
記住我
All of the human Siglecs display broad affinity for sialoglycans [19]. However, sialic acid is incorporated into many oligosaccharide chains on cell-surface glycoproteins and glycolipids [19]. Siglec-glycan interactions can also be influenced by many factors, including: (1) the monosaccharide composition and stereochemistry of the glycan, (2) secondary interactions with non-carbohydrate elements of protein scaffolds, and (3) the valency of glycan presentation on the cell surface [29, 30]. Each Siglec thus binds a specific set of glycosylated ligands. Given this complexity, we wanted to determine what specific Siglec ligands are most commonly expressed by AML cells.
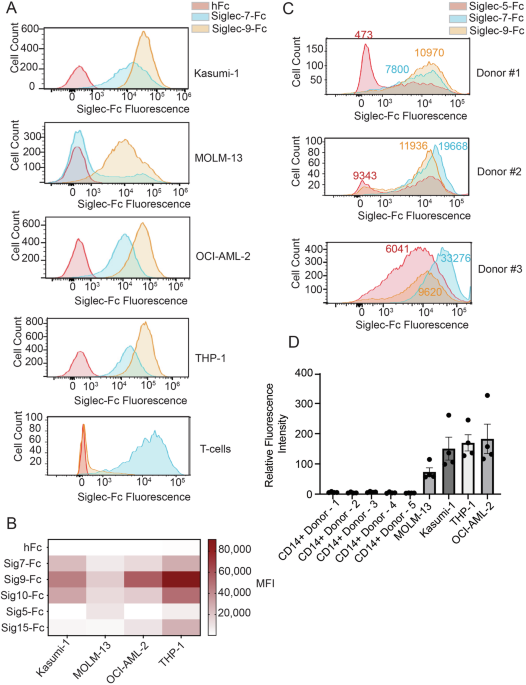
We first assembled a panel of AML cell lines including THP-1, Kasumi-1, MOLM-13, and OCI-AML-2 cells [31]. These lines are genetically heterogenous and have been extensively characterized by genomics and transcriptomics, making them useful starting models [31]. We then stained these cells with fluorescent Siglec-5, -7, -9, -10 and -15-Fc chimeras. These are dimeric staining reagents that are routinely used to detect the expression of Siglec ligands by flow cytometry [32]. We chose these specific Siglecs because they are most commonly expressed by the cytotoxic immune cells (NK cells, macrophages) that are involved in cancer immune surveillance [19]. Multiple cell lines expressed ligands for Siglecs -7, -9 and -10 (Fig. 1A, B). Conversely, ligands for Siglecs-5 and -15 were either absent or poorly expressed in most cases (Fig. 1A, B). We also observed considerable heterogeneity in Siglec-Fc staining for some members of the family. Siglec-7-Fc, for instance, bound strongly to some cell lines (e.g., OCI-AML-2) but showed little binding to others (MOLM-13). Interestingly, we found that Siglec-9 ligands were consistently expressed at the highest levels in all cell lines tested (Fig. 1B). This contrasts with prior work on T-cell acute lymphoblastic leukemia (T-ALL) and chronic lymphocytic leukemia (CLL), in which ligands for other Siglecs were typically expressed at higher levels [25, 26, 28]. Siglec-9-Fc was also found to not bind other hematopoietic cell types, such as primary T-cells, which do express Siglec-7 ligands (Fig. 1A) [33]. Siglec-9 ligands are thus selectively and abundantly expressed on AML cell lines.
Fig. 1: AML cells express Siglec-binding glycans.
