
記住我





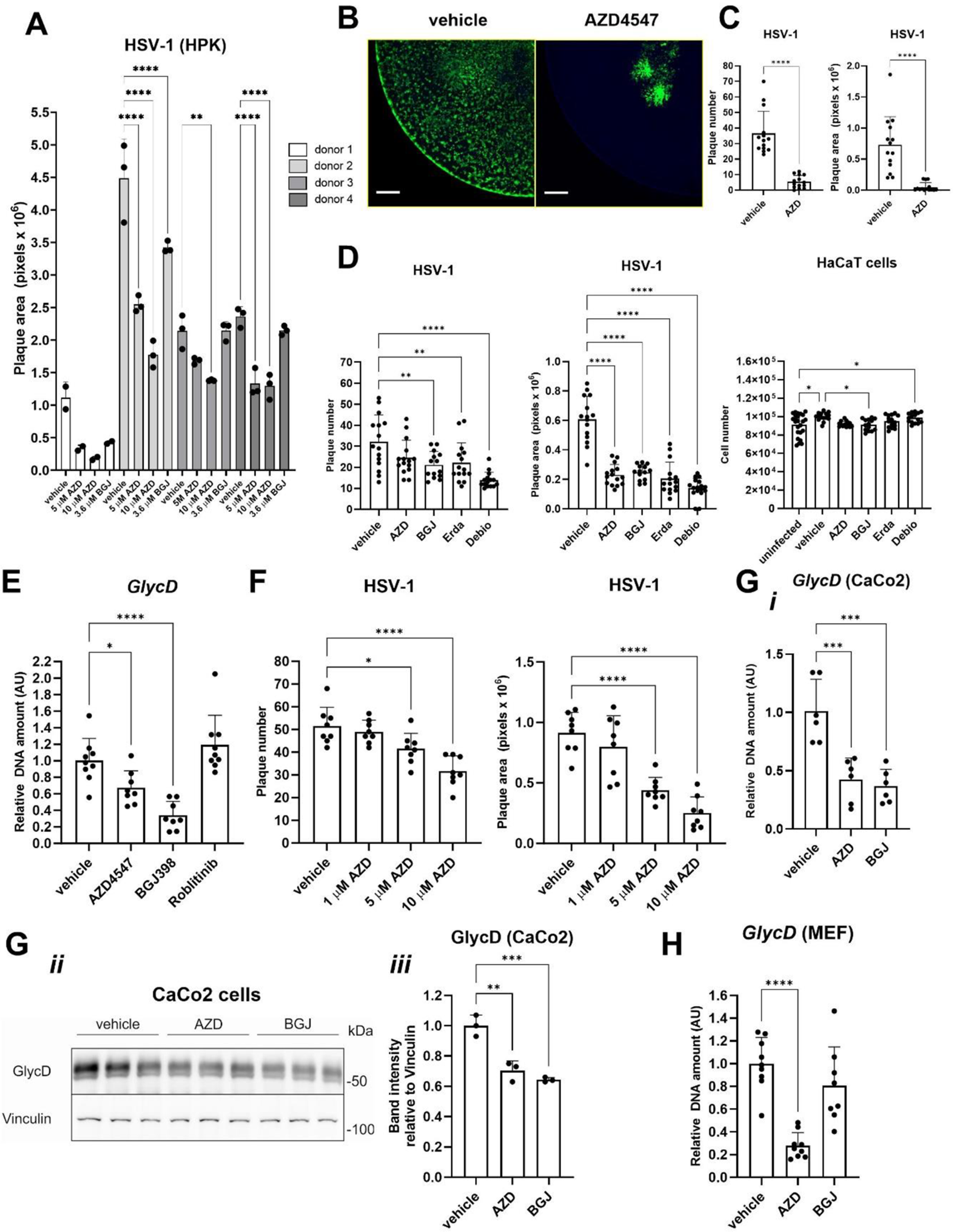
To examine the neutrophil phenotype, peripheral blood and BAL fluid samples were collected prospectively from LTx patients with CLAD, stable lung transplant recipients or LTx patients with pulmonary infection (Fig. 1A). Within the CLAD category, most patients were diagnosed with an obstructive phenotype of CLAD i.e. bronchiolitis obliterans syndrome (BOS). Only 1/8 of the BAL fluid and 1/6 of the peripheral blood samples were derived from patients with a restrictive CLAD phenotype i.e. restrictive allograft syndrome (RAS). No significant differences were observed in age, sex and underlying disease and almost no acute rejection or lymphocytic bronchiolitis was observed on biopsies taken at the time of BAL fluid or blood sampling (Table 1). Medication use was also not significantly different across the patient groups, except for treatment with montelukast for CLAD patients. Patients with CLAD were also sampled at later post-operative time points compared to infected or stable LTx patients, which is to be expected given the increased morbidity of CLAD at higher post-operative days (PODs) [1]. Total and differential blood leukocyte counts were comparable for all patient groups (Table 1; Fig. 2). In the BAL fluids, no significant differences were noted for BAL fluid return, total cells, lymphocyte and eosinophil counts. Macrophages were significantly decreased in percentage, but not in absolute numbers, in CLAD and infected relative to stable LTx patients. Finally, the percentage and absolute counts of neutrophils in the BAL fluids of CLAD and infected LTx patients were significantly increased compared to stable LTx recipients (Table 1; Fig. 2).
Fig. 2
BAL fluid and blood neutrophil counts from LTx patients included for phenotypical neutrophil analysis. BAL fluid and peripheral blood samples were collected from CLAD (n = 8 BAL fluids & 6 blood samples), infected (n = 16 BAL fluids & 15 blood samples) and stable LTx patients (n = 9 BAL fluids & 21 blood samples). Data are shown as box-and-whisker plots (box: median with interquartile range, whiskers: full data distribution), with each dot representing an individual patient sample, and statistically analyzed by Kruskal-Wallis tests with Dunn’s multiple comparisons tests for comparisons between BAL fluids or blood samples of different patient categories: * p < 0.05; ** p < 0.01; *** p < 0.001
