPs in NP and AF cells as observed in H2O2-injected miceTo verify that the observed changes in the expression of YTHDF2, FOXO3, TIMP1, MMP1, MMP3, MMP7, and MMP9 in aged and H2O2-injected mouse IVDs were induced by ROS, we treated mouse NP and AF cells with increasing concentrations of H2O2 (0, 50, 100, and 150 µM). Following H2O2 treatment, a rapid increase in intracellular ROS levels was observed in both NP and AF cells (Fig. 3A and B). Immunoblotting results revealed a dose-dependent increase in the protein levels of YTHDF2, MMP1, MMP3, MMP7, and MMP9 with increasing concentrations of H2O2 (Fig. 3C and D). Conversely, the expression levels of FOXO3 and TIMP1 decreased progressively with increasing H₂O₂ concentrations (Fig. 3C and D). These findings suggest that the expression of YTHDF2, FOXO3, TIMP1, MMP1, MMP3, MMP7, and MMP9 is directly influenced by ROS levels.
Fig. 3
H2O2 dose-dependently changed the expression levels of YTHDF2/FOXO3/TIMP1/MMPs in NP and AF cells. (A and B) Changes in intracellular ROS levels in NP and AF cells after treatment with varying concentrations of H2O2 (0, 50, 100, and 150 µM) for 6 h. ROS levels were quantified using DCFH-DA staining. (A) NP cells. (B) AF cells. (C and D) Protein levels of YTHDF2, FOXO3, TIMP1, MMP1, MMP3, MMP7, and MMP9 in NP (C) and AF (D) cells following H2O2 treatment. *P < 0.05, **P < 0.01
YTHDF2 was an upstream regulator of FOXO3, TIMP1, and MMP1/3/7/9Among YTHDF2, FOXO3, TIMP1, MMP1, MMP3, MMP7, and MMP9, YTHDF2 is an m6A reader that selectively recognizes and binds to the m6A-modified RR(m6A)CH consensus sequence, whereas FOXO3 functions as a transcription factor. Thus, both YTHDF2 and FOXO3 have the potential to regulate the expression of TIMP1 and MMP1, MMP3, MMP7, and MMP9. To investigate whether YTHDF2 and FOXO3 are involved in regulating the expression of TIMP1 and MMP1/3/7/9, we generated knockdown (KD) and overexpression (OE) cell lines for YTHDF2 and FOXO3 in NP cells (Fig. S3). Using these cell lines, we measured the mRNA and protein levels of YTHDF2, FOXO3, TIMP1, MMP1, MMP3, MMP7, and MMP9. Our results indicated that altering FOXO3 levels did not affect the expression of YTHDF2 but did lead to changes in the expression of TIMP1 and MMP1/3/7/9 mRNA levels (Fig. 4A). Specifically, knockdown of FOXO3 resulted in decreased TIMP1 expression and increased expression of MMP1/3/7/9 (Fig. 4A). Conversely, overexpression of FOXO3 upregulated TIMP1 expression and reduced the levels of MMP1/3/7/9 (Fig. 4A). Unlike FOXO3, modulation of YTHDF2 levels affected the expression of FOXO3, TIMP1, and MMP1/3/7/9 (Fig. 4B). Knockdown of YTHDF2 upregulated FOXO3 and TIMP1 expression, but downregulated MMP1/3/7/9 (Fig. 4B). Overexpression of YTHDF2 had the opposite effect, decreasing FOXO3 and TIMP1 levels while increasing MMP1/3/7/9 expression (Fig. 4B). Immunoblot results confirmed that protein expression patterns of YTHDF2, FOXO3, TIMP1, MMP1, MMP3, MMP7, and MMP9 were consistent with their corresponding mRNA changes in both FOXO3KD/OE and YTHDF2KD/OE cell lines (Fig. 4C and D). These findings suggest that FOXO3 functions as an upstream regulator of TIMP1 and MMP1/3/7/9, while YTHDF2 acts upstream of FOXO3.