A The indicated cell lines were incubated with 1 μg/mL Siglec-Fc reagents precomplexed with 1 μg/mL AlexFluor488-antihuFc for 30 minutes on ice. For T-cell staining, primary peripheral blood mononuclear cells (PBMCs) were co-stained with Siglec-Fc reagents and an anti-CD3 antibody. The cells were then analyzed by flow cytometry to detect the expression of different Siglec-binding ligands. Representative plots for each cell type are provided. For T-cells, the plot shown indicates Siglec-Fc fluorescence in CD3+ cells. B Average median fluorescence intensity (MFI) of Siglec-Fc staining in the indicated AML cell lines. The average of n = 3 independent biological replicates is plotted. C Patient-derived primary AML cells were incubated and analyzed as in (A). Representative plots are shown. Median fluorescence intensity for Siglec-Fc staining is indicated directly on the flow cytometry plot. D PBMCs from multiple healthy blood donors and the indicated cell lines were stained with Siglec-9-Fc as in (A). For PBMC samples, CD14+ monocytes were gated by co-staining with a fluorescent anti-CD14 antibody. The relative fluorescence intensity (RFI) for each sample is indicated so as to normalize for differences in autofluorescence between cell types. RFI is equal to the MFI(Sig9-Fc)/MFI(hFc). Mean values plotted, error bars indicate SEM.
As immortalized cells often exhibit phenotypic changes under in vitro growth conditions, we next wanted to explore whether patient-derived primary AML cells also expressed Siglec-9 ligands. We took cells that had been isolated from the peripheral blood and/or bone marrow of AML patients and stained them with Siglec-Fc reagents as above. As primary cell numbers were limited for this experiment, we confined our analysis to studying the expression of Siglec-7 and Siglec-9 ligands. We also used Siglec-5-Fc as a background control, given that it exhibited low binding to most of the AML cell lines we tested (Fig. 1B). All AML cells displayed high expression of Siglec-7 and Siglec-9 ligands, while Siglec-5 ligand expression was quite low, mirroring the results we obtained in cell line models (Fig. 1C). We observed similar patterns of Siglec ligand expression on CD34+/CD38- and CD34+/CD38+ populations (Supplementary Fig. 1). We did observe greater heterogeneity in Siglec-Fc staining in primary samples when compared to cell lines. This is unsurprising given the greater genetic and phenotypic heterogeneity present in primary samples vs. immortalized cell lines. Finally, we compared the expression of Siglec-9 ligands in AML cell lines to healthy myeloid immune cells. We isolated PBMCs from five healthy blood donors and analyzed the binding of Siglec-9-Fc to CD14+ monocytes. Siglec-9 ligand expression was dramatically higher in all AML cell lines than in these untransformed myeloid cells (Fig. 1D).
Taken together, these results show that AML cells broadly express Siglec-binding glycans and imply that Siglec-9 may be one target of particular interest for this disease. We, therefore decided to interrogate how Siglec-9 ligands are generated in AML cells, what specific glycan structures act as Siglec-9 ligands in AML, and whether these ligands play a functional role in driving AML immune evasion.
Genome-wide CRISPR screening reveals putative genes driving Siglec-9 ligand expression in AMLIn a previous study, we conducted several genome-wide CRISPR screens that revealed regulators of Siglec-7 and Siglec-9 ligand expression in the myeloid leukemia cell line K-562 [26]. Briefly, K-562-dCas9KRAB cells were transduced with a genome-wide library of single guide RNAs (sgRNAs), stained with fluorescent Siglec-Fc chimeras, and sorted by FACS to isolate cells that showed a decrease in Siglec ligand expression. sgRNAs enriched in this low-staining population were then identified through next-generation sequencing, producing an atlas of genes whose knockdown reduces the expression of specific Siglec ligands. By validating hits from this screen, we were then able to extensively characterize a specific O-glycan structure that binds the Siglec-7 receptor and mediates immune evasion in T-ALL [25, 26]. No further analysis or validation, however, was conducted on hits from the corresponding Siglec-9 screen. We thus chose to interrogate this dataset to identify genes that may regulate Siglec-9 ligand expression in AML specifically (Fig. 2A).
Fig. 2: Functional genomics analysis reveals ST3GAL4 as a potential driver of Siglec-9 ligand expression in AML.