Heterogeneity in neutrophil maturity in the circulation and lungs of LTx patientsBAL fluid cells and purified peripheral blood neutrophils were subjected to multicolor flow cytometry and microscopy analysis to examine neutrophil maturity. Compared to cells from healthy donors, a (significantly) increased yet highly variable number of immature neutrophils – featured by a lack of membrane expression of CD10 [23] – was detected in both BAL fluids and blood of CLAD, infectious and stable LTx patients (Fig. 3A). No significant differences were observed between the different patient groups. Lower expression levels of the low-affinity Fcγ receptor, i.e. FcγRIII (CD16), and the adhesion molecule L-selectin (CD62L) were detected on the blood neutrophils of the LTx patients compared to healthy controls (Fig. 3B, C & Supplementary Fig. 4A). In conjunction with CD10, these two markers can be exploited to distinguish between mature (3–4 nuclear lobes) neutrophils (CD10+CD16highCD62Lhigh), hypersegmented (> 4 nuclear lobes) neutrophils (CD10+CD16highCD62Ldim) and immature neutrophils (CD10−CD16dimCD62Lhigh) [24, 25]. Confirming the data regarding CD10 expression levels, a minor but significantly increased percentage of immature neutrophils was observed in the blood of stable and infected LTx patients compared to healthy controls, showing fully mature circulating neutrophils (Fig. 3D, E). Interestingly, a significantly increased fraction of the mature neutrophils in LTx patients were characterized as hypersegmented cells, which has been associated with a hypermature, aged neutrophil phenotype (Fig. 3F) [26]. The marked variability in neutrophil maturity between different patients, regardless of the disease group, was also microscopically visible. Some LTx patient blood samples exhibited high numbers of immature neutrophils with a banded nucleus and even early neutrophil precursors, i.e. metamyelocytes. Hypermature, hypersegmented neutrophils were observed in other samples, correlating to the expression levels of CD10, CD62L and CD16 (Fig. 3G-I). Despite this considerable inter-patient variability in neutrophil maturity, no direct correlation could be established between the maturation state in the blood and the type or dosage of medication nor disease severity.
