
記住我

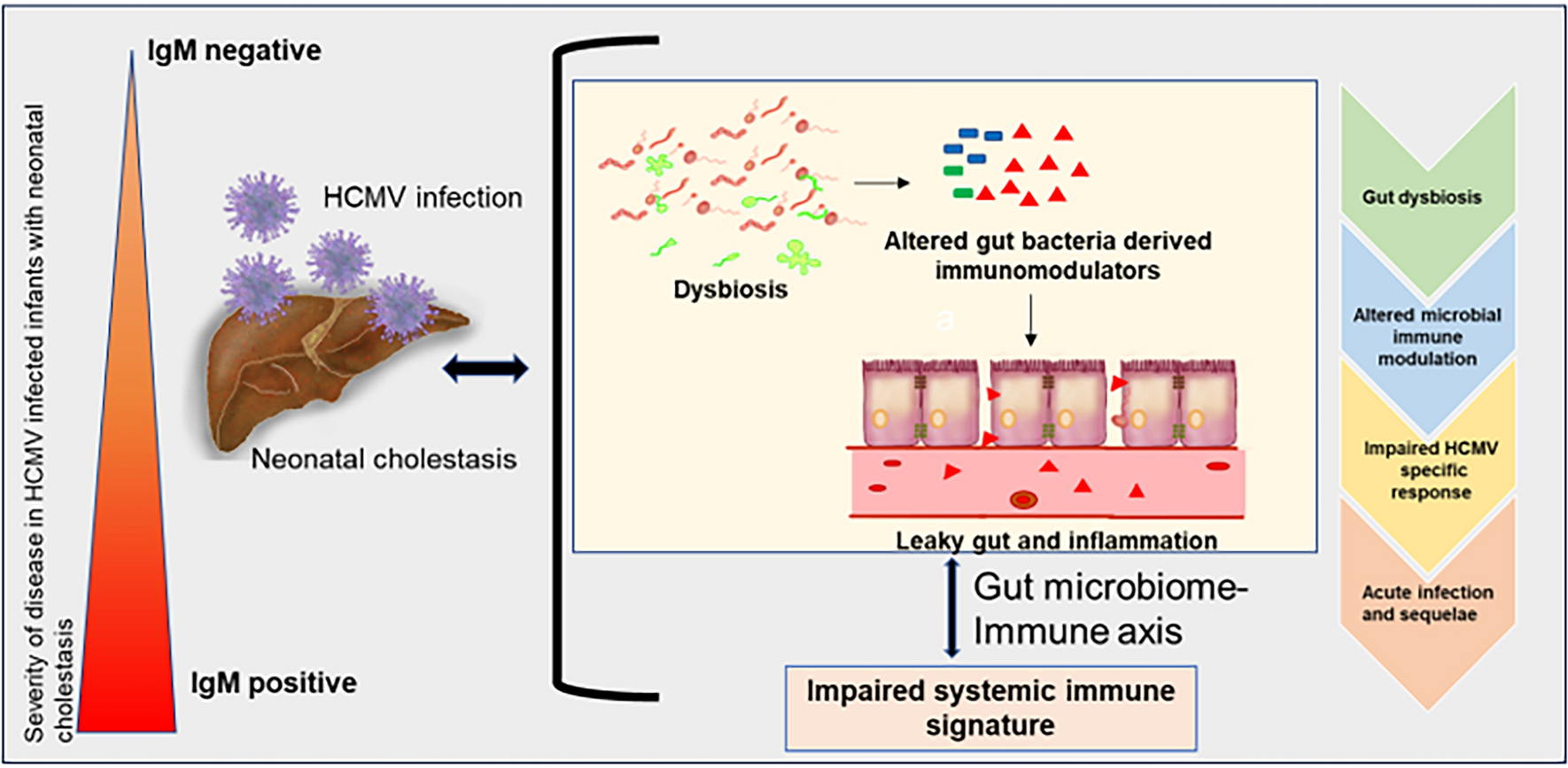




After assessing the impact of the observed dysbiosis (Supplementary Fig. 1), major gut bacteria-derived short-chain fatty acids namely acetate, butyrate, and propionate were quantified in the fecal samples of healthy and HCMV infected groups. We observed that fecal acetate levels were the highest in the healthy group 175 (54.50–453.2) mmol/L Fig. (1A), which corresponded to the highest Bifidobacterium: Enterobacteriaceae ratio reported for the group above (Supplementary Fig. 1B). The levels of acetate were significantly lower in both HCMV infected groups. There was a strong trend, although not significant (p = 0.321), towards higher acetate levels in the IgM negative group, 102.3 (8.524–262.8) mmol/L compared to IgM positive group, 47.97 (1.349–173.3) mmol/L which was supported by ROC curve analysis (AUC, 0.7556; Fig. 1A, Supplementary Fig. 2). Butyrate levels were significantly reduced in the IgM positive group, 5.786 (3.474–7.461) mmol/L compared to the healthy, 13.44 (3.409–44.38) mmol/L and IgM negative group, 9.196 (4.325–35.82) mmol/L which was also supported by the ROC curve analysis, with no significant variation in propionate levels. Fecal SCFA analysis was corroborated by PICRUSt2 analysis (Fig. 1B), wherein progressive lower representation of pathways involved in the production of acetate and butyrate from healthy to IgM negative and ultimately IgM positive infants was seen. The high abundance of Enterobacteriaceae observed in the dysbiotic microbiome of HCMV infected infants was further confirmed with a significant and progressively higher level of fecal LPS, which was observed within infected individuals, with the IgM positive group having the highest levels, 14.76 (3.774–36.87) pg/g followed by the IgM negative group, 7.918 (2.590–16.53) pg/g and finally healthy controls, 4.471 (1.702–6.742) pg/g respectively (Fig. 1A). Furthermore, PICRUSt2 analysis, predicted significantly higher biosynthetic potential of gut microbiota for LPS in HCMV infected infants with NC (Fig. 1B).