Fig. 4
Impact of FOXO3 and YTHDF2 Knockdown and overexpression on the expression of TIMP1 and MMPs. (A) mRNA expression levels of YTHDF2, TIMP1, and MMPs in FOXO3KD and FOXO3OE cells. Quantitative PCR was performed on RNA from control knockdown (ControlKD-1 and ControlKD-2), FOXO3 knockdown (FOXO3KD-1 and FOXO3KD-2), control overexpression (ControlOE-1 and ControlOE-2), and FOXO3 overexpression (FOXO3OE-1 and FOXO3OE-2) cell lines (NP background) to assess levels of FOXO3, YTHDF2, TIMP1, MMP1, MMP3, MMP7, and MMP9. (B) mRNA expression levels of FOXO3, TIMP1, and MMPs in YTHDF2KD and YTHDF2OE cells. RNA from ControlKD-1/2, YTHDF2KD-1/2), ControlOE-1/2, and YTHDF2OE-1/2 cell lines (NP background) was analyzed to detect the expression levels of the same genes as in (A) by RT-qPCR. (C) Protein expression of YTHDF2, TIMP1, and MMPs in FOXO3KD and FOXO3OE cells. Protein samples from cells in (A) were analyzed by Western blot to detect levels of the indicated proteins. (D) Protein expression of FOXO3, TIMT1, and MMPs in YTHDF2KD and YTHDFOE cells. Protein samples from cells in (B) were subjected to Western blot analysis to detect levels of the indicated proteins. (E) The binding of FOXO3 on the promoter of TIMP1 by ChIP assay using cells in (A). (F) The binding of YTHDF2 on the promoter of TIMP1 by ChIP assay using cells in (B). **P < 0.01, ***P < 0.001
Additionally, we utilized ChIP assays to investigate whether YTHDF2 and FOXO3 could bind to the promoters of TIMP1 and MMP1/3/7/9. Our findings indicated that FOXO3 can bind to the TIMP1 promoter but not to the promoters of MMP1/3/7/9 (Fig. 4E and S4A-S4D). The binding of FOXO3 to the TIMP1 promoter was significantly reduced in FOXO3KD cells and notably increased in FOXO3OE cells (Fig. 4E). YTHDF2, however, did not show binding to the promoters of either TIMP1 or MMP1/3/7/9 (Fig. 4F and S4E-S4H). These results suggest that FOXO3 may regulate TIMP1 expression by directly binding to its promoter, and that TIMP1 subsequently influences the expression of MMP1/3/7/9. YTHDF2 appears to influence the expression of TIMP1 and MMP1/3/7/9 indirectly thorough a FOXO3-dependent mechanism.
YTHDF2 was required for FOXO3 mRNA destabilizationSome m6A readers, such as YTHDF1, YTHDF2, and YTHDF3, are involved in m6A-mediated mRNA destabilization [13,14,15]. Given our observations of consistently inverse expression patterns between YTHDF2 and FOXO3 in aged IVDs, H2O2-injected IVDs, and H2O2-treated AF/NP cells, we hypothesized that YTHDF2 might function by negatively regulating the stability of FOXO3 mRNA. To test this hypothesis, we performed RIP assays on IVDs from 52-week-old mice (n = 8), H2O2-injected IVDs (n = 8), and H2O2-treated (0 and 150 µM) AF/NP cells to assess the binding of IgG (negative control), YTHDF1, YTHDF2, YTHDF3, YTHDC1, YTHDC2, IGF2BP1, IGF2BP2, and IGFBP3 to FOXO3 mRNA. The results revealed that only YTHDF2 was able to bind to FOXO3 mRNA in both mouse IVD and AF/NP cells, whereas other m6A readers did not bind to FOXO3 mRNA (Fig. 5A-5D). Notably, the binding capacity of YTHDF2 to FOXO3 mRNA was significantly enhanced in degenerated IVDs and in H2O2-treated cells compared to controls (Fig. 5A-5D). Additionally, nine GGACU motifs were identified within the 3’-UTR (untranslated region) of FOXO3 mRNA, which were potential binding sites for YTHDF2 (Fig. 5E). Further investigations into the effects of YTHDF2 knockdown and overexpression in AF and NP cellular backgrounds revealed that YTHDF2 deficiency significantly increased FOXO3 mRNA stability, while its overexpression dramatically reduced it (Fig. 5F and I). These findings suggest that YTHDF2 plays a negative regulatory role in controlling the stability of FOXO3 mRNA.