Fig. 3
Phenotypical analysis of the maturation of BAL fluid and blood neutrophils from LTx patients. BAL fluid and peripheral blood samples were collected from CLAD (n = 8 BAL fluids & 6 blood samples), infected (n = 16 BAL fluids & 15 blood samples) and stable LTx patients (n = 9 BAL fluids & 21 blood samples), as well as healthy controls (HC, n = 17 blood samples). (A-F) Flow cytometry was used to evaluate the membrane expression of (A) CD10, (B) CD16 and (C) CD62L on BAL fluid and blood neutrophils (gated as CD16+CD66b+ cells). (D-F) Based on CD16 and CD62L expression, blood neutrophils were characterized as immature, mature or hypersegmented cells. (G-I) Representative microscopy images of blood neutrophils from a (G) healthy volunteer or from a LTx patient with (H) immature or (I) hypersegmented neutrophils, as indicated by arrows. The neutrophil expression levels of CD10, CD16 and CD62L of these individuals are indicated below the pictures. (J-L) Membrane expression of (J) CXCR4 or (K,L) CXCR4 in combination with CD10 to classify CXCR4+ BAL fluid neutrophils as immature or aged. Results represent percentages of positive neutrophils or median fluorescence intensity (MFI). Data are shown as box-and-whisker plots (box: median with interquartile range, whiskers: full data distribution), with each dot representing an individual patient sample, and statistically analyzed by Kruskal-Wallis tests with Dunn’s multiple comparisons tests for comparisons between BAL fluids or blood samples of different patient categories: * p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001. Mann-Whitney U tests were performed for BAL fluid-blood comparisons per patient category: $$ p < 0.01; $$$ p < 0.001; p < 0.0001 vs. blood. Some BAL fluid samples could not be included in the flow cytometry analysis for every marker, due to shortage of neutrophils
The reduced CD10 expression levels also suggested the presence of some immature neutrophils in the BAL fluids of the LTx patients (Fig. 3A). The chemokine receptor CXCR4 is typically present on immature neutrophils in the bone marrow [27], but can also be upregulated on aged neutrophils [28]. In the LTx patient BAL fluids, up to 21% of neutrophils expressed CXCR4 (Fig. 3J). Since CD62L expression is very low or absent on BAL fluid neutrophils (Fig. 3C & Supplementary Fig. 4A), CXCR4 was used in conjunction with CD10 to distinguish immature (CXCR4+CD10−) from aged (CXCR4+CD10+) neutrophils. Both subsets were found in the BAL fluids of the LTx patients (Fig. 3K, L). A significant negative correlation was found between the administered dose of tacrolimus and the percentage of CD10+ mature neutrophils in the BAL fluid (Supplementary Fig. 5A). However, this correlation was not observed with the numbers of mature peripheral blood neutrophils. Furthermore, no correlations were found with other immunosuppressive drugs or disease severity. In conclusion, our findings reveal a high level of heterogeneity in neutrophil maturation in the blood and BAL fluids of LTx patients, regardless of type or severity of disease, ranging from immature to aged, hypersegmented neutrophils.
