
記住我


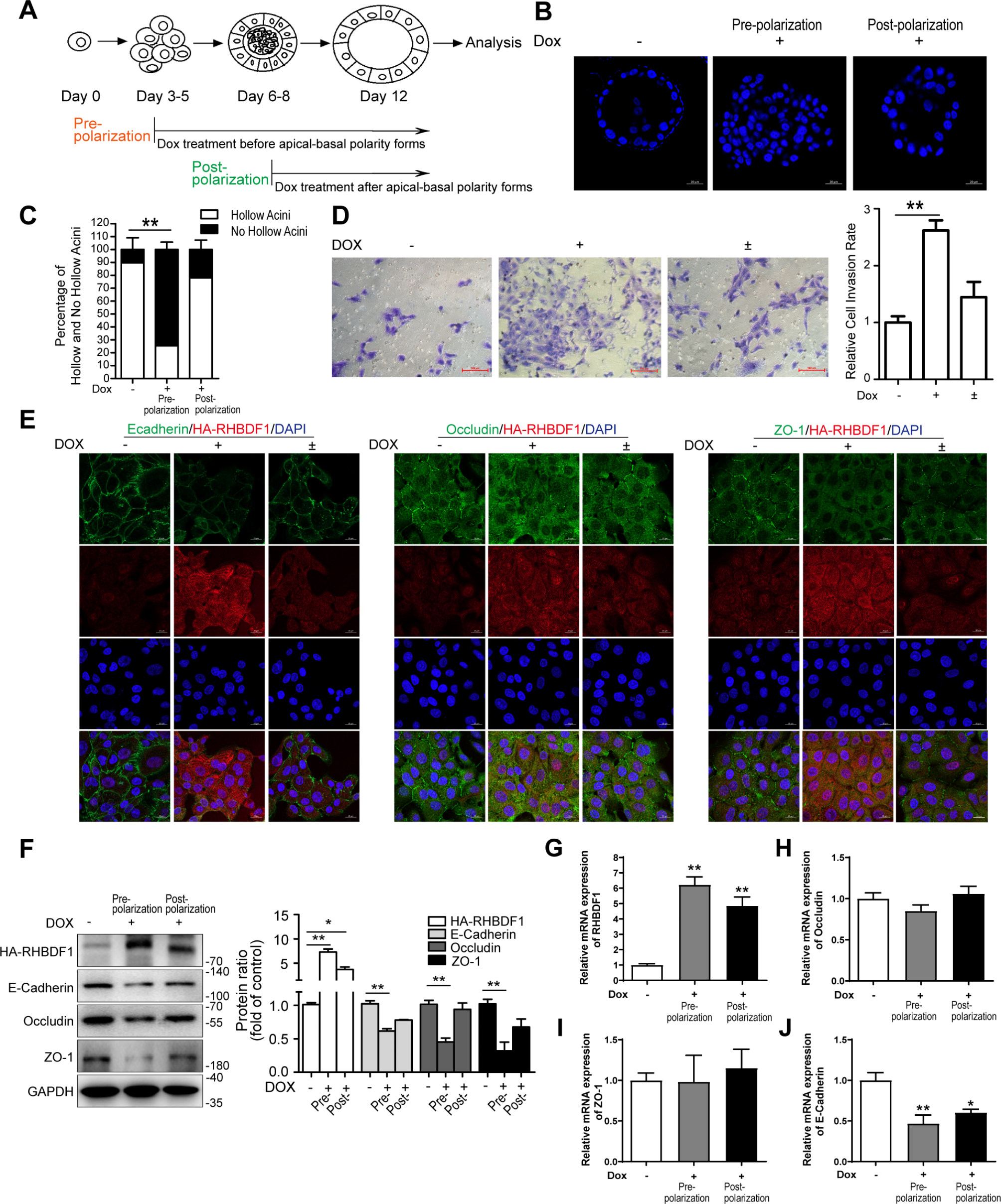

To explore the function of EZH1/2 in oocyte meiosis prophase I, we cultured mouse embryonic ovaries at 14.5 post-coitum (dpc) for 3 ~ 4 days to mimic this process in vitro. Initially, EZH1/2 function in embryonic ovaries was suppressed by EZH1/2 activity inhibitors. UNC1999 inhibits EZH2 and EZH1 histone-lysine N-methyltransferase activity through competitive inhibition of the cofactor S-Adenosyl-l-methionine (SAM). After treatment of UNC1999 for 3 days, H3K27me3 level in whole fetal ovaries was remarkably decreased compared to the vehicle control group (treated by an equivalent volume of DMSO) (Fig. 1A). The fluorescence intensity of H3K27me3 also showed a marked decrease in UNC1999 group (Fig. 1B, F). Similarly, EZH1 and EZH2 protein levels were reduced after UNC1999 treatment (Fig. 1A). These results indicate that EZH1/2 function was effectively suppressed by inhibitor UNC1999. DDX4 is a marker of germ cells and the number of DDX4-positive cells was clearly reduced in embryonic ovary after UNC1999 treatment (Fig. 1B, G). GSK343 is a highly potent and selective EZH2 inhibitor with an IC50 of 4 nM. GSK343 also suppressed the H3K27me3 and reduced the DDX4-positive cells in embryonic ovaries cultured for 3 days in vitro (Fig. 1C, H, I). These observations suggest that the inhibition of EZH1/2 leads to germ cell loss in embryonic ovary during meiosis prophase I.
Fig. 1
Inhibition of enzyme activation or gene knockout of Ezh1/2impairs the oocyte development in fetal mouse ovaries. (A) Protein levels of EZH1, EZH2, and H3K27me3 in whole fetal ovaries cultured with Ezh1/2 inhibitor UNC1999 checked by western blotting (WB). DMSO is a negative control of the inhibitor treatment (vehicle group). β-actin was used as an internal control. (B, C) Representative images of immunofluorescence (IF) staining of DDX4 and H3K27me3 in fetal ovaries treated with EZH1/2 inhibitor UNC1999 or GSK343 (green color). DDX4 (red color) is an oocyte marker. The