Fig. 5
YTHDF2 was required for the stability of FOXO3 mRNA. (A) RIP assay of FOXO3 mRNA in L1/L2 IVDs from mice aged 8 weeks and 52 weeks, using antibodies against IgG, YTHDF1, YTHDF2, YTHDF3, YTHDC1, YTHDC2, IGFBP1, IGFBP2, and IGFBP3. (B) RIP assay of FOXO3 mRNA in L1/L2 IVDs from mice injected with PBS or H2O2, employing the same antibodies as in (A). (C) RIP assay of FOXO3 mRNA in NP cells treated with or without H2O2, using the same antibodies as in (A). (D) RIP assay of FOXO3 mRNA in AF cells treated with or without H2O2, utilizing antibodies as listed in (A). (E) Schematic representation of the m6A binding sites within the FOXO3 mRNA structure. (F-I) FOXO3 mRNA stability assessment. (F) FOXO3 mRNA decay in YTHDF2KD cells (NP background), measured after treatment with actinomycin D (2 µg/mL) at specified time points (0, 3, and 6 h), detected by RT-qPCR. (G) FOXO3 mRNA stability in YTHDF2KD cells (AF background) under similar conditions as in (F). (H) FOXO3 mRNA decay in YTHDF2OE cells (NP background). (I)FOXO3 mRNA stability in YTHDF2OE cells (AF background). *P < 0.05, **P < 0.01, ***P < 0.001
FOXO3 recruited histone lysine acetyltransferase CREBBP (CBP) and mediator complex subunit 1 (Med1) to regulate TIMP1 expression in the absence of ROS stimulationIn our investigation detailed in Fig. 4, we demonstrated that FOXO3 bound to the promoter region of TIMP1. Analysis of the TIMP1 promoter (2000 bp upstream of the ATG start site) identified a specific FOXO3 binding site, GTAAACA, located at positions − 1967 to -1973 (Fig. 6A). In NP cells overexpressing FOXO3 (FOXO3OE-1), Flag-tagged FOXO3 was purified through IP assay, followed by LC-MS/MS analysis, which identified 112 proteins interacting with FOXO3 (Table S7). Notably, two transcriptional regulators—CBP and Med1—were found among these interactors (Table S7). Further IP assays in NP/AF cells overexpressing FOXO3 (FOXO3OE-1) confirmed that FOXO3 could pull down both CBP and Med1 (Fig. 6B and C). To dissect the assembly of the FOXO3, CBP, and Med1 complex, we transfected NP cells with Flag- and Myc-tagged constructs in pairwise combinations. Co-IP experiments demonstrated a direct interaction between FOXO3 and CBP but not between FOXO3 and Med1 (Fig. 6D). Conversely, CBP interacted directly with both FOXO3 and Med1 (Fig. 6E), indicating that CBP served as a pivotal bridge linking FOXO3 and Med1 (Fig. 6F).
Fig. 6
FOXO3 recruited CBP and Med1 to assemble a complex in vivo and in vitro. (A) Identification of the FOXO3 binding site on the TIMP1 promoter. (B and C) FOXO3 interacted with CBP and Med1 in NP and AF cells overexpressing pCDNA3.1-Flag-FOXO3, assessed through immunoprecipitation assays using IgG-conjugated agarose A and anti-Flag resin. (B) In NP cells. (C) In AF cells. (D) Co-IP demonstrating direct interaction between FOXO3 and CBP, but not Med1. (E) Co-IP demonstrating direct interaction between CBP and FOXO3, as well as CBP and Med1. (F) Schematic diagram illustrating the assembly of the Med1-CBP-FOXO3 complex. (G) mRNA levels of TIMP1 in ControlKD, CBPKD, Med1KD, ControlOE, CBPOE, and Med1OE cells (NP background). (H) Protein levels of TIMP1 in the same cells as described in (G). (I) ChIP assays confirming the binding of the Med1-CBP-FOXO3 complex to the TIMP1 promoter, using antibodies against Med1, CBP, and FOXO3 in the same cells described in (G). **P < 0.01, ***P < 0.001
To further validate the function of this complex, both knockdown and overexpression cell lines of CBP and Med1 in NP background were generated (Fig. S5). Results indicated that depletion of either CBP or Med1 suppressed TIMP1 expression, while their overexpression led to an upregulation of TIMP1 (Fig. 6G and H). ChIP assays showed enrichment of CBP and Med1 on the TIMP1 promoter region. Disruption of any component of the FOXO3-CBP-Med1 complex decreased the enrichment of the others on the TIMP1 promoter, whereas overexpression of any complex member increased their collective enrichment (Fig. 6I).