A K-562-dCas9KRAB cells were transduced with a genome-wide library of sgRNAs and stained with Siglec-9-Fc as in Fig. 1A. Cells exhibiting low binding were sorted by FACS and sequenced to reveal enriched sgRNAs. Screen hits were analyzed using MAGeCK. Selection score indicates the strength of sgRNA enrichment in the low-staining population vs. an unsorted control. A positive score indicates that sgRNAs for the indicated gene were more enriched in the low-staining population. A negative score indicates they were less enriched. B Biosynthetic pathway for the synthesis of Sialyl-LacNAc-containing glycans. Biosynthetic enzymes that were recovered as screen hits are indicated. C Relative mRNA expression levels of ST3GAL4 in patient AML samples with the indicated genetic alterations (BloodSpot Database) are shown. ST3GAL4 expression in hematopoietic stem cells (HSC), common myeloid progenitors (CMP), and monocytes from healthy donors are shown as a comparison. An unpaired Student’s t-test was performed to detect statistically significant differences in mRNA expression between each AML subtype and HSCs. *** indicates P < 0.001.
In other cancer types, dysregulation of cell-surface sialylation has been directly connected to changes in the activity of specific glycan biosynthetic enzymes [34,35,36]. We, therefore filtered our hit list to identify any hit genes known to be involved in carbohydrate metabolism (using the GO term GO:0005975, carbohydrate metabolic process). As expected, this analysis identified many hit genes which are known components of the core sialic acid biosynthesis pathway (e.g., CMAS, GNE, SLC35A1) [26]. Additionally, we found several glycosyltransferases - enzymes that catalyze the linkage of a monosaccharide to a specific underlying glycan substrate. For example, knockdown of the sialyltransferase enzymes ST3GAL3 and ST3GAL4 was found to decrease Siglec-9 ligand expression (Fig. 2A). In vitro, both enzymes have been shown to catalyze addition of sialic acid to acceptor substrates containing Galβ1-4GlcNAc (LacNAc), producing Neu5Acα2-3Galβ1-4GlcNAc (Sialyl-LacNAc) (Fig. 2B) [37].
We also found that knockdown of several other glycosyltransferases actually increased the expression of Siglec-9 ligands. Interestingly, these enzymes also have been reported to act on the LacNAc motif (Fig. 2A, B). For example, B3GNT2 competes with ST3GAL3 and ST3GAL4 to catalyze the formation of GlcNAcβ1-3Galβ1-4GlcNAc (Fig. 2B) [38]. ST8SIA4, similarly, modifies the Sialyl-LacNAc motif by adding sialic acid in an α2,8-linkage (Fig. 2B) [39]. The fact that all these enzymes act on a similar substrate suggests that Siglec-9 has a distinct specificity for glycans containing Sialyl-LacNAc [39]. This finding would be broadly consistent with several reports describing the binding of Siglec-9 to synthetic glycan arrays [40]. While glycosylation can be quite cell-type specific, ST3GAL enzymes have also been implicated as regulators of Siglec-9 ligand expression in some cell-based models, including in HEK293 cells and pancreatic cancer cell lines [34, 36, 41].
We then performed several other analyses to determine whether any of these genes exhibited altered expression in AML. First, we consulted the BloodSpot database [10], which provides a detailed characterization of mRNA expression in both healthy and malignant hematopoiesis. Expression levels of ST3GAL3 and B3GNT2 were broadly similar across different genetic subtypes of AML as well as in healthy hematopoietic precursors (Supplementary Fig. 2A, B). However, we found a marked upregulation of ST3GAL4 expression in several subtypes of AML when compared to terminal monocytes or hematopoietic stem cells (Fig. 2C). Secondly, we interrogated DepMap [42] to analyze the expression of ST3GAL4 across a broad panel of AML cell lines. We observed that AML cell lines exhibited much higher expression of ST3GAL4 than cell lines derived from other forms of blood cancer (Supplementary Fig. 3). These data imply that the remodeling of AML glycosylation may be at least partially driven by upregulation of ST3GAL4 expression.