nucleus was stained with DAPI (blue). Scale bar:50 μm; (D) Protein levels of EZH1, EZH2, and H3K27me3 in wildtype (WT) or Ezh1/2 double knockout (dKO) embryonic ovaries checked by WB. β-actin was used as an internal control. (E) The levels of H3K27me3 and DDX4 stained by IF in fetal mouse ovaries after knockout Ezh1/2. The red color indicates H3K27me3 and the green color is DDX4. The nucleus was stained with DAPI (blue). Scale bar: 50 μm; (F-K) Quantification of H3K27me3 fluorescence intensity or DDX4 positive cells in B, C, and E. (L-N) TUNEL assays on fetal ovaries with treatment of EZH1/2 inhibitors or dKO ovaries. Green is a positive color. The nucleus was stained with DAPI (blue). Scale bar: 50 μm; (O-R) Quantitative analysis of TUNEL positive cells in L-N. (R, S) The protein level of active caspase3 and caspase3 checked by WB in fetal ovaries treated with UNC1999 or deleted Ezh1/2 gene. GAPDH was used as an internal control. Vehicle: an equivalent volume of DMSO. WT: wild type; dKO: double knockout of both Ezh1 and Ezh2 in germ cells. Each group had 20 ovaries. The number of images assessed in a single experiment was at least 3.The experiments were repeated at least 3 times (N = 3). The data were presented as mean ± S.D. The asterisk (*) denotes a statistically significant difference between the vehicle (or WT) and treatment groups. NS: no significant, *P < 0.05, **P < 0.01 (t-test)
Secondly, the Ezh1/2 gene was specifically deleted in oocytes at the initiation of meiosis using a pLV-mAlpl-promoter-NLS-EGFP-Cre. The embryonic ovary at 14.5 dpc isolated from wildtype or Ezh1−/−, Ezh2flox/flox mice was infected with lentivirus expressed pLV-mAlpl-promoter-NLS-EGFP-Cre and cultured for 3 days. The protein levels of H3K27me3, EZH1 and EZH2 was significantly reduced in the double knockout (dKO) ovaries (Fig. 1D, E, J). The number of DDX4-positive cells was also significantly reduced in dKO ovaries (Fig. 1K), consistent with the observations following inhibitor treatment.