Additionally, we examined the effects of oxidative stress by treating NP cells with H2O2. The results showed that H2O2 treatment notably reduced FOXO3 expression but did not significantly affect the expression levels of CBP or Med1 (Fig. S6A). However, the treatment significantly decreased the enrichment of the FOXO3-CBP-Med1 complex on the TIMP1 promoter (Fig. S6B). These findings elucidate the pivotal role of the FOXO3-CBP-Med1 complex in regulating TIMP1 expression in the absence of ROS stimulation and illustrate how ROS stimulation diminishes this complex’s binding to the TIMP1 promoter, leading to decreased TIMP1 expression.
Altered expression of the FOXO3-CBP-Med1 complex changed MMP1, MMP3, MMP7, and MMP9 levelsTIMP1 is an inhibitor of MMPs, and we observed an inverse expression trend between TIMP1 and MMP1/3/7/9 in degenerated IVDs and H2O2-treated cells. Given that the FOXO3-CBP-Med1 complex can regulate TIMP1 expression, we subsequently explored the effects of knocking down and overexpressing components of this complex on the expression of MMP1/3/7/9. Our findings indicated that knocking down any members of the FOXO3-CBP-Med1 complex led to upregulation of MMP1/3/7/9, while overexpressing these complex members suppressed MMP1/3/7/9 expression (Fig. S7A-S7D). Similar to FOXO3, neither CBP nor Med1 could bind directly to the promoter regions of MMP1/3/7/9 (Fig. S8A-S8D), suggesting that their regulatory effect on these MMPs was indirect.
Administration of YTHDF2 inhibitor in H2O2-treated cells upregulated TIMP1 expression but downregulated MMP1/3/7/9 expressionSince YTHDF2 was an upstream regulator of FOXO3, we hypothesized that inhibiting YTHDF2 might counteract the effects of ROS accumulation on TIMP1 and MMPs. To test this, we co-treated NP cells with the YTHDF2 inhibitor DC-Y13-27 (0, 20, and 40 µM) and H2O2 (0 and 150 µM). Following treatment, intracellular ROS levels were measured and found to be consistent with those observed with H2O2 treatment alone, indicating that DC-Y13-27 did not alter ROS levels in cells (Fig. 7A). Compared to treatment with H2O2 alone, the combined treatment with DC-Y13-27 and H2O2 consistently reduced YTHDF2 expression and resulted in increased expression of FOXO3 and TIMP1, along with decreased expression of MMP1, MMP3, MMP7, and MMP9 (Fig. 7B). RIP results demonstrated that treatment with DC-Y13-27 decreased the binding affinity of YTHDF2 to FOXO3 mRNA (Fig. 7C). Furthermore, the binding of the FOXO3-CBP-Med1 complex to the TIMP1 promoter was enhanced following treatment with DC-Y13-27 (Fig. 7D). These findings suggest that inhibition of YTHDF2 can reverse the ROS-mediated regulation of the FOXO3/TIMP1/MMP signaling axis.