Several prior studies have explored the function of ST3GAL4 in AML and myeloid cell biology. One previous work found that elevated expression of ST3GAL4 was associated with worse survival in a large cohort of AML patients [43]. Mechanistically, other studies have proposed that ST3GAL4 may drive AML progression by synthesizing Sialyl Lewis X antigens [43, 44]. These glycans have been found to act as ligands for the selectin family of receptors, facilitating the survival of AML stem cells in the bone marrow niche [43]. However, the possible role of ST3GAL4 in generating ligands for other glycan-binding receptors has not been interrogated. We therefore hypothesized that ST3GAL4 may also drive elevated synthesis of Siglec-9 ligands, allowing AML cells to escape from immune surveillance.
Knockout of ST3GAL4 ablates expression of Siglec-9 ligands in AML cell linesAML is a highly genetically heterogeneous disease [31]. We thus wanted to assess whether ST3GAL4 drives Siglec-9 ligand expression in a panel of genetically diverse cell models. We started by using CRISPR-Cas9 to disrupt the expression of ST3GAL4 in the four AML cell lines we originally profiled for Siglec ligand expression (Fig. 1A). Cells were lentivirally transduced with a construct encoding Cas9 and a sgRNA against ST3GAL4. Following selection, genomic DNA was extracted and the region around the expected sgRNA cut site was profiled by Sanger sequencing (Fig. 3A). Editing at the ST3GAL4 locus was quantitated by TIDE analysis (Fig. 3B) [45]. We observed significant formation of frameshift-inducing indels at the ST3GAL4 locus in all cell lines (Fig. 3B). We then stained cells with a fluorescently labeled Siglec-9-Fc chimera. In all cell lines, we observed a marked reduction in Siglec-9-Fc staining (Fig. 3C). Conversely, we observed that parallel knockout of the closely related sialyltransferase ST3GAL3 had no effect on Siglec-9 ligand expression in these AML cell lines (Supplementary Fig. 4A–D). Observing such a large, selective effect from knockout of a single sialyltransferase thus indicates a significant, specific function for ST3GAL4 in Siglec-9 ligand biosynthesis.
Fig. 3: ST3GAL4 knockout reduces expression of Siglec-9 ligands in AML cell lines.
A The indicated AML cell lines were transduced with a plasmid encoding Cas9 and an sgRNA targeting an exon of ST3GAL4. Genomic DNA was extracted from both the wild-type (WT) and ST3GAL4 knockout (KO) cell lines. PCR was performed to amplify a sequence around the sgRNA annealing site. The amplified region was sequenced and analyzed by TIDE software to quantify editing at the ST3GAL4 locus. The plot depicts a representative Sanger sequencing trace at the ST3GAL4 locus of the WT and KO cells, along with the expected break site. B The graph indicates the average percentage of insertions and deletions (indels) in the ST3GAL4 gene for the indicated AML cell lines transduced with an sgRNA against ST3GAL4. Mean values are plotted for n = 3 biological replicates, error bars indicate SEM. C WT and ST3GAL4 KO AML cell lines were analyzed for Siglec ligand expression. The average mean fluorescence intensity (MFI) of Siglec-9-Fc staining is shown for WT and ST3GAL4 KO cells. Mean values plotted for n = 3 biological replicates. Statistical significance was determined by a Student’s two-tailed t-test. *P < 0.05, **P < 0.01. D A monoclonal MOLM-13 ST3GAL4 KO cell line was generated through clonal dilution. Sanger sequencing of the ST3GAL4 locus is depicted. E MOLM-13 ST3GAL4 KO cells were transduced with a ST3GAL4 WT cDNA containing silent mutations to eliminate the sgRNA binding site. WT, ST3GAL4 KO, and ST3GAL4-transduced cells were then stained with fluorescent Siglec-9-Fc. The indicated flow cytometry plots are representative of n = 3 biological replicates.