Then, cell apoptosis in fetal mouse ovaries was assessed using TUNEL assay. After treatment with UNC1999, cell apoptosis (TUNEL-positive cells) was increased in embryonic ovaries (Fig. 1L, O). But, the negative control chemical RSL3 (an inhibitor of glutathione peroxidase 4 (GPX4)) did not enhance cell apoptosis (Fig. 1M, P). The number of TUNEL-positive cells was also raised in dKO ovaries (Fig. 1N, Q). Active caspase 3 levels, a marker of apoptosis, were elevated in both UNC1999-treated and dKO ovaries (Fig. 1R, S).
These results demonstrate that EZH1/2inhibition or gene knockout enhances cell apoptosis and leads to loss of oocytes in embryonic ovaries during oocyte meiosis prophase I, suggesting that EZH1/2 plays an important role in this developmental stage of the embryonic mouse ovary.
EZH1/2-deficiency impedes meiotic progression in fetal mouse ovariesWe further investigated the impact of EZH1/2 on meiotic prophase I using meiosis-specific markers. Phosphorylated histone H2AX (γ-H2AX) marks double-strand breaks (DSBs) and is present from the leptotene to pachytene stages [24, 25]. Here, the γ-H2AX signal weaken in fetal ovary cultured with normal medium for 3 days (equaling 17.5 dpc), but remained strong in UNC1999-treated ovaries (Fig. 2A, C). To exactly observe the γ-H2AX in chromosome, nuclear spreading of fetal oocytes was performed. Results showed that few γ-H2AX foci in the vehicle group, but a substantial number of foci in oocytes from fetal ovary with UNC1999 or GSK343-treatment (Fig. 2B, E). In dKO ovaries, γ-H2AX foci also remained at a high level compared to wildtype ovaries (Fig. 2F). Statistical analysis showed that the number of γ-H2AX foci in oocytes at the late pachytene stage was maintained in fetal ovary with UNC1999 or GSK343-treatment or dKO ovaries (Fig. 2D, G, H). These results suggested that DNA damage repair was not completed in Ezh1/2-deficient oocyte.
Fig. 2
Defective Ezh1/2 function leads to failure of DNA break repair and disrupts oocyte meiosis progression in embryonic mouse ovaries. (A) Representative images of γ-H2AX with IF staining in fetal mouse ovaries treated with EZH1/2 inhibitor UNC1999. Red is a positive color. DDX4 is green. The nucleus was stained with DAPI (blue). Scale bar: 50 μm, enlarged scale bar: 10 μm; (C) Quantitative analysis of γ-H2AX fluorescence intensity (A). (B, E, F) Representative images γ-H2AX (red), SCP3 (red), and DAPI (blue) by IF staining in pachytene stage oocytes with chromosome spread. SCP3, the major cytoskeletal protein of the synapsis complex, is continuously expressed throughout premeiotic phase I and is a chromosomal linkage site. Here SCP3 staining is present in the displayed chromosome. Oocytes were isolated from fetal ovaries treated with EZH1/2 inhibitors (UNC1999 (B) and GSK343(E)) or dKO ovaries (F). Scale bar:2 μm; (D, G, H) Quantitative analysis of γ-H2AX foci in B, E, F. (I-L) Representative images of IF staining of MSY2 (a marker of diplotene phase of meiosis) in fetal ovaries with the treatment of EZH1/2 inhibitor (UNC1999 (I) or GSK343 (J)), negative control chemical RLS3 (K), and lentivirus (dKO, L). Green is a positive color. DDX4 staining indicated oocytes. H3K27me3 was stained as red color. The nucleus was stained with DAPI (blue). Scale bar: 50 μm, enlarged scale bar: 10 μm; (M-P) Quantitative analysis of MSY2 fluorescence intensity in I-L. (Q, R) Representative images of SCP1 IF staining in fetal ovaries treated with UNC1999 or dKO ovaries. SCP1 was stained as green color. DDX4 is red. The nucleus was stained with DAPI (blue). Scale bar: 50 μm, enlarged scale bar:10 μm; (S, T) Quantitative analysis of SCP1 fluorescence intensity (Q and R). The number of images assessed in a single experiment was at least 3. The experiments were repeated at least 3 times (N = 3). The data were presented as mean ± S.D. *P < 0.05, **P < 0.01 (t-test)
