
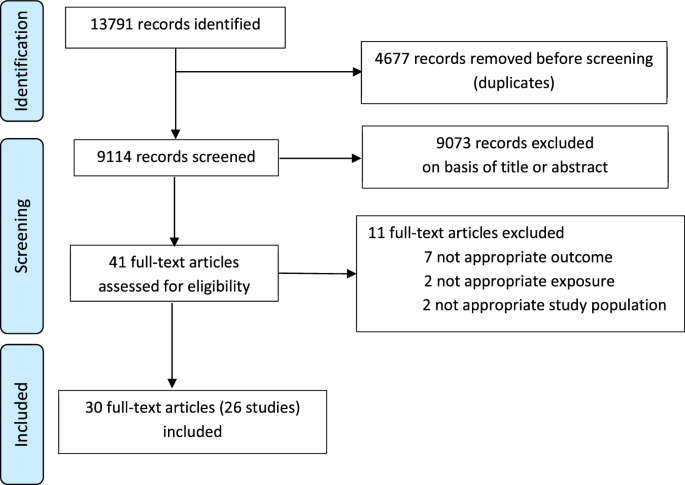
記住我

To examine whether anti-PvRBP2b or anti-TfR1 antibodies could inhibit P. vivax invasion into human reticulocytes we applied a flow cytometry-based invasion assay. In a first experiment using three different clinical isolates, we evaluated the impact on reticulocyte invasion of 3E9 mouse monoclonal anti-PvRBP2b Abs previously shown to inhibit both the binding of PvRBP2b to TfR1 and the invasion of a few Thai and Brazilian isolates17. As negative inhibition control, we included the 8G7 anti-PvRBP2b mouse monoclonal Abs (mAbs), shown not to affect either the binding of PvRBP2b to TfR1 or invasion into reticulocytes, as well as 043038, a humAbs specific of tetanus toxin C terminal fragment. An anti-Duffy mouse mAbs, 2C3, was used as positive inhibition control19. All mAbs were tested at 100 μg/ml. As expected, both negative inhibition controls did not inhibit reticulocyte invasion, however we observed only a modest (mean 17.4% ± 1.73 SEM), non-significant invasion inhibition by 3E9 (p = 0.2683) compared to the 043038 control (Fig. 1A, raw invasion data are provided in Supplementary Data 1).
Fig. 1: Anti-PvRBP2b antibodies do not inhibit the invasion of Cambodian clinical isolates.
The mean percentage (± SEM) of reticulocyte invasion by P. vivax clinical isolates in the presence of different anti-PvRBP2b antibodies tested at 100, 250, and 500 μg/ml is represented. All invasions are normalized to no-antibody controls. HumAbs 043038 specific of tetanus toxin C terminal fragment and the anti-Duffy mouse mAbs 2C3 were used as negative and positive invasion inhibition controls for each experiment. Each dot represents the invasion for a different clinical isolate with a single technical replicate per isolate. Differences are assessed by the Kruskal-Wallis test with Dunn’s post-hoc correction (*p < 0.05; ***p = 0.0007; ****p < 0.0001). A Invasion of P. vivax in presence of 100 μg/ml of mouse anti-PvRBP2b mAbs 3E9 and 8G7. B Invasion of P. vivax in presence of 500 μg/ml (100 μg/ml for the 2C3 control) mouse anti-PvRBP2b mAbs 8G7, 3E9, 6H1, 10B12, mAbs pool (each mAb at one-third of the final concentration) and rabbit polyclonal mAbs (1527). C Invasion of P. vivax in presence of 100 μg/ml of anti-PvRBP2b humAbs 239229 and 241242. D Invasion inhibition of P. vivax in presence of 500 μg/ml (100 μg/ml for the 2C3 control) of humAbs 237235, 239229, 241242, 253245, 258259, 260261, 326327. E Invasion inhibition of P. vivax in the presence of 500 μg/ml (100 μg/ml for the 2C3 control) anti-PvRBP2b humAbs combinations (250 μg/ml each huAbs). F Invasion inhibition of P. vivax in presence of anti-PvRBP2b humAbs (2392209 or 241242 each at 250 μg/ml) or 043038 control in combination with anti-PvDBP 099100 at 250 μg/ml (final concentration of humAbs of 500 μg/ml) compared to invasion in presence of 099100 alone at 250 μg/ml or 2C3 control at 100 μg/ml.
We then increased the concentration of mAbs to 500 μg/ml and evaluated two additional anti-PvRBP2b mouse monoclonal (6H1, 10B12) and a rabbit polyclonal Abs (1527), all previously shown to inhibit binding and invasion17. Overall, using 19 different clinical isolates, we show that none of the anti-PvRBP2b mAbs or the 1527 polyclonal Abs significantly inhibited invasion into reticulocytes compared to the 043038 control (p > 0.9999 for all comparisons) (Fig. 1B). The highest average inhibition was of 17.8% (±2.69 SEM) and was observed for the 8G7 negative control mAbs. We also evaluated the combination of the three mAbs 3E9, 6H1 and 10B12 together (mAbs pool) each added at a third of 500 μg/ml and again, we failed to observe any significant invasion inhibition (p > 0.9999). Targeting TfR1 by the addition of 500 μg/ml of OKT9 mAb, previously shown to inhibit the binding of PvRBP2b to TfR1, did not lead to significant invasion inhibition (p > 0.9999) (Fig. 1B).