We then used clonal dilution to generate monoclonal cell lines with defined knockouts in the ST3GAL4 gene. For these experiments, we focused on the MOLM-13 cell line, as we found these cells were best able to survive and proliferate following clonal dilution. We were able to isolate a MOLM-13 ST3GAL4 KO line with a homozygous +1 frameshift insertion in the ST3GAL4 gene (Fig. 3D). These cells demonstrated a near-complete loss of Siglec-9 ligand expression relative to WT cells (Fig. 3E). To definitively link these effects to ST3GAL4 disruption, we then transduced ST3GAL4 KO cells with a ST3GAL4 WT cDNA. Exogenous overexpression of ST3GAL4 restored Siglec-9 ligand expression back to WT levels (Fig. 3E). Taken together, this collection of data definitively implicates ST3GAL4 as the key synthase controlling expression of Siglec-9 ligands in AML cells.
Comprehensive glycomics analysis reveals the specific impact of ST3GAL4 KO on cell-surface sialylationWe next sought to define the biochemical structure of the specific Siglec-9 ligands that are synthesized by ST3GAL4. In vitro, ST3GAL4 was originally characterized to transfer sialic acid onto LacNAc-containing acceptor substrates (Fig. 2B) [46, 47]. As this motif is most commonly associated with N-linked glycans, ST3GAL4 is often presumed to act primarily on these structures [34]. However, LacNAc can also be found in certain types of O-linked glycans and neolacto-series glycolipids. Indeed, cell-based studies have shown that ST3GAL4 inhibition can affect the synthesis of Sialyl Lewis X structures on both N- and O-linked glycans [44]. It is thus likely that the effects produced by ST3GAL4 KO will vary depending on the broader glycomic and transcriptomic profile of a given cell model. Because of this, we chose to specifically characterize the effects of ST3GAL4 KO on cell-surface glycosylation patterns in our AML cell lines.
We isolated N-linked and O-linked glycans from WT and ST3GAL4 KO cells using separate methods. N-glycans were enzymatically released (e.g., with PNGaseF), while O-glycans required a chemical method (NaBH4 reduction) [48]. The relative abundance of different structures was then globally profiled by MALDI-MS. A full summary of altered glycan structures can be found in Supplementary Tables 1–3. We first broadly analyzed how the sialylation of different glycans is altered by ST3GAL4 KO. There was a dramatic decrease in the abundance of sialylated N-linked glycans, with a corresponding increase in N-linked structures that contained no sialic acid (Fig. 4A). Changes in sialylation were observed for a large number of underlying structures, with 23 out of 77 detected N-linked glycans showing statistically significant decreases in expression (Supplementary Table 1). In prior work, ST3GAL4 has been shown to specifically catalyze the formation of fucosylated Sialyl Lewis X antigens on the surface of cells [44]. However, we observed strong decreases in sialylation of both fucosylated and non-fucosylated N-linked structures (Fig. 4B, C). Similarly, branching of N-glycans did not seem to affect the cellular response to ST3GAL4 KO. We observed in our datasets that both bi-, tri-, and tetra-antennary structures displayed comparable decreases in sialylation (Fig. 4D, E). These data confirm that ST3GAL4 plays a broad role in the α2,3-sialylation of N-linked glycans and imply that these structures may serve as important Siglec-9 ligands on AML cells.
Fig. 4: ST3GAL4 KO primarily affects the expression of N-linked sialoglycans on AML cells.
