
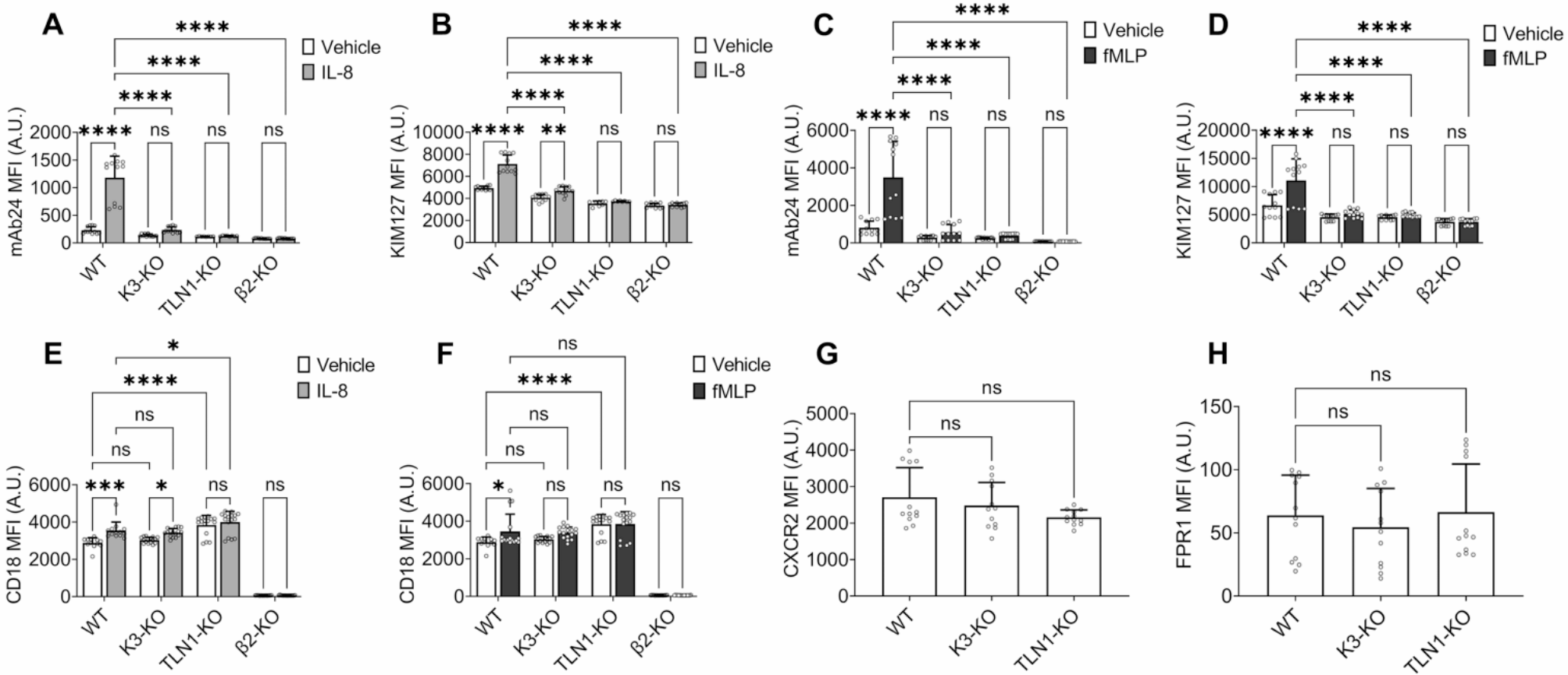
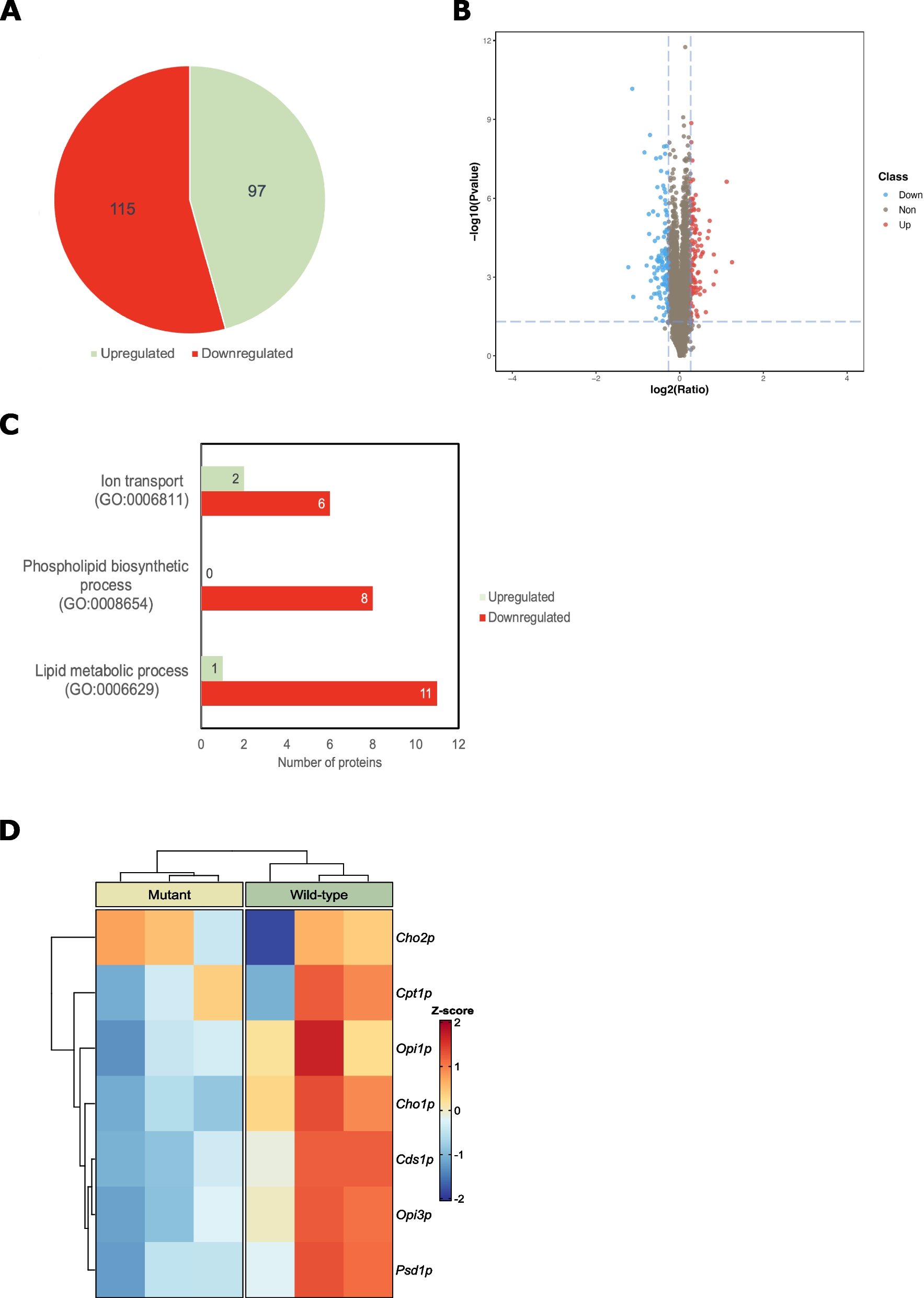
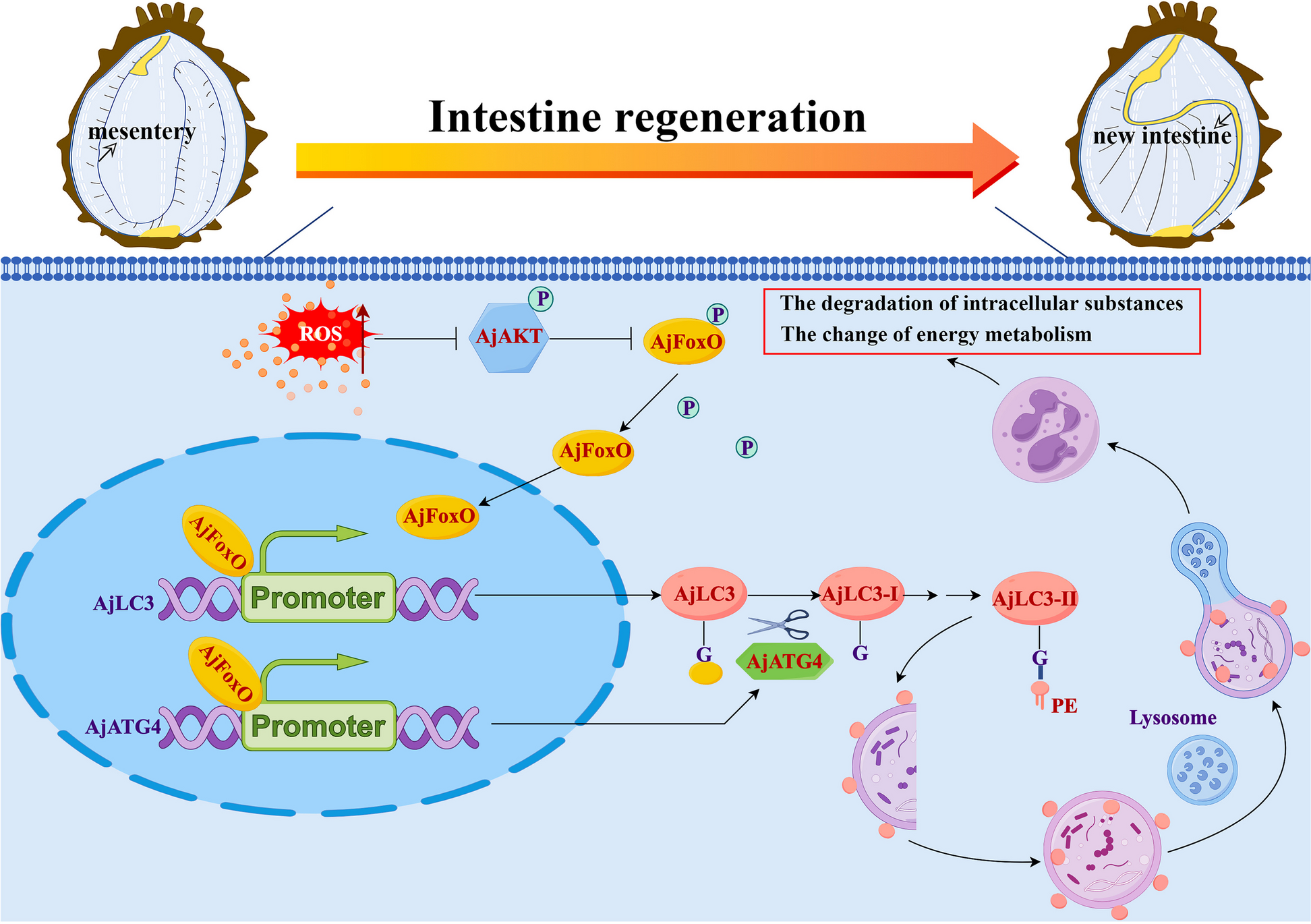
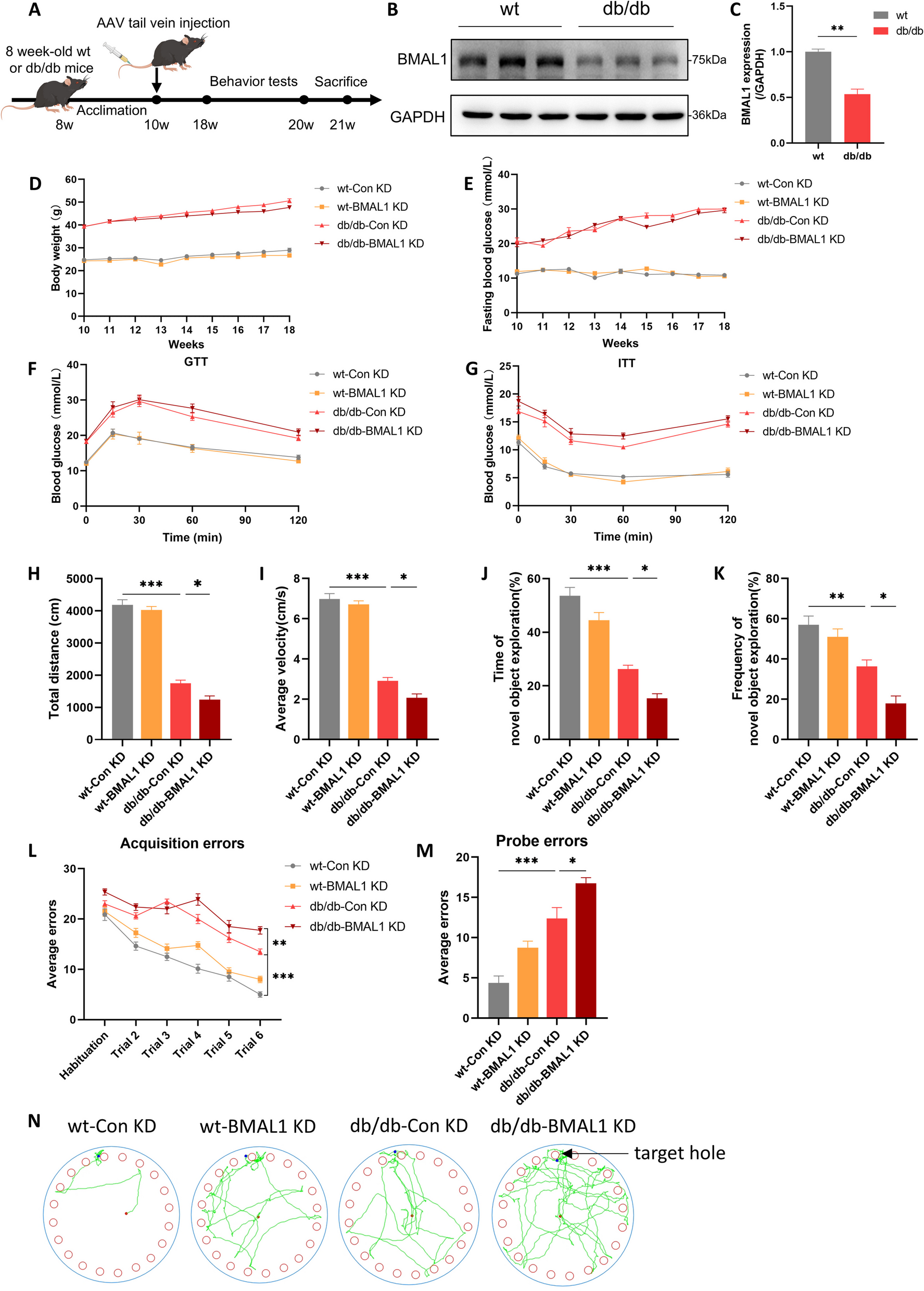
記住我




To identify miRNAs that are reduced upon ER stress in both cancer and noncancer human airway epithelial cells (Calu 3 and 16HBE14o-, respectively), we compared miRNA expression profiles obtained with two independent genome-wide approaches, genome-wide miRNA expression arrays (for Calu 3 cells) [15] and next generation sequencing (for 16HBE14o- cells) [18]. Both of these approaches