Abnormal DNA damage repair may interfere the meiotic prophase progression. Therefore, the effect of the Ezh1/2-deficient on meiotic prophase progression in oocytes was further explored. MSY2 is a marker of the diplotene stage [26]. When mouse fetal ovaries at 15.5 dpc were cultured for 3 days (equaling 18.5 dpc), MSY2 was expressed in oocytes from vehicle ovaries. However, MSY2 protein was largely absent in oocytes in ovaries with UNC1999 or GSK343 treatment (Fig. 2I, J). Statistical analysis confirmed that the MSY2 fluorescence intensity was significantly reduced in the UNC1999 or GSK343 ovaries (Fig. 2M, N). However, the negative control chemical RSL3 did not affect the expression of MSY2 in embryonic ovaries (Fig. 2K, O). After ovaries (15.5 dpc) were infected with lentivirus for 3 days in vitro, few oocytes in the dKO ovaries expressed MSY2 (Fig. 2L, P). These results imply that oocytes with Ezh1/2 deficiency are impeded in reaching the diplotene stage.
SCP1 is the major component of the synaptonemal complex (SC) transverse filament and disappears at the diplotene stage [27]. Here, we performed SCP1 immunofluorescence analysis in fetal mouse ovaries and found that SCP1 was lost in oocytes in vehicle or WT ovaries when 15.5 dpc ovaries were cultured for 3 days in vitro, while SCP1 expression persisted in the UNC1999-treated or dKO ovaries (Fig. 2I, J, K, L). The observation of SCP1 confirmed that oocytes did not enter the diplotene stage when EZH1/2 function was suppressed.
The above results show that most oocytes in EZH1/2-deficient ovaries fail to repair DNA damage, and thereby do not enter the diplotene stage, suggesting that EZH1/2 is required to ensure the normal process of meiotic prophase I in fetal oocytes.
Ezh1/2 deficiency causes abnormal expression of genes associated with DSBs repair initiation during meiosis prophase I in embryonic ovariesTo discover the mechanism by which Ezh1/2 deficiency impairs DSB repair, we assessed the function of proteins associated initiation of DSBs repair in meiotic prophase I. Ataxia-telangiectasia mutated proteins (ATM) is a serine/threonine protein kinase and play a key role in the initiation of DSBs repair. Immunofluorescence assays were performed to assess the levels of ATM and phospho-ATM (Ser1981) in mouse embryonic ovaries treated with UNC1999 for 3 days in vitro. The fluorescence intensity of ATM did not significantly change after the UNC1999 treatment (Fig. 3A, E). However, the fluorescence intensity of p-ATM was significantly decreased after treatment with UNC1999 or GSK343 (Fig. 3B, C, F, G). In dKO ovaries, the fluorescence intensity of p-ATM was also remarkably reduced (Fig. 3D, H). Meanwhile, p-ATM protein levels were dropped in dKO ovaries (Fig. 3I), and the ratio of p-ATM to ATM was clearly reduced (Fig. 3I). Hormad1, a meiosis-specific protein that promotes synapsis and recombination of homologous chromosomes in meiotic prophase, may affect the downstream phospho-ATM [28,29,30,31]. Western blotting and RT-qPCR analysis results showed that the protein and mRNA of Hormad1 was decreased in the UNC1999-treated or dKO ovaries (Fig. 4A, B, C, D).