Fig. 7
DC-Y13-27 diminished the effect of H2O2 on the expression of FOXO3/TIMP1/MMPs in vitro. (A) Changes in intracellular ROS levels in NP cells following treatment with H2O2 (0 and 150 µM) and DC-Y13-27 (0, 20, and 40 µM) for 6 h. ROS levels were quantified using DCFH-DA staining. (B) Protein expression levels of YTHDF2, FOXO3, Med1, CBP, TIMP1, MMP1, MMP3, MMP7, and MMP9 in NP cells after treatment with H2O2 and DC-Y13-27. (C) RIP assay to detect FOXO3 mRNA association with YTHDF2 in NP cells treated with H2O2 and DC-Y13-27 (DC), using IgG and anti-YTHDF2 antibodies. (D) Assessment of the binding dynamics of the Med1-CBP-FOXO3 complex on the TIMP1 promoter in NP cells after treatment with H2O2 and DC-Y13-27 (DC). ns represents no significant difference. *P < 0.05, **P < 0.01, ***P < 0.001
Administration of DC-Y13-27 in aged and H2O2-injected mice attenuated the progression of IDDThe effective suppression of MMPs by DC-Y13-27 in vitro under H2O2 treatment led us to evaluate its potential to inhibit the progression of IDD in mice. We initially randomized 8-week-old mice into two groups: one receiving weekly injections of PBS and the other receiving DC-Y13-27 injections (Fig. 8A). ROS levels in the IVDs of both the PBS and DC-Y13-27 groups (52-week-old mice) were significantly higher than in untreated 8-week-old mice (Fig. 8B). However, there was no notable difference in ROS levels between the PBS and DC-Y13-27 groups at the 52-week point (Fig. 8B). Mice injected with PBS exhibited severe degeneration at 52 weeks, whereas those treated with DC-Y13-27 showed only mild degeneration (Fig. 8C). Quantitative analysis of H&E staining, MRI scans, and disc height measurement confirmed that DC-Y13-27 significantly delayed IDD progression in mice (Fig. 8D and G).
Fig. 8
Administration of DC-Y13-27 delayed IDD progression in aged and H2O2-injected mice. (A) Schematic representation of DC-Y13-27 administration in aged mice. (B) ROS levels in lumbar IVDs from young mice (8 weeks old), 52-week-old mice treated with PBS or DC-Y13-27 (DC). (C) Representative H&E stained sections of L1/L2 IVDs from young mice (8 weeks old) and 52-week-old mice treated with PBS or DC-Y13-27. Scale bars = 100 μm. (D) Quantitative scoring of the NP in degenerative IVDs shown in (C). (E) Quantitative scoring of the AF in degenerative IVDs illustrated in (C). (F) Representative MRI images of lumbar IVDs (L1 to L3) from young mice (8 weeks old) and 52-week-old mice treated with PBS or DC-Y13-27. (G) Measurement of disc height in L1/L2 IVDs. (H) Schematic representation of DC-Y13-27 and H2O2 injection procedure near lumbar discs in mice. (I) ROS levels in lumbar IVDs of mice treated with PBS, H2O2, and H2O2 + DC-Y13-27 (DC). (J) Representative H&E staining images of lumbar IVDs (L1/L2) from mice administered with PBS, H2O2, and H2O2 + DC-Y13-27 (DC). Scale bars = 100 μm. (K) Quantified NP scores of IVDs demonstrating degeneration in mice treated as described in (J). (L) Quantified AF scores of IVDs showing degenerative changes in mice treated as indicated in (J). (M) Representative MRI scans of lumbar IVDs (from L1 to L3) in mice treated with PBS, H2O2, and H2O2 + DC-Y13-27 (DC). (N) Disc height measurements in L1/L2 IVDs from mice treated with PBS, H2O2, and H2O2 + DC-Y13-27 (DC). ns represents no significant difference. *P < 0.05, **P < 0.01, ***P < 0.001
To further evaluate the effects of DC-Y13-27 under oxidative stress, we randomized another set of 8-week-old mice into three groups: PBS, H2O2, and a combination of H2O2 and DC-Y13-27, with injections administered weekly for 12 weeks (Fig. 8H). Results indicated that ROS levels in the IVDs of mice treated with DC-Y13-27 or H2O2 + DC-Y13-27 were significantly higher than those in the PBS group, yet there was no notable difference between the DC-Y13-27 and H2O2 + DC-Y13-27 groups (Fig. 8I). At 20 weeks, the PBS group showed no signs of IDD, the H2O2 group exhibited severe degeneration, while the H2O2 + DC-Y13-27 group displayed only mild degeneration (Fig. 8J). Quantitative analysis of H&E staining, MRI scans, and disc height measurement also supported these findings (Fig. 8K and N). Analysis of YTHDF2, FOXO3, TIMP1, Med1, CBP, and MMPs expression in these mouse groups revealed that DC-Y13-27 treatment significantly reduced the expression of YTHDF2 and MMP1/3/7/9, increased the expression of FOXO3 and TIMP1, and did not affect the expression of Med1 and CBP (Fig. S9). These results demonstrate that DC-Y13-27 effectively suppresses the expression of YTHDF2, thereby modulating downstream signaling pathways to mitigate IDD progression.
留言 (0)