were based on two classic ER stress models, a proteasome inhibitor, ALLN (calpain inhibitor I) and a glycosylation inhibitor, tunicamycin, to activate the UPR. The activation of both UPR adaptive and apoptotic signaling was verified by parallel genome-wide mRNA expression arrays (for Calu 3 cells) [15] and next generation sequencing (for 16HBE14o- cells) [6]. The increase in heat shock protein family A (Hsp70) member 5 (HSPA5, also known as BIP) mRNA levels was accompanied by a similar elevation of CHOP (DDIT3) mRNA levels in both cell lines and in both ER stress models, confirming that the UPR was fully activated (Supplemental Fig. 1) We selected miRNAs that were decreased by both ER stressors in both Calu 3 and 16HBE14o- cells (Fig. 1AB). The only miRNA found under all these conditions was miR-1244 (Fig. 1AB). Next, we verified that ER-stress related downregulation of miR-1244 expression was independent of the pharmacological ER stressor as well as that this miRNA was also reduced in other human cells such as HeLa S3. As shown in Fig. 1C, hsa-miR-1244 was reduced independently of the mechanism of UPR induction not only in Calu 3 and 16HBE14o- cells, but also in HeLa cells. Furthermore, miR-1244 levels remained unaffected when the cells were treated with the cytotoxic agent C-1305 (Supplemental Fig. 2). Taken together, this data suggests that the downregulation of miR-1244 during ER stress is representative and potentially important for the UPR in human cells. However, the magnitude of hsa-miR-1244 reduction in response to ER stress varied among cell types and ER stressors and this may reflect differences in basal expression levels of hsa-miR-1244.