A OCI-AML-2 WT and ST3GAL4 KO cells were lysed and glycans were extracted using a mix of chemical and enzymatic methods. Free glycans were permethylated and subjected to MS-based glycomics analysis using MALDI-TOF. The graph indicates the percentage of all N-linked structures that contain sialic acid. B The graph displays the percent abundance of the indicated glycan structure as a proportion of all N-linked glycans in WT and ST3GAL4 KO cells. C The graph displays the percent abundance of the indicated terminally fucosylated glycan structure as a proportion of all N-linked glycans in WT and ST3GAL4 KO cells. D The graph displays the percent abundance of the indicated biantennary glycan structure as a proportion of all N-linked glycans in WT and ST3GAL4 KO cells. E The graph displays the percent abundance of the indicated triantennary glycan structure as a proportion of all N-linked glycans in WT and ST3GAL4 KO cells. F OCI-AML-2 WT and ST3GAL4 KO cells were processed as in (A). The graph indicates the percentage of all O-linked structures containing sialic acid in WT and ST3GAL4 KO cells. G The graph displays the percent abundance of the indicated disialyl core 2 structure as a proportion of all O-linked glycans in WT and ST3GAL4 KO cells. H The graph displays the percent abundance of the indicated disialyl core 1 structure as a proportion of all O-linked glycans in WT and ST3GAL4 KO cells. In all cases, mean values are plotted for n = 3 biological replicates. Error bars indicate SEM.
We also observed that ST3GAL4 KO impacted other classes of glycosylation. The overall level of sialylation on O-linked structures was broadly similar in both WT and ST3GAL4 KO cell lines (Fig. 4F). However, we did observe some significant changes in O-glycan composition. For example, we found a statistically significant decrease in the abundance of disialyl core 1 glycosylated structures (Fig. 4G, H). This was associated with a corresponding increase in disialylated core 2 O-glycans (Fig. 4G, H, Supplementary Table 2). This was surprising because ST3GAL1, not ST3GAL4, is typically thought to be the primary sialyltransferase acting on the Galβ1-3GalNAc motif in human cells [49]. However, some studies have found that ST3GAL4 can transfer sialic acid onto Galβ1-3GalNAc-containing substrates in vitro [37]. It is thus likely that ST3GAL4 may also remodel O-linked glycosylation when overexpressed in AML cell lines. To complete the picture, we finally conducted a glycolipidomics analysis to assess whether ST3GAL4 KO impacts the sialylation of cell-surface lipids. This revealed no major difference in sialylation of lipid structures between WT and KO cells (Supplementary Table 3). Taken together, these findings imply that elevated expression of ST3GAL4 in AML cell lines broadly re-structures glycosylation of N- and O-linked glycoproteins, but not glycolipids.
Siglec-9 binds multiple classes of N-linked sialoglycansGiven these diverse effects of ST3GAL4 KO on different aspects of glycan biosynthesis, we next explored whether Siglec-9 exhibits preferential binding to specific types of Sialyl-LacNAc-containing glycans. Synthesis of complex, sialylated N-glycans requires the activity of the biosynthetic enzyme MGAT1 (Fig. 5A). We therefore used a previously described CRISPR-Cas9 KO strategy to generate cell lines with a deficiency in complex N-glycan biosynthesis [50]. We transduced OCI-AML-2 cells with an sgRNA against the MGAT1 enzyme and stained cells with the lectin L-PHA, which binds to complex N-linked glycans [51]. Transduction of cells resulted in a polyclonal cell pool containing a mixture of MGAT1 KO (L-PHA-) and WT (L-PHA+) cells (Fig. 5B). Co-staining with Siglec-9-Fc revealed that L-PHA- cells exhibited a dramatic loss of Siglec-9 ligand expression (down to ~20% of WT levels) (Fig. 5C). The magnitude of this effect was approximately equivalent to the effect of knocking out ST3GAL4 in the same cell line. Notably, staining was not fully reduced to baseline, indicating that Siglec-9 may also exhibit residual binding to some other types of structures such as O-linked glycans. However, these data conclusively demonstrate that Siglec-9 binds a ligand on AML cells that contains an N-linked Sialyl-LacNAc motif.
Fig. 5: Siglec-9 binds primarily to N-linked sialoglycans.