Uniform activation phenotype of neutrophils in the lungs of stable, CLAD and infected LTx patientsUsing flow cytometry, we also investigated the presence of activation markers and chemoattractant receptors on neutrophils. Expression levels of the neutrophil degranulation markers CD66b and CD11b and the integrin chain CD11c were significantly increased on BAL fluid in comparison to blood neutrophils of LTx patients with the exception of CD11b in stable LTx recipients (Fig. 4A-C). The selectin ligand CD15 (Sialyl-LewisX) only exhibited a trend to be upregulated on neutrophils in BAL fluids of LTx patients with infection (Fig. 4D). Instead, CD15 expression was significantly increased on blood neutrophils from LTx patients with infection or CLAD compared to stable LTx recipients, but no significant differences with healthy controls were observed. Neutrophil chemoattractant receptors CXCR1, CXCR2 and complement receptor C5aR were all significantly downregulated on BAL fluid compared to blood neutrophils in the different disease categories of LTx patients (Fig. 4E-G & Supplementary Fig. 4B-D). Higher administered doses of oral corticosteroids were associated with higher remaining expression levels of CXCR2 and C5aR on BAL fluid neutrophils (Supplementary Fig. 5B, C), but not on peripheral blood neutrophils. Corticosteroids or other administered drugs also did not correlate with changes in expression of degranulation markers on the neutrophils. The increased degranulation and upregulation of integrins, along with the internalization of chemoattractant receptors upon ligand-induced stimulation, suggest neutrophil activation in the lungs of all LTx patients [8, 29, 30]. However, no significant differences were observed between the different patient groups, indicating a uniform neutrophil activation phenotype in the lungs. The partial downregulation of CXCR2 on blood neutrophils of all LTx patients and the significantly increased CD66b expression on blood neutrophils of LTx patients with infection, compared to healthy controls, may indicate that patient neutrophils already become partially activated in circulation (Fig. 4A, F). However, a partial downregulation of CXCR2 may also be a sign of neutrophil aging [28], and unlike in COVID-19 patients, no internalization of the complement receptor C5aR was observed on circulating neutrophils of LTx patients [31]. Furthermore, the formyl peptide receptors FPR1 and FPR2 and, to a minor extent, CCR1 (but not CCR2) – which both are normally not expressed by neutrophils - were upregulated on BAL fluid neutrophils (Fig. 4H-J & Supplementary Fig. 4E). Minor upregulation of CCR1 on neutrophils in BAL fluid may be explained by the presence of IFN-γ which has been reported to upregulate CCR1 on neutrophils [32] and was detected in BAL fluids from lung transplant patients in a previous report [17]. Conversely, the decoy interleukin receptor IL1-R2 was significantly downregulated on BAL fluid neutrophils (Fig. 4K). Additionally, a small population of neutrophils expressed the antigen-presenting molecule HLA-DR (Fig. 4L), similar to what was observed on neutrophils from COVID-19 patients [21, 33]. No expression of the chemokine receptor CXCR3 and the active IL-1 receptor, i.e. IL1-R1, was detected on the LTx patient neutrophils. For some patients, paired BAL fluid and peripheral blood samples could be obtained. Flow cytometry analysis on these samples led to similar observations (Supplementary Fig. 6). Finally, in the BAL fluids of the LTx patients, we measured concentrations of neutrophil elastase, an important protease released during neutrophil degranulation (activation). Interestingly, neutrophil elastase levels were significantly increased in CLAD and infected compared to stable LTx patients (Fig. 4M) and correlated to the number of BAL fluid neutrophils (Fig. 4N, O). In conclusion, neutrophils in the BAL fluids of LTx patients exhibit a uniform, activated phenotype regardless of the disease state (stable, CLAD or infected) or immunosuppressive treatment of the patient. The augmented inflammation in LTx patients with CLAD or infection compared to stable LTx recipients may therefore be mostly related to increased neutrophil recruitment to the lungs, rather than an enhanced activation of individual cells.