Fig. 1
UPR is accompanied by reduced levels of miR-1244. (A). Volcano plots illustrate the ER stress related changes in miRNA expression profiles in Calu 3 cells exposed to Tm (2.5 µg/ml) (left) and ALLN (100µM) (right). The miRNAs that were changed by 2 log2 fold (P value < 0.05) were selected, and downregulated ones are marked with blue and upregulated ones are marked with red. (B). Volcano plots are shown that represent ER stress-related changes in miRNA expression profiles in 16HBE14o- exposed to Tm (left) and ALLN (right). The miRNAs that were (P value < 0.05) changed by 2 log2 fold (P value < 0.05) were selected, and downregulated ones are marked with blue whereas upregulated ones are marked with red. (C) Relative miR-1244 expression in different cell types following UPR activation with various ER stressors. Calu-3, 16HBE14o-, and HeLa cells were treated with different ER stressors (Tm, 2.5 µg/ml Tg – thapsigargin, 50 nM, ALLN 100µM, dithiothreitol – DTT 4 mM, and brefeldin A – BA, 0.4 µg/ml) for 8 h and total RNA enriched in miRNAs was isolated. mir-1244 levels were quantified by qPCR and normalized to RNU48. Results are plotted as fold changes over control miR-1244 levels in untreated cells. Data represents the mean ± SD of three independent experiments. *P < 0.05, **P < 0.001, ***P < 0.0001 were considered significant
Thus, to define the capacity of miR-1244 regulatory activity, we performed an NGS based genome-wide analysis of mRNA expression profiles in HeLa cells after miR-1244 overexpression with its synthetic analog (hsa-miR-1244 mimic) and more importantly, after inhibition of endogenous miR-1244 with a specific inhibitor (hsa-miR-1244 antagomiR). Here we applied a miR-1244 target selection analysis that would allow us to focus on this molecule’s role in UPR signaling. Given that miRNA effects on gene expression serve a modulatory function and result from simultaneous interactions of many different miRNA with specific targets [29], we set our selection threshold to 1.5 log2 fold change. Furthermore, since ER stress reduced miR-1244 levels, we decided to include in our analysis transcripts that were identified to be upregulated during ER stress in our previous genome wide analysis in 16HBE14o- cells [6]. As shown in Fig. 2A, this approach resulted in identification of a large group of potential direct miR-1244 target mRNAs (107 mRNAs) (Supplemental Data Set D) that were upregulated by ER stress and upregulated by inhibitor or downregulated by mimic. The functional aspects of these gene changes were strongly and significantly related to UPR that included IRE1 signaling, protein processing in ER, and interleukin-regulated apoptosis (Fig. 2B, Supplemental Data Set E). Notably, the potential miR-1244 targets included crucial receptors and mediators of UPR including XBP1, ATF6, IRE1 (ERN1) and PERK as well as genes implicated in ER stress related apoptosis such as NOXA, PUMA (BBC3) or XIAP (Fig. 2C).
Fig. 2
miR-1244 targets are involved in UPR. (A) Venn diagram summarizing the strategy used to select ER stress related miR-1244 targets. The analysis included mRNAs that were either downregulated by miR-1244 mimic or upregulated by its inhibitor (by 1.5 log2 fold). ER stress upregulated genes in 16HBE14o- cells (by 1.5 log2 fold) and the genes that were predicted with MIRDIP to contain miR-1244 binding sites are shown (with P value < 0.05). (B) The summary of the gene ontology analysis of selected miR-1244 targets − (104 mRNAs) that were upregulated by ER stress and upregulated by inhibitor or downregulated by mimic are shown (Supplemental Data Set). (C) The hierarchical clustering analysis (Pearson correlation based) of qPCR analyzed changes in expression of miR-1244 UPR related targets in HeLa cells exposed to Tm or Tg for 16 h and transfected with scramble control, miR-1244 mimic or antagomiR are shown. The values represent the mean of 3 independent experiments. (D) HeLa cells were treated with Tm (2.5 µg/ml) or Tg (500nM) for the times indicated, and total RNA enriched in miRNAs was isolated. MiR-1244 levels were quantified by qPCR and normalized to RNU48. Data represents the mean ± SD of three independent experiments. *P < 0.05 was considered significant
Next, to verify these changes in the UPR, our follow up studies were based on the independent validation (qPCR) of these potential miR-1244 targets, as well as the related changes in BIP and CHOP expression in HeLa cells exposed to ER stress and transfected with either miR-1244 mimic or inhibitor. All these genes were tested in no stress conditions as well. Since ALLN is known to partially inhibit IRE1 activity, as a secondary ER stress model we used thapsigargin (Tg), an inhibitor of sarcoplasmic/endoplasmic reticulum Ca2+ ATPase [30]. As shown in Fig. 2D, both Tm and Tg had similar effects on miR-1244 levels and reduced this miRNA expression significantly from 8 to 16 h of ER stress exposure. Notably, as shown by hierarchical clustering analysis, reduction of miR-1244 in no ER stress condition by its antagomiR supports expression of prosurvival factors including IRE1, XBP1s, and BIP and therefore resembles proadaptive UPR activity. Importantly, in no stress conditions, miR-1244 overexpression increases the levels of CHOP (DDIT3) and BBC3, and thus promotes apoptotic signals (Fig. 2C). Taken together, these data suggest that the downregulation of miR-1244 during UPR supports the activities that restore ER homeostasis and assure survival.