Naturally acquired human monoclonal anti-PvRBP2b antibodies do not inhibit P. vivax reticulocyte invasionWe determined whether naturally acquired anti-PvRBP2b humAbs isolated from Cambodian donors could inhibit reticulocyte invasion by Cambodian P. vivax isolates. These humAbs have previously been characterized with their PvRBP2b binding sites determined and have been shown to inhibit the binding of PvRBP2b to TfR1 and to reticulocytes18.
We first evaluated two different humAbs at 100 μg/ml, a concentration at which we previously showed that anti-PvDBP humAbs inhibit P. vivax invasion12. These two humAbs, 239229 and 241242 bind to non-overlapping epitopes on PvRBP2b. Using 15 different clinical isolates, we show that none of these two humAbs inhibit the invasion of P. vivax in reticulocytes compared to the 043038 control (p > 0.7905) (Fig. 1C).
We then increased the concentration to 500 μg/ml and evaluated five additional blocking humAbs. We also included the humAb 099100 as an additional positive inhibition control targeting PvDBP12,20,21. Using 25 different clinical isolates, no significant inhibition was observed for any of the seven anti-PvRBP2b humAbs (Fig. 1D). The highest average inhibition measured was 18.4% (±2.97 SEM) for the humAb 260261 and was not significantly different from the 043038 control (p > 0.999).
The results we obtained here contrast with those previously reported, so we investigated whether our invasion assay readout could affect the results. Our assay relies on flow cytometry to measure P. vivax invasion, while previous work showing anti-PvRBP2b activity used microscopy readout17. We tested the activity of 500 μg/ml of two humAbs (239229 and 241242) and the 1527 polyclonal anti-PvRBP2b using three to eight different clinical isolates and measured invasion by microscopy side-by-side with flow cytometry for most isolates. No significant difference was observed comparing the invasion rates obtained with flow cytometry or microscopy and regardless of the readout, no invasion inhibition was observed (Supplementary Fig. S2).
Combination of anti-PvRBP2b humAbs together or with anti-PvDBP humAbs does not increase P. vivax invasion inhibitionAs some anti-PvRBP2b humAbs have different inhibitory mechanisms, it was previously proposed that some combinations of humAbs could act synergistically or additively18. We therefore evaluated the combination of 241242 targeting the N-terminal domain of PvRBP2b and leading to steric hindrance with TfR1with (i) 239229 binding to the C-terminal domain of PvRBP2b, (ii) 237235 believed to disturb PvRBP2b binding to transferrin allosterically, (iii) 326327 causing steric hindrance with the reticulocyte membrane18. We also evaluated the combination of 237235 with 326327. All combinations used 250 μg/ml for each humAb (500 μg/ml in total) and were tested with three different clinical isolates. None of the combinations tested showed invasion inhibition, with the highest average inhibition measured being 5.26% (±7.70 SEM) (Fig. 1E).
Finally, we performed a series of invasion assays to determine if adding 239229 or 241242 anti-PvRBP2b humAbs (binding respectively to the C and N-terminal of PvRBP2b) would enhance invasion inhibition caused by 099100 anti-PvDBP humAb. All combinations were done using 250 μg/ml for each humAb (500 μg/ml) and tested with four to five different clinical isolates. There was no significant difference in invasion inhibition between the combination of anti-PvDBP 099100 with 239229 (average inhibition 53.8% ± 7.1) or with 241242 (average inhibition 55.1% ± 4.5) compared to the combination of 043038 non-specific control with 099100 (average of 51.9% ± 3.2, p > 0.999) (Fig. 1F). The invasion inhibition was also not different between 099100 alone tested at 250 μg/ml (average inhibition 47.1% ± 4.8) and the combination of 043038 with 099100 (p > 0.999).
Sequence polymorphism of PvRBP2b among P. vivax clinical isolates does not affect inhibition caused by most anti-PvRBP2b AbsTo determine whether PvRBP2b polymorphism could explain the lack of inhibition by anti-PvRBP2b Abs, the PvRBP2b gene was sequenced to cover amino acid positions from 100 to 600 from all 50 clinical isolates used in the invasion assays.
Overall, the gene was highly polymorphic, with 44 different protein haplotypes identified among the 50 isolates sequenced, all of which differed from the Sal1 reference (Supplementary Data 2).