Fig. 3
Ezh1/2 function deficiency suppresses the phosphorylation of ATM in mouse fetal mice. (A) Representative images of IF staining of ATM (red) and DDX4 (green) in fetal ovaries treated with EZH1/2 inhibitor (UNC1999). The nucleus was stained with DAPI (blue). Scale bar: 50 μm, enlarged scale bar: 10 μm; (B-D) The level of p-ATM in fetal ovaries treated with UNC1999 (B), GSK343(C), and lentivirus (D, dKO). Green is the positive color for p-ATM. DDX4 is stained red. The nucleus was stained with DAPI (blue). Scale bar: 50 μm, enlarged scale bar: 10 μm; (E) Quantitative analysis of ATM fluorescence intensity in A. (F-H) Quantitative analysis of p-ATM fluorescence intensity in B-D. (I) The level of p-ATM in WT or dKO ovaries checked by WB. GAPDH was used as an internal control. The ratio of p-ATM to ATM protein is shown in the right figure. Every group had 20 ovaries. The number of images assessed in a single experiment was at least 3. The experiments were repeated at least 3 times (N = 3). The data were presented as mean ± S.D. *P < 0.05, **P < 0.01 (t-test)
Fig. 4
Ezh1/2 function deficiency results in abnormal expression of genes involved in the initiation of DSBs. (A-B) The protein level of hormad1 in fetal ovaries treated with UNC1999 or lentivirus. GAPDH and ERK were used as an internal control. Quantitative analysis of Hormad1 protein level normalized by GAPDH protein. Every group had 20 ovaries. (C-J) The relative expression levels of Hormad1, Mre11, Rad50, and Nbs1 mRNA in fetal ovaries treated with UNC1999 or lentivirus (dKO) by RT-qPCR. Gapdh is a reference gene. The data were normalized with that WT at 14.5 dpc + 0 days. The experiments were repeated at least 3 times (N = 3). Data were presented as mean ± S.D. The asterisk (*) denotes a statistically significant difference between the vehicle (or WT) and treatment groups. *P < 0.05, **P < 0.01 (t-test)
ATM protein kinase is activated by DSBs through the Mre11-Rad50-Nbs1 (MRN) DNA repair complex and orchestrates signaling cascades that initiate the DSBs repair mechanism in meiosis [32]. When 14.5 dpc ovaries were cultured with UNC1999 or lentivirus for 1 or 2 days, the relative expression levels of Mre11, Rad50, and Nbs1 were not significantly altered (Fig. 4E, F, G, H, I, J, P > 0.05). However, after 3 days of culture in vitro, the expression of Mre11, Rad50, and Nbs1 was significantly decreased in UNC1999-treated or dKO ovaries.
These results suggest that EZH1/2 governs the initiation of DSBs repair by ATM-MRN complex.
Abnormal gene expression in fetal mouse ovaries following Ezh1/2 DepletionTo investigate the molecular consequences of Ezh1/2 deficiency in fetal mouse ovaries, we extracted mRNA from WT and dKO ovaries at 14.5 dpc + 3 days and conducted RNA sequencing. A total 19,070 genes (S1 Data) were analyzed, revealing 244 differentially expressed genes (DEGs) based on a standard |log2 (fold change) | > 0 and Q value (padj) < 0.05 (S2 Data). These DEGs included 148 up-regulated genes and 96 down-regulated genes (Fig. 5A). The expression patterns of these DEGs were visualized in a heat map (Fig. 5B, Supplemental file 1). These genes were categorized into several groups, including those involved in embryonic development, DNA methylation, histone modification, and DNA damage repair, etc. The relative expression levels of these genes were further validated by RT-qPCR. As depicted in Fig. 5C and D, we confirmed the up-regulation of genes associated embryonic development (Tle6, Pkd2, Ptk2b, Ren1, Sod3, Zp2, Zp3, Lrrc38, Kcnk18, Creg1, B4galt4, Gper1), DNA methylation (Aqp5, Pdpn, St3ga16, Paqr5, Chpf, Tm6sf1, Psen2, Abcg1), histone modification (Arc, Fabp7, S100g, Meox2, Mmp12, Muc1, Enpp2, Gprc5b, Ahcy), and DNA damage or DSBs repair (Gsta2, Foxq1, Tgfbr2, Ptgds, Gucy2g, Aldh1b1, Tpp1, Ctss, Lyz2, Tal2, Pak6, Rybp). Conversely, in Fig. 5E and F, we verified the downregulation of genes associated with embryonic development (Cwc22, I111ra2, Iqcb1, Lypd10, Tchh11, Tuba3b, Wdr54, Zfp990, Neur13, Spib, Myh4), DNA methylation (Ankrd34b, Ankrd37, Ppp1r3g, Pkp1, Aire, Phyhipl), histone modification (Hoxd1, Ndrg1, Rhox9, Prdm9, Stra8), and DNA repair or DSBs (Tex30, Prdm9, Stra8, Gm960, Iho1).