Notably, however, none of the miR-1244 UPR related potential targets had the clearly bidirectional pattern (decrease/increase expression) upon mimic and inhibitor (antagomiR) treatments, respectively, in all 3 experimental settings (Supplemental Fig. 3). Both ERN1 and XBP1s contain putative miR-1244 biding sites, however, miRNA overexpression had no significant effect on these two transcripts, whereas miR-1244 antagomiR rescued XBP1s mRNA only during Tg treatments (Supplemental Fig. 3A). Notably, however, the inhibition of miR-1244 resulted in accumulation of ERN1 transcript in both ER stress models (Supplemental Fig. 3B). As shown in Supplemental Fig. 3CD, miR-1244 overexpression had a mild reducing effect on both PERK and ATF6 mRNA levels in Tg treated cells only, while antagomiR was ineffective in all conditions. A similar pattern has been observed by magnesium transporter 1 (MAGT1), where miR-1244 overexpression related downregulation was observed in all conditions, while antagomiR remained ineffective (Supplemental Fig. 3E). Mild effects of both mimic and antagomiR during ER stress were also observed for ER degradation enhancing alpha-mannosidase like protein 1 (EDEM1) (Supplemental Fig. 3F). The mRNA of this potential target of miR-1244, was slightly elevated by antagomiR in both Tm and Tg models but reduced by mimic only after Tm. Interestingly, both miR-1244 inhibition and overexpression resulted in reduced heat shock protein family D (Hsp60) member 1 (HSPD1) levels during ER stress (Supplemental Fig. 3G). Importantly, despite the fact that BIP transcript lacks mir-1244 binding site, the levels of this mRNA were significantly induced in the presence of miR-1244 inhibitor in ER stress conditions (Tm and Tg), while mimic had only mild reducing effect on this transcript during Tm treatment (Supplemental Fig. 3H). Taken together, this data suggests that reduction of miR-1244 during ER stress supports IRE1 signaling as well as BIP activity.
In contrast, this miRNA could provide a link to the mitochondrial UPR, since both overexpression and inhibition of miR-1244 reduced the expression of a crucial mitochondrial chaperone, HSPD1 [31]. The link between miR-1244 and mitochondrial homeostasis is also reflected in the expression profiles of its two other potential targets, PUMA (BBC3) and NOXA (PMAIP1), that link the UPR with mitochondrial apoptosis [32]. As shown in Supplemental Fig. 4A, miR-1244 overexpression elevates the BBC3 levels in no stress conditions, whereas during Tm and Tg treatments, both mimic and antagomiR significantly attenuated the levels of this transcript. miR-1244 related effects were also observed for NOXA (Supplemental Fig. 4B). Furthermore, deregulation of miR-1244 expression during ER stress, resulted in significantly lower levels of another potential direct target of this miRNA – antiapoptotic XIAP, that is also responsible for controlling caspase activity (Supplemental Fig. 4C). Despite the fact that miR-1244 overexpression was indirectly elevating CHOP levels under control conditions, it was not effective in Tm treated cells, whereas during Tg treatments, mimic transfection also resulted in reduced levels of this transcript that were similar to antagomiR (Supplemental Fig. 4D). Although this data does not pinpoint which of the cell death pathways involved transcripts is a crucial target of miR-1244, they suggest that maintaining lower levels of this miRNA during ER stress may be crucial for maintaining survival for a longer period of time.