Fig. 4
Phenotypical analysis of activation markers and receptors on BAL fluid and blood neutrophils from LTx patients. BAL fluid and peripheral blood samples were collected from CLAD (n = 8 BAL fluids & 6 blood samples), infected (n = 16 BAL fluids & 15 blood samples) and stable LTx patients (n = 9 BAL fluids & 21 blood samples), as well as healthy controls (HC, n = 17 blood samples). Flow cytometry was used to evaluate the membrane expression of (A) CD66b, (B) CD11b, (C) CD11c, (D) CD15, (E) CXCR1, (F) CXCR2, (G) C5aR, (H) FPR1, (I) FPR2, (J) CCR1, (K) IL-1R2 and (L) HLA-DR on BAL fluid and blood neutrophils (gated as CD16+CD66b+ cells). Results represent percentages of positive neutrophils or median fluorescence intensity (MFI). (M) Neutrophil elastase levels in the BAL fluid supernatants were determined by ELISA. (N, O) Correlation between neutrophil elastase concentrations and the percentage and absolute numbers of neutrophils in the LTx BAL fluid samples (CLAD, infection and stable combined). Data are shown as box-and-whisker plots (box: median with interquartile range, whiskers: full data distribution), with each dot representing an individual patient sample, and statistically analyzed by Kruskal-Wallis tests with Dunn’s multiple comparisons tests for comparisons between BAL fluids or blood samples of different patient categories: * p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001. Mann-Whitney U tests were performed for BAL fluid-blood comparisons per patient category: $ p < 0.05; $$ p < 0.01; $$$ p < 0.001; p < 0.0001 vs. blood. Correlation analysis was performed calculating a Spearman correlation coefficient and plotted utilizing a simple linear regression line. Some BAL fluid samples could not be included in the flow cytometry analysis for every marker, due to shortage of neutrophils
Patient characteristics for the CXCL8 proteoform cohortTo provide insight into the putative mechanisms underlying neutrophil accumulation in the lungs of the LTx patients, we performed differential quantification of CXCL8 proteoforms known to possess up to 30-fold differences in activity. CXCL8 proteolysis was assessed in BAL fluid supernatants from a second cohort of CLAD, infected and stable LTx patients, as well as in COVID-19 and influenza patients (Fig. 1B). No significant differences were observed in age, sex, underlying disease, POD of sampling or medication use between the different patient groups (Table 2). However, total cell counts and both neutrophil percentages and absolute counts were significantly increased in the BAL fluids of LTx patients with CLAD or infection compared to stable lung transplant recipients (Fig. 5A, B). It is important to note that for CLAD and infected LTx patients, neutrophil-rich BAL fluid samples (> 15% neutrophils) were specifically selected, to be able to determine endogenous CXCL8 proteoforms. Detailed clinical data for the LTx patients can be found in Table 2. Clinical data of the COVID-19 and influenza patients are described in Supplementary Table 1.
Fig. 5
Detection of endogenous CXCL8 proteoforms in BAL fluid. (A-C) Neutrophil counts and CXCL8 levels in BAL fluid samples from CLAD (n = 12), infected (n = 6) and stable LTx patients (n = 6) used for CXCL8 proteoform characterization. (D,E) Endogenous CXCL8 proteoforms were determined by ISTAMPA in the BAL fluid supernatants of LTx patients with CLAD or infection. Total CXCL8 levels were below the detection limit for ISTAMPA analysis in stable LTx recipients. CXCL8 proteoforms were presented according to their relative abundance in the BAL fluids, expressed as percentage of total CXCL8. (F,G) Correlation between the relative abundance of the most potent proteoform CXCL8(9–77) and the percentage and absolute numbers of neutrophils in the LTx BAL fluid samples (CLAD and infection combined). (H,I) ISTAMPA analysis of endogenous CXCL8 proteoforms in COVID-19 (n = 24) and influenza (n = 7) BAL fluid supernatants. Data are shown as box-and-whisker plots (box: median with interquartile range, whiskers: full data distribution), with each dot representing an individual BAL fluid sample, and statistically analyzed by Kruskal-Wallis tests with Dunn’s multiple comparisons tests: * p < 0.05; ** p < 0.01; *** p < 0.001. Correlation analysis was performed calculating a Spearman correlation coefficient and plotted utilizing a simple linear regression line