Fig.1
Estimation of fecal microbial products– (A) Levels of fecal short-chain fatty acids assessed by HPLC and fecal LPS by ELISA in HCMV infected infants (N = 15 each) with NC compared to healthy controls (N = 10) along with significant ROC curve analysis for butyrate and LPS; (B) Relative proportion of predicted significant Metacyc pathways associated with SCFA synthesis obtained by PICRUSt2 analysis. Data represented as Median. Statistical significance calculated by Kruskal –Wallis test with Dunn’s multiple comparison test; *, p < 0.05; **, p < 0.01, ***, p < 0.001; and ****, p < 0.001
Taken together, these results clearly demonstrate a disparate and dysbiotic gut microbiome in HCMV infected infants with NC that could contribute to a potentially inflammatory milieu in the gut.
Elevated fecal inflammatory markers in HCMV-infected infants with NCAs the dysbiotic signature present in HCMV infected individuals was suggestive of a pro-inflammatory milieu, we quantified the fecal levels of cytokines and markers associated with intestinal inflammation such as calprotectin, β-Defensin 2, and myeloperoxidase (MPO). We observed significantly higher and apparently hierarchical levels of all pro-inflammatory cytokines evaluated in HCMV infected infants with NC compared to healthy controls with IgM positive individuals seeming to have the highest levels (Fig. 2A). This observation also extended to all three inflammation associated markers evaluated. A trend (based on median values), though not significant was observed for the anti-inflammatory cytokine IL-10 with highest levels observed in healthy controls, 652.7 (353–971.6) pg/g and progressively lower levels observed in HCMV infected individuals and no clear trend in TGF-β. ROC curve analysis ascertained the discriminatory potential of some of these significant signatures—IL-1β, IL-6, IL-23 and MPO (Fig. 2B and Supplementary Fig. 2).