A Biosynthetic pathway for the synthesis of complex N-linked glycans. MGAT1 catalyzes an essential first step in this pathway. B OCI-AML-2-Cas9 cells were transduced with an sgRNA against MGAT1 and co-stained with fluorescent L-PHA (5 µg/mL) along with fluorescent Siglec-9-Fc (1.5 µg/mL) as described above. A representative flow cytometry plot and gating strategy is shown. C Mean fluorescence intensity of Siglec-9-Fc staining in L-PHA+ and L-PHA- cell populations is shown. 2′ indicates control staining with a fluorescent secondary antibody. D Biosynthetic pathway for the synthesis of terminally fucosylated N-linked glycans. FUT7 catalyzes an essential step in this pathway. E K-562-dCas9-VPR cells were transduced with sgRNAs targeting FUT7 and stained with Siglec-9-Fc as described above. The median fluorescence intensity of Sig9-Fc staining is shown. F Biosynthetic pathway for the synthesis of branched N-linked glycans. MGAT4A catalyzes an essential first step in this pathway. G K-562-dCas9VPR cells were transduced with sgRNAs targeting MGAT4A and stained with Siglec-9-Fc as described above. The median fluorescence intensity of Sig9-Fc staining is shown. Mean values are plotted for n = 3 biological replicates in all cases, error bars indicate SEM.
Our MS profiling data showed that ST3GAL4 KO broadly affects many N-linked structures containing Sialyl-LacNac. We, therefore wondered whether Siglec-9 preferentially binds to a specific structure or set of structures that incorporate this motif. We first assessed whether increased fucosylation of this epitope would alter the cell-surface binding of Siglec-9-Fc. As there are several functionally redundant fucosyltransferase genes that act on Sialyl-LacNac [52], we reasoned that it would be impractical to disrupt fucosylation through a genetic knockout approach. Instead, we applied a gain-of-function CRISPR activation (CRISPRa) strategy to overexpress FUT7 (which fucosylates the N-linked Sialyl-LacNac structure, Fig. 5D) [52]. sgRNAs against this enzyme were transduced into cells expressing dCas9-VP64 (a transcriptional activation domain). Significantly elevated expression of FUT7 was observed following transduction (Supplementary Fig. 5). However, no significant impacts on Siglec-9-Fc binding were observed (Fig. 5E). We adopted a similar approach to assess whether perturbation of N-glycan branching alters cell-surface binding of Siglec-9-Fc. Here, we used CRISPRa to overexpress the branching enzyme MGAT4A (Fig. 5F, Supplementary Fig. 5). Again, no impact on Siglec-9-Fc binding was observed despite strong upregulation of enzyme expression at the mRNA level (Fig. 5G, Supplementary Fig. 5). These data indicate that the binding of Siglec-9 to N-linked Sialyl-LacNac is largely independent of the glycan context in which this structure is presented.
ST3GAL4 inhibits phagocytosis of AML cells by macrophages through the synthesis of Siglec-9 ligandsSiglec-9 is a key inhibitory receptor expressed by phagocytic cells like macrophages and dendritic cells [53]. Macrophages are an important player in restraining the growth of AML cells in the bone marrow microenvironment [54]. We, therefore wanted to investigate if the loss of Siglec-9-binding ligands on the surface of AML cells makes them more sensitive to phagocytic attack by macrophages. First, we isolated primary monocytes from healthy PBMCs and differentiated them into macrophages using a previously described protocol [21]. After differentiation, cells expressed high levels of Siglec-7 and Siglec-9 as well as the macrophage markers CD11b and CD14 (Fig. 6A). To assess the impact of ST3GAL4 KO on phagocytosis, we labeled MOLM-13 WT or ST3GAL4 KO cells (Fig. 3D, E) with the cell-permeable dye CellTrace Far Red (CTFR). We then co-cultured these cells with mature macrophages for 4 hours (Fig. 6B). We measured phagocytosis of AML cells by quantitating the overall increase in CTFR fluorescence in CD11b+ cells (Fig. 6C–E). ST3GAL4 KO cells displayed significantly more sensitivity to phagocytosis when compared to WT cells, indicating a role for this enzyme is promoting immune evasion by AML cells.
Fig. 6: ST3GAL4 KO sensitizes AML cells to phagocytosis.
留言 (0)