Detection of endogenous CXCL8 proteoforms in the lungs of LTx, COVID-19 and influenza patientsSimilar to the increased neutrophil numbers, total CXCL8 levels were significantly elevated in the BAL fluids of LTx patients with CLAD and even more upon infection in comparison to stable LTx recipients (Fig. 5C). To characterize the specific CXCL8 proteoforms in these BAL fluid samples, we utilized the ISTAMPA method, which combines immunosorbent pre-purification of total CXCL8 (i.e. all CXCL8 proteoforms) with sensitive nano-LC-MS/MS to identify individual natural CXCL8 proteoforms [16]. In the BAL fluids of both individuals with CLAD and LTx patients with infection, CXCL8 was NH2-terminally truncated, with intact CXCL8(1–77) representing only a median of 5–20% of the total CXCL8 content (Fig. 5D, E). Apart from the full-length protein, the NH2-terminally truncated proteoforms CXCL8(6–77), CXCL8(8–77) and CXCL8(9–77) - known to exhibit 10 to 30-fold superior biological activity - were most abundant. No significant differences in CXCL8 proteoforms were found between the LTx patients with CLAD or infection. Importantly, a significant positive correlation was observed between the most active proteoform, i.e. CXCL8(9–77), and both the percentage as well as absolute counts of neutrophils in the BAL fluids of the LTx patients (Fig. 5F, G). This correlation was not observed for other CXCL8 proteoforms. Interestingly, in some BAL fluid samples, CXCL8(10–77) was detected, a proteoform never identified before in biological samples. Although presumed to be inactive due to proteolytic cleavage within the ELR motif [34], it was reported to exhibit some remaining receptor affinity and in vitro chemotactic activity, while its capacity for inducing neutrophil elastase release is strongly impaired [13]. Finally, in the BAL fluids of COVID-19 and influenza patients (Fig. 5H, I) and similar to LTx patients, CXCL8 was also mostly cleaved towards its more active proteoforms CXCL8(6–77), CXCL8(8–77) and CXCL8(9–77) and the presumably inactive proteoform CXCL8(10–77), suggesting CXCL8 proteolysis may be important during both chronic and acute neutrophilic inflammation. An overview of the top-down MS/MS spectra of detected CXCL8 proteoforms in the BAL fluids is provided in Supplementary Fig. 7. Total levels of CXCL8 are shown in Supplementary Fig. 8. In conclusion, our study demonstrates that CXCL8 consists of highly potent truncated CXCL8 proteoforms in LTx patients with CLAD and infection, as well as in patients with COVID-19 or influenza. The most potent proteoform CXCL8(9–77) thereby specifically correlated with the neutrophil counts in the LTx BAL fluids. An amplification loop, wherein (neutrophil-derived) proteases process CXCL8 into more active proteoforms to increase neutrophil chemotaxis and activation, therefore likely contributes to the excessive neutrophilic inflammation in these diseases.
Proteolytic processing of CXCL8 in BAL fluids & inhibition of proteolysisMany proteases have been reported to cleave the NH2-terminus of CXCL8 in vitro [15, 35,36,37,38,39,40]. To further understand how these CXCL8 proteoforms are generated in patients, intact CXCL8(1–77) was spiked into a small volume of LTx BAL fluid samples. After 3 h of incubation at 37 °C, allowing proteases present in the BAL fluids to process CXCL8, proteoforms were analyzed by ISTAMPA. In the LTx patients with CLAD or infection, proteolytic processing predominantly yielded CXCL8(6–77), in addition to minor amounts of the most potent proteoforms CXCL8(8–77) and CXCL8(9–77). In BAL fluids of stable LTx recipients, where endogenous CXCL8 concentrations were below the detection limit of ISTAMPA, processing of intact CXCL8(1–77) occurred significantly slower, and CXCL8(8–77) and CXCL8(9–77) were not generated (Fig. 6A). Notably, CXCL8(10–77) – which was highly abundant in the BAL fluid samples (Fig. 5) – could not be generated from intact CXCL8(1–77) within 3 h. In the COVID-19 and influenza patients, comparable proteolytic processing was observed although it tended to be slower in comparison to the CLAD and infected LTx patients (Fig. 6A). In COVID-19 BAL fluids, we noticed that shorter or longer (up to 16 h) incubation times only resulted in, respectively, decreased or increased processing towards more potent CXCL8(6–77) and the shorter CXCL8(8–77) and CXCL8(9–77) proteoforms but not to the less potent CXCL8(10–77) form (Fig. 6B). Furthermore, enhanced proteolytic (elastinolytic) activity in the COVID-19 BAL fluid samples [21] accelerated the processing speed of CXCL8(1–77) towards CXCL8(6–77) (Fig. 6C, D).