Fig. 5
Knockout of Ezh1/2 interferes with the expression of many functional genes in fetal mouse ovaries. Gene transcription in embryo ovaries were sequenced on an Illumina HiSeq 2500 with 125 bp single end sequencing and gene expression data were analyzed by using DAVID (the Database for Annotation, Visualization, and Integrated Discovery, [https://david.ncifcrf.gov]). (A) Volcano plot of differentially expressed genes in WT and dKO ovary tissues on 14.5dpc + 3 days as determined by RNA-seq. Blue or red dots represent genes that are differentially expressed (|log2FC|>=1, Qvalue < = 0.05) between WT and dKO group. Red dots indicate upregulated genes, and green dots indicate down-regulated genes. Genes not significantly differentially expression (no DEGs) are shown in gray. The FC in log2FC is fold change, which represents the ratio of the expression between the two samples (groups). Log10 (Expression of WT/dKO): the normalized expression value is taken as the log, usually using log10. (B) Heatmap of physiological function associated genes differentially regulated between WT and dKO ovary tissues on 14.5dpc + 3 days. Heatmap of the clustering of the different expressed genes. Green indicates lower expression, red indicates higher. FPKM of each gene from different samples was normalized to Z-score. (C-F) The relative expression level of genes in fetal ovaries treated with lentivirus by RT-qPCR. Gapdh is a reference gene. The values in each group were averaged and normalized with that of WT group. The experiments were repeated at least 3 times (N = 3). Data were presented as the mean ± S.D; The asterisk (*) denotes a statistically significant difference between the WT and dKO groups. *P < 0.05, **P < 0.01 (t-test)
These findings show that Ezh1/2 deficiency impacts gene expression in various processes during embryonic ovarian development, including germ cell development and oocyte meiosis in fetal mouse ovaries.
Impact of EZH1/2 Knockout on Histone Methylation in Fetal Mouse OvariesDynamic changes in histone modifications are essential for the regulation of double-strand break repair during meiotic prophase I in oocytes. Histones such as H3K4me3, H3K79me2/3, H3K9me3, and H4K20me2 play significant roles in the meiosis of oocytes. These histones are involved in the selection of the homologous recombination (HR) repair pathway by facilitating chromatin opening through methylation of H3K9 and H4K20 and recruitment of repair proteins. After infecting 14.5 dpc ovaries with lentivirus in vitro for 3 days, we observed decreased methylation levels of H3K9me3 and H4K20me2 in dKO ovaries (Fig. 6A, B). mRNA level of their target genes L3mbtl4 and Fbxo44 was also increased in dKO ovaries (Fig. 6C, D). These findings suggest that the knockout of Ezh1/2 affects the activity of other histones modification.
Fig. 6
Deletion of Ezh1/2 affects other histone proteins function in mouse embryonic ovaries. (A, B) The protein levels of H3K9me3 and H4K20me2 in embryonic ovaries checked by WB. Histone H3 was used as an internal control. Every group had 20 ovaries. (C, D) The relative expression levels of L3mbtl4 and Fbxo44 mRNA in fetal ovaries measured by RT-qPCR. Gapdh is a reference gene. The experiments were repeated at least 3 times (N = 3). Data were presented as the mean ± S.D. The asterisk (*) denotes a statistically significant difference between the WT and dKO groups. *P < 0.05, **P < 0.01 (t- test)
留言 (0)