Fig.2
Estimation of fecal inflammatory markers—(A) Levels of fecal cytokines and (B) gut inflammatory markers assessed by ELISA in HCMV infected infants with NC (N = 15 each) compared to healthy controls (N = 10); (C) Significant ROC curve analysis of fecal inflammatory markers between HCMV infected infants with NC. Data represented as Median. Statistical significance was calculated by and Kruskal–Wallis test with Dunn’s multiple comparison test; *, p < 0.05; **, p < 0.01, ***, p < 0.001; and ****, p < 0.001
Loss of gut integrity and microbial translocation in HCMV infected infants with NCAfter elucidating a distinct dysbiotic microbiome and a corresponding pro-inflammatory signature present in HCMV infected individuals, we evaluated fecal levels of sIgA, known to regulate homeostatic composition of the gut microbiota [19] (Fig. 3A), where an incremental and progressively lower level of sIgA was observed from healthy controls, 4546 (3351–5452) µg/g of feces to HCMV infected IgM negative, 2960 (571.7–4355) µg/g of feces and IgM positive individuals, 2378 (910.4–3930) µg/g of feces respectively. Next, we evaluated markers such as fecal protease, indicative of inflammation [28], and zonulin levels, representative of gut permeability [20] (Fig. 3A). Both these markers were significantly and incrementally elevated compared to healthy controls in IgM negative and IgM positive infants respectively, suggestive of impaired gut barrier function within infected individuals. Interestingly, concurrent estimation of plasma LPS revealed detectable levels, which were significantly higher in IgM positive individuals, 31.76 (6.628–68.47) pg/ml. ROC curve analysis (Fig. 3B), depicts the clear discriminatory potential of these markers in IgM positive and IgM negative HCMV infected individuals (AUC-0.72). Taken together, these results clearly demonstrate the occurrence of microbial translocation in conjunction with increased gut permeability within HCMV infected infants with NC.
Fig.3
Estimation of fecal gut-integrity and bacterial translocation markers–(A) Levels of fecal metabolites assessed by ELISA in HCMV infected infants with NC (N = 15 each) and healthy controls (N = 10); (B) Significant ROC curve analysis of fecal gut-integrity markers between HCMV infected infants with NC. Data represented as Median. Statistical significance was calculated by and Kruskal–Wallis test with Dunn’s multiple comparison test; *, p < 0.05; **, p < 0.01, ***, p < 0.001; and ****, p < 0.001
Microbiome-immune network interactions characterizing HCMV mediated pathology in infants with NCIn an effort to integrate the triad of dysbiotic gut-microbiome, associated microbial products, and observed host immune phenotypes with the pathology and severity of HCMV infection in the context of NC, we obtained an extensive correlation network (Supplementary Fig. 3). Figure 4 depicts sub-networks that represent interactions for deciphering potentially actionable pathways for therapeutic interventions in future. Our previously reported associations of Granzyme B expressing NK cells, Gammaproteobacteria and Finegoldia magna [14] (reproduced as supplementary Fig. 5) were further extended to include a positive association with fecal LPS (r = 0.5293) and a negative association of this subset with fecal butyrate levels (r = − 0.4686). The latter, in turn, were positively correlated with Bifidobacterium (r = 0.5492) and negatively associated with fecal LPS (r = − 0.6027) (Fig. 4A). With respect to the T cell compartment (Fig. 4B), we observed a similarly strong negative correlation between CD4 positive Granzyme B expressing T cells and Finegoldia magna (r = − 0.5497). Also, a distinct correlation network involving Firmicutes and the CD4/CD8 ratio, a known disease progression marker, was seen, where frequency of this taxa was independently correlated with circulating frequencies of CD4 positive (r = 0.6509) and CD8 positive T cells (r = − 0.6509) and in turn, positively with the CD4/CD8 ratio (r = 0.6509). Interestingly, our previous report on this cohort showed depleted levels of Firmicutes in IgM positive HCMV infected individuals with concurrently lower CD4/CD8 ratios. Propionate and acetate were both positively correlated with this ratio. In addition, propionate was also observed to have a negative correlation with frequency of circulating cytotoxic CD8 positive T cells expressing Granzyme B (r = − 0.5485). Interestingly, we also noted a positive correlation of propionate with fecal LPS (r = 0.3376), suggestive of a potential compensatory response to elevated levels of the former. Next (Fig. 4C), we derived a leaky gut network centred on the microbial translocation marker, LPS, whose levels, as expected, were positively associated with pathogenic bacteria such as Escherichia (r = 0.4388) and Acinetobacter (r = 0.3478) as well as fecal levels of gut permeability marker zonulin (r = 0.3396). Additionally, Firmicutes and Bifidobacterium were positively associated with fecal acetate (r = 0.3624) and butyrate levels (r = 0.5492) respectively, both of which in turn positively correlated with sIgA (r = 0.4117, r = 0.4371) and negatively associated with fecal LPS (r = − 0.3801). Butyrate and sIgA levels also correlated negatively with translocated plasma LPS (r = − 0.4723, r = − 0.4587). Finally, and as expected, butyrate levels were also negatively correlated with fecal zonulin levels (r = − 0.4117) In summation, the observed leaky gut phenotype appeared to be dependent on the interplay of factors governing levels of fecal butyrate, acetate and sIgA. Consequently (Fig. 4D), we report another sub-network where pro-inflammatory mediators were positively correlated with pathogenic bacteria and LPS negatively correlated with taxa known to beneficially influence gut health. Further, IL-10, an anti-inflammatory cytokine was negatively correlated with both Enterobacteriaceae (r = − 0.3482) and LPS (r = − 0.3237).