Fig. 6
Proteolytic processing of intact CXCL8 in BAL fluid. (A) Recombinant human CXCL8(1–77) [125 ng] was spiked in 10 µL of BAL fluid supernatant from CLAD (n = 12), infected (n = 6) or stable LTx patients (n = 6) and from patients with COVID-19 (n = 10) or influenza (n = 7). After incubation for 3 h at 37 °C, CXCL8 proteolysis was analyzed by ISTAMPA. (B) CXCL8 proteoforms were also determined immediately after spiking CXCL8 [without 37 °C incubation but including the required 30 min pre-purification of the CXCL8 proteoforms at room temperature (RT) as part of the ISTAMPA procedure] and after 16 h of incubation at 37 °C in the COVID-19 BAL fluids. CXCL8 proteoforms were presented according to their relative abundance in the BAL fluids, expressed as percentage of total CXCL8. (C,D) Correlation between the relative abundance of the intact CXCL8(1–77) or the truncated CXCL8(6–77) proteoform after 3 h incubation at 37 °C and the elastinolytic activity in the COVID-19 BAL fluid samples [21]. Data are shown as box-and-whisker plots (box: median with interquartile range, whiskers: full data distribution), with each dot representing an individual BAL fluid sample, and statistically analyzed by Kruskal-Wallis tests with Dunn’s multiple comparisons tests: * p < 0.05; **** p < 0.0001. Correlation analysis was performed calculating a Spearman correlation coefficient and plotted utilizing a simple linear regression line
CXCL8(8–77) and CXCL8(9–77) may, at least partially, be products of further processing from the CXCL8(6–77) proteoform. To investigate this, CXCL8(6–77) was incubated in BAL fluid samples from both COVID-19 and LTx patients for 3 h at 37 °C and indeed, proteolysis towards CXCL8(7–77), CXCL8(8–77) and CXCL8(9–77) was observed. This processing was mediated by metalloproteases, as it could be inhibited by EDTA (Fig. 7A). Finally, as a proof-of-concept experiment, we aimed to block the initial processing of intact CXCL8(1–77) towards CXCL8(6–77) and the shorter proteoforms, as this has potential to reduce CXCL8 activity and consequent neutrophil chemotaxis and activation. Inhibition of serine proteases or metalloproteases with AEBSF or EDTA, respectively, could partially prevent NH2-terminal proteolysis (Fig. 7B). However, when both inhibitors were used together, near-complete inhibition of processing was achieved, suggesting that a combination of serine proteases and metalloproteases are involved in the processing of intact CXCL8 in the patient lungs. In conclusion, our findings show rapid proteolytic activation of CXCL8 in BAL fluids from LTx patients with CLAD or infection. Stable LTx recipients not only have very low total CXCL8 levels in their lungs, but also exhibit slower CXCL8 processing, which may explain their reduced airway neutrophilia. Targeting endogenous proteases in the lungs could be a promising therapeutic strategy to reduce CXCL8 processing, thus limiting excessive neutrophil infiltration, activation and inflammation, and should be further investigated in vivo.
Fig. 7
Inhibition of proteolytic processing of CXCL8 in BAL fluid. (A) Recombinant human CXCL8(6–77) [125 ng] was spiked in 10 µL of BAL fluid supernatant (n = 3–6) from COVID-19 (indicated by dots) and CLAD patients (indicated by triangles) and incubated for 3 h at 37 °C, in the absence or presence of the metalloprotease inhibitor EDTA. (B) Recombinant human CXCL8(1–77) [125 ng] was spiked in 10 µL of BAL fluid supernatant (n = 8–9) from COVID-19 (dots) and CLAD patients (triangles) and incubated for 3 h at 37 °C, in the absence or presence of the serine protease inhibitor AEBSF or/and the metalloprotease inhibitor EDTA. After incubation, CXCL8 proteolysis was determined by ISTAMPA. All CXCL8 proteoforms were presented according to their relative abundance in the sample, expressed as percentage of total CXCL8. Data are shown as box-and-whisker plots (box: median with interquartile range, whiskers: full data distribution), with each dot representing an individual BAL fluid sample, and statistically analyzed by Mann-Whitney U tests or Kruskal-Wallis tests with Dunn’s multiple comparisons tests: * p < 0.05; *** p < 0.001; **** p < 0.0001
留言 (0)