We also attempted to decipher the UPR mechanism responsible for miR-1244 reduction during ER stress. hsa-miR-1244 (also known as miR1244-1) is encoded on Chromosome 2: 231,713,314 − 231,713,398 forward strand (ENSG00000284378) and conserved only between Homo sapiens and Pan troglodytes. This genomic location is also occupied by prothymosin alpha, the PTMA gene (Chromosome 2: 231,706,895 − 231,713,551), but the PTMA expression remained constant during the ER stress (Supplemental Fig. 5). This suggests that downregulation of hsa-miR-1244 is independent of PTMA. Furthermore, we have not located an IRE1 consensus sequence within pre-miR-1244 and therefore excluded the possibility that this miRNA can be reduced via IRE1 associated degradation during ER stress. Therefore, to connect miR-1244 levels with the main arms of UPR, we have used inhibitors of the UPR branches that include 4µ8C for IRE1, ceapin-A7 for ATF6, and ISRIB that limits the PERK activities [33,34,35]. We applied these compounds during ER stress induced by Tm or Tg for 16 h. As shown in Fig. 3A, miR-1244 levels were increased in the presence of the ISRIB and ATF6 inhibitor, whereas the IRE1 inhibitor led to further reduction of this miRNA expression in Tm treated cells. A similar, but not identical pattern was observed in Tg treated cells, where ISRIB, ceapin-A7, and 4µ8C all rescued the miR-1244 levels. Taken together, the data pinpointed PERK and ATF6 signals as crucial for miR-1244 downregulation during ER stress and assigned IRE1 pathway with a modulatory role.
Fig. 3
UPR pathway inhibitors affect miR-1244 levels during ER stress. (A) HeLa cells were treated with Tm (2.5 µg/ml) or Tg (500 nM) for 16 h with DMSO vehicle (Ctrl) or ISRIB (1 µM), 4µ8C (50µM), ceapin-A7(4µM), and total RNA enriched in miRNAs was isolated and miR-1244 levels were quantified by qPCR and normalized to RNU48. Data represents the mean ± SD of three experiments. *P < 0.05, **P < 0.001, ***P < 0.0001, were considered significant. (B) The summary of the gene ontology analysis of transcription factors binding sites in the pre-miR-1244 promoter region is shown. (C) HeLa cells were transfected with miR-1244 mimic, inhibitor or scramble control were exposed to ER stress for 16 h (Tm or Tg) and the caspase 3/7 activity was accessed and expressed as normalized relative light units were monitored every 15 min. The results from 6 measurements (n = 6) are plotted. Error bars represent standard deviations. P < 0.05, **P < 0.001, ***P < 0.0001 were considered significant
Next, we analyzed the promoter regions (chr2:231705255–231718193 (GRCh38/hg38)) for the UPR related factors using GeneHancer software [36]. This tested if the region was able to bind 336 transcription factors (TFs), among which a significant number were identified to be downregulated in mice with the XBP1 knockout, while some of the others were specific for cellular stress responses (Fig. 3B). Interestingly, some of them were assigned to NOXA signaling, and that could explain why the expression of this gene was dramatically downregulated by both miR-1244 mimic and antagomiR (Fig. 3B). Notably, a group of UPR related transcription factors was identified as well and included TFs specific for all 3 main branches of these pathways: ATF6, IRE1 and PERK (ATF4) (Fig. 3B). Although these data do not define specific transcription factor responsible for miR-1244 expression during ER stress, they are in good agreement with the effects of UPR inhibitors and indicate that the crosstalk between the all main UPR branches determines the extend of miR-1244 reduction.
Since the miR-1244 target genes included both adaptive and apoptotic activities of UPR, to further investigate the impact of this miRNA on cell fate response to ER stress, we followed caspase 3 and 7 activities and HeLa cell viability using holomicroscopy based real time analysis [26] during UPR activation with both stressors and in the presence of miR-1244 analog and inhibitor.
As shown in Fig. 3C, after 16 h of exposure to both stressors, the caspase activity measured by quantifying the signal from the proluminescent caspase-3/7 DEVD-aminoluciferin substrate was moderately elevated, while miR-1244 overexpression increased the caspase activity in both stressed and nonstressed HeLa cells. Importantly, the relative luciferase signals observed during ER stress were comparable to previously reported in [18]. Notably, antagomiR transfection had a mild but significant reducing effect on ER stressed cells, while it increased significantly caspase activity in nonstress conditions to the levels observed in stress conditions.