Fig.4
Microbiome-Immune interactions in the pathogenesis of infection- The correlation network represents the pairwise Spearman correlation analysis of the obtained fecal bacterial products (Fig. 1), inflammatory markers (Fig. 2), gut integrity and bacterial translocation markers (Fig. 3) with each other and with relative abundance of bacterial taxa of all the study participants and relative frequencies of immune subsets in HCMV infected infants with NC reported in the previous study [14]. Red lines indicate a positive correlation and blue indicates a negative correlation. The strength of the correlation is indicated by the intensity of the color of the lines in the network. Only the significant correlations have been plotted. The coloured boxes indicate the following: Orange- Relative abundance of bacterial taxa; Pink- Relative frequencies of immune cell subsets; Green-Fecal microbial products; Yellow- Fecal inflammatory markers; Purple- Fecal Gut integrity markers; (A) Granzyme B expressing cells regulatory network; (B) T cell compartment regulatory network; (C) Leaky gut network; (D) Inflammatory cascade network
Correlation analysis was also carried out for the IgM amount expressed in terms of absolute absorbance for the HCMV infected groups with relative abundance of bacterial taxa reported in the previous study [14] and the fecal metabolites assessed in this study (Figs. 1, 2 and 3) (Supplementary Fig. 6). IgM levels (indicative of viral replication) and abundance of inflammation associated bacterial taxa (Gammaproteobacteria) (r = 0.623), microbial translocation markers (Plasma LPS) (r = 0.447) and dysregulated gut integrity markers (Zonulin) (r = 0.554) were positively correlated with each other. On the other hand, IgM levels negatively correlated with butyrate (r = − 0.4) and also with Bacteroides (r = − 0.421), Veillonella (r = -0.492), and also Bifidobacterium breve (r =− 0.42).
An integrated signature defining the spectrum of HCMV mediated pathology in NCIn an effort to determine the discriminatory potential of all the above reported data with respect to our study groups, principal component analysis (PCA) was employed. PCA involving all fecal analytes described above may be promising in delineating profiles, distinct from healthy controls and corresponding to varying degrees of pathology in infected individuals (PC1, 40.19%). (Fig. 5A) Cumulative analysis incorporating relative abundance of observed taxa obtained in the previous study [14] and fecal analytes reported in this study also resulted in clearly discriminatory profiles (Fig. 5B, PC1, 20.92%). Finally, we integrated systemic cellular immune subset signatures reported in the previous study[14], where available, into the PCA (Fig. 5C) within a subset of HCMV infected individuals to consolidate the discriminatory gut microbiome—immune axis at play which governs HCMV mediated pathology in the context of NC (PC1, 21.78%).
Fig.5
Principal component analysis based on; (A) Estimated fecal bacterial products, inflammatory and gut integrity markers of HCMV infected infants (N = 15 each) with NC and healthy controls (N = 10); (B) Estimated fecal bacterial products, inflammatory and gut integrity markers and relative abundance of bacterial taxa of HCMV infected infants with NC (N = 15 each) and healthy controls (N = 10); (C) Estimated fecal bacterial products, inflammatory and gut integrity markers, relative abundance of bacterial taxa and relative frequencies of immune cell subsets of HCMV infected infants with NC (N = 9 each)
留言 (0)