We stratified the invasion data by assessing the impact of PvRPB2b polymorphism within known epitopes recognized by the Abs tested. For each known epitope, we determined whether invasion in the presence of Abs was affected by polymorphism. The three mouse mAbs 3E9, 6H1 and 10B12 bind respectively to residues 330-343, 212-232 and 405-423 (Fig. 2A)18,22. All isolates tested with 3E9 at 500 μg/ml were WT in the residues 330-343 and were not inhibited by the mAbs (Fig. 2B). Of the four isolates tested against 6H1, one was WT in the residues 212-232 and the three others had all the same two mutations (T220K and T224K) and none were inhibited by these mAbs (Fig. 2B). Among the four isolates tested with 10B12 at 500 μg/ml, one was WT in the residues 405-423 and three others had the same single mutation (K412N) and none were inhibited by this mAbs. (Fig. 2B). These results indicate that P. vivax invasion is not inhibited by 3E9, 6H1 and 10B12, regardless of mutations in their respective epitopes.
Fig. 2: The absence of invasion inhibition by anti-PvRBP2b Abs is not dependent on PvRBP2b sequence polymorphism of P. vivax isolates.
A Epitopes recognized by anti-PvRBP2b mAbs18,22. B Invasion in the presence of 500 μg/ml of mouse anti-PvRBP2b mAbs is not affected by mutations in residues recognized by 6H1, 3E9, and 10B12 compared to the wild-type (WT). C Invasion in the presence of 500 μg/ml of humAbs is not affected by mutations in residues recognized by 237235, 239229, 253245, 258259 and 260261 compared to WT. For 241242 and 326327, all isolates had at least one mutation in the epitopes recognized by these humAbs. HumAbs 043038 specific of tetanus toxin C terminal fragment and the anti-Duffy mouse mAbs 2C3 were used as negative and positive invasion inhibition controls for each experiment. All invasions are normalized to no-antibody controls.
We similarly determined the impact of polymorphism in known epitopes18 of humAbs on their capacity to inhibit invasion. For 5 of the 7 humAbs (237235, 239229, 253245, 258259, and 260261), isolates WT in the epitopes were evaluated for their response to these humAbs and none showed significant invasion inhibition against these compared to the 043038 control (Fig. 2C). Similarly to mouse mAbs, these results indicate that P. vivax invasion is not inhibited by 237235, 239229, 253245, or 258259 regardless of mutations in their respective epitopes. For two humAbs (241242 and 326327), all isolates tested had at least one mutation in the epitopes, so the impact of polymorphism could not be determined for these (Fig. 2C).
Finally, to evaluate whether polymorphism would explain the absence of invasion inhibition by the rabbit polyclonal anti-PvRPB2b 1527, we performed IFA experiments on segmented schizonts from 4 different clinical isolates each having a different PvRBP2b haplotype. The polyclonal Abs recognized each of the isolates evaluated demonstrating that natural polymorphism does not prevent binding of the 1527 Abs to PvRBP2b (Fig. 3)
Fig. 3: Polyclonal anti-PvRBP2b 1527 Abs binds to merozoites from all four P. vivax clinical isolates tested by immunofluorescence.
Each row shows representative images of segmented schizonts for a different clinical isolate. A: bright field, B: Hoechst 33342, C: 1527, D: merged.
TfR1 is not recycled following trypsin treatmentWe verified that reticulocyte invasion of P. vivax isolates from Cambodia displays the same sensitivity to enzymes as previously described23,24. As expected, reticulocytes treated with neuraminidase or trypsin were permissive to P. vivax invasion while chymotrypsin treatment abolished invasion (Fig. 4A). As TfR1 is trypsin sensitive, these results indicate that cleaving TfR1 from the reticulocytes does not prevent invasion by P. vivax. We determined if an intracellular pool of TfR1 could be recycled on the reticulocyte surface following trypsin treatment by flow cytometry. We confirmed that trypsin treatment removed over 80% of TfR1 as seen by the reduced percentage of reticulocytes expressing TfR1. The percentage of reticulocytes expressing TfR1 continues to decrease due to the maturation in vitro of reticulocytes into normocytes (which are TfR1 negative) (Fig. 4B). This indicated that after surface removal by trypsin treatment, there is no internal pool of TfR1 that can be recycled to the surface.
Fig. 4: P. vivax Cambodian isolates are trypsin-resistant.
A Inhibition of P. vivax invasion after chymotrypsin, neuraminidase or trypsin treatment of reticulocytes. Only chymotrypsin treatment inhibits P. vivax merozoite invasion. B The absence of recycling of TfR1/CD71 on trypsin-treated reticulocytes. Trypsin treatment removed > 80% of TfR1 molecules on the surface of reticulocytes and the TfR1 levels remain below 10% to the original level. Data expressed are the % of the control (% of non-treated TfR1+ reticulocytes at Time 0). This experiment is representative of 4 separate experiments with 4 different batches of cord blood reticulocytes. All invasions are normalized to no-antibody controls.
留言 (0)