To better understand this the cell fate decision dynamics, we performed real time and label free holographic microscopy-based monitoring of cell death and viability using a HoloMonitor® time-lapse cytometer. Holographic microscopy was used to follow the optical thickness and irregularity of cells exposed for up to 24 h to Tm and Tg in the presence or absence of miR-1244 mimic or antagomiR (Figs. 4, 5 and 6 and Supplemental Figs. 6–8). Healthy cells are irregular in shape and thin, whereas dying cells are round and thick [25,26,27,28]. As shown in Fig. 4 and Supplemental Fig. 6, antagomiR had no significant effect on cellular survival in nonstress conditions, whereas after 8 h of experiment the miR-1244 mimic increased the number of dying cells even under nonstress conditions. In HeLa cells exposed to Tm, there was a significant increase in dying cells after 16 h of ER stress exposure, however, miR-1244 antagomir dramatically inhibited cell death after 16 and 24 h (Fig. 5, Supplemental Fig. 7). In Tg treated cells, the miR-1244 mimic significantly decreased the number of healthy cells starting at 8 h, and the antagomiR had similar levels of healthy cells as the Tg only treated cells, indicating that the antagomiR had no effect on cell survival during the Tg treatment (Fig. 6, Supplemental Fig. 8). In summary, the results suggest that ER stress reduction of miR-1244 expression contributes to the pro-survival arm of UPR.
Fig. 4
Overexpression of miR-1244 promotes cell death. The results of real-time monitoring of cell viability are shown with the real time and label free holographic microscopy using a HoloMonitor M4® time-lapse cytometer of HeLa cells transfected with miR-1244 mimic or inhibitor or the scramble control and 48 h later monitored up to 24 h. Images were collected every 15 min (from 8 independent optical fields), and the distribution of live (blue) and dying cells (red) based on their optical thickness (Y-axis) and irregularity (X-axis) is presented at the 0, 8, 16 and 24 h time points. The images from up to 5 independent optical fields were collected and analyzed according to manufacturer’s instructions with HoloMonitor® App Suite software. Representative samples are shown in Supplemental Fig. 6. For all analyses, the same cell parameter qualifications were applied. Experiments were performed in triplicate. Based on the cells irregularity and average optical thickness the percentages of healthy cells (A) and of dying cells (B) were calculated. Data represents the mean ± SE of three independent experiments. *P < 0.05, **P < 0.001, ***P < 0.0001 were considered significant
Fig. 5
miR-1244 influences the fate of cells challenged with Tm induced ER stress. The results of real-time monitoring of cell viability with the real time and label free holographic microscopy are shown using a HoloMonitor M4® time-lapse cytometer of HeLa cells transfected with miR-1244 mimic or inhibitor or the scramble control and 48 h later treated with Tm (2.5 µg/ml) up to 24 h. Images were collected every 15 min (from 5 independent optical fields), and the distribution of live (blue) and dying cells (red) based on their optical thickness (Y-axis) and irregularity (X-axis) is presented at the 0, 8, 16 and 24 h time points. The images from up to 5 independent optical fields were collected and analyzed according to manufacturer’s instructions with HoloMonitor® App Suite software. Representative samples are shown in Supplemental Fig. 7. For all analyses, the same cell parameter qualifications were applied. Experiments were performed in triplicate. Based on the cells irregularity and average optical thickness the percentages of healthy cells (A) and of dying cells (B) were calculated. Data represents the mean ± SE of three independent experiments. *P < 0.05, **P < 0.001, ***P < 0.0001 were considered significant
Fig. 6
Exogenous miR-1244 influences the fate of cells challenged with Tg induced ER stress. The results of real-time monitoring of cell viability with the real time and label free holographic microscopy are shown using a HoloMonitor M4® time-lapse cytometer of HeLa cells transfected with miR-1244 mimic or inhibitor or the scramble control and 48 h later treated with Tg (500 nM) up to 24 h. Images were collected every 15 min (from 5 independent optical fields), and the distribution of live (blue) and dying cells (red) based on their optical thickness (Y-axis) and irregularity (X-axis) is presented at the 0, 8, 16 and 24 h time points. The images from up to 5 independent optical fields were collected and analyzed according to manufacturer’s instructions with HoloMonitor® App Suite software. Representative samples are shown in Supplemental Fig. 8. For all analyses, the same cell parameter qualifications were applied. Experiments were performed in triplicate. Based on the cells irregularity and average optical thickness the percentages of healthy cells (A) and of dying cells (B) were calculated. Data represents the mean ± SE of three independent experiments. *P < 0.05, **P < 0.001, ***P < 0.0001 were considered significant
留言 (0)