
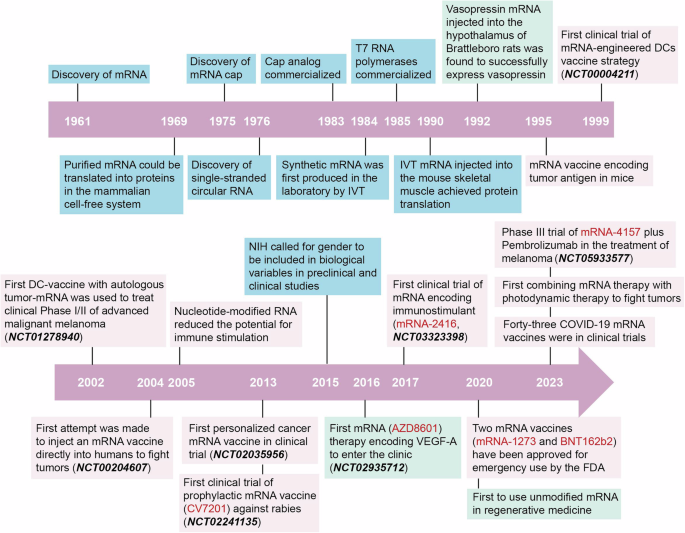
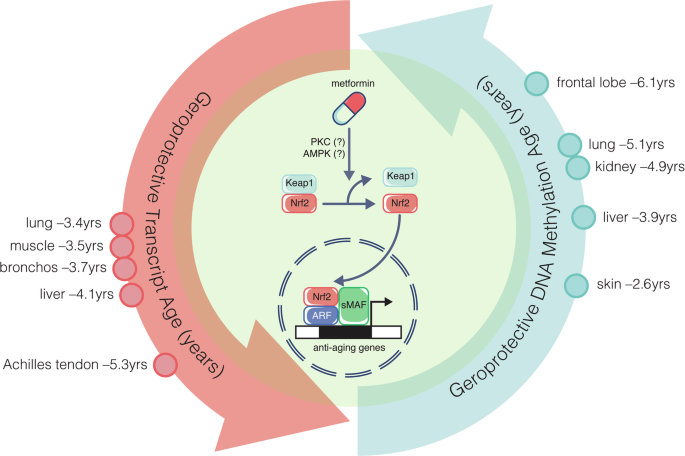
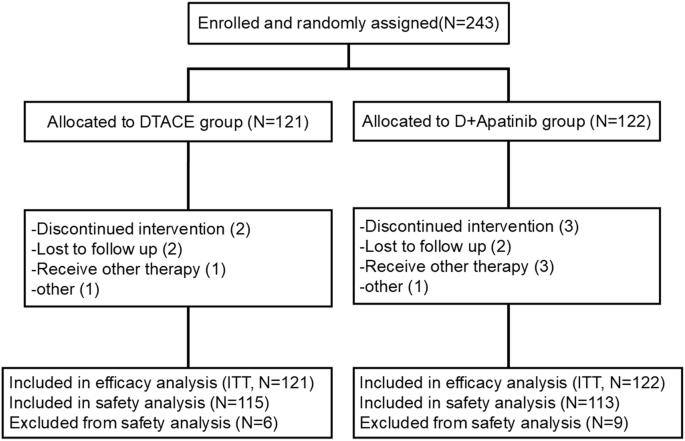
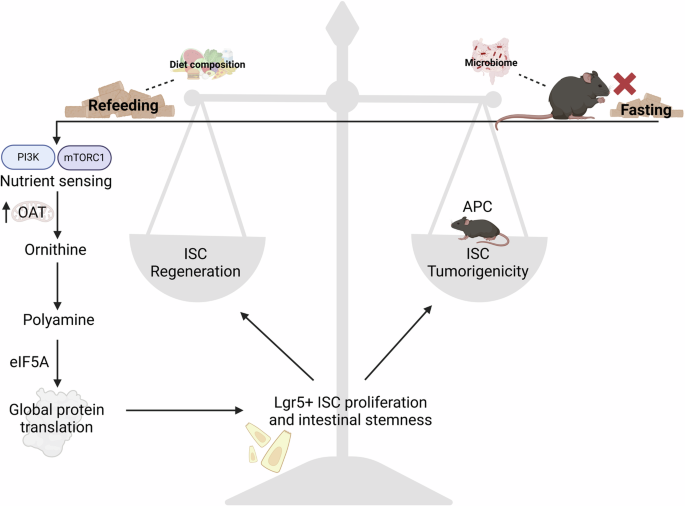
記住我
In order to explore the expression of STX11 in lung fibrosis, we screened the database related to IPF in GEO database. As shown in Supplementary Fig. 1a–f, the expression of STX11 was significantly decreased in IPF group compared with the control group (GSE10667, GSE24206, GSE32537, GSE53845, GSE72073, GSE110147). Even though no statistical significance was observed in GSE21369 and GSE35145 data sets, its expression in IPF group exhibited a decreasing trend (Supplementary Fig. 1g, h). Additionally, we performed a meta-analysis of STX11 expression data from GEO database. As shown in Supplementary Fig. 1i, STX11 expression was markedly lower in IPF group (SMD = −1.88, 95% CI: −2.59 to −1.18, p < 0.0001).
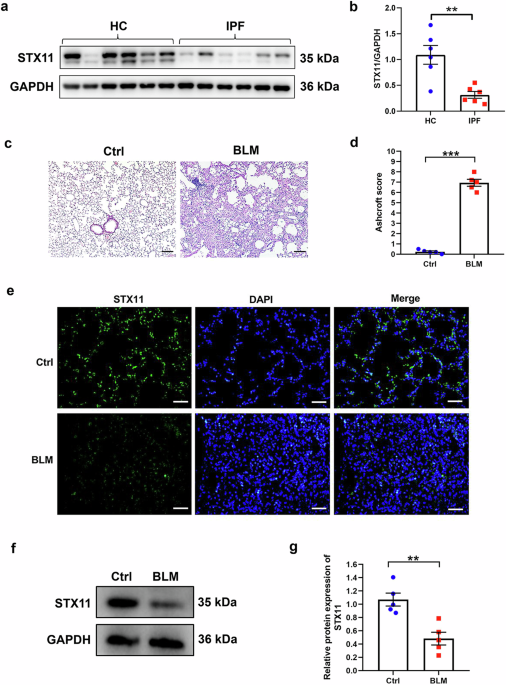
We further verified STX11 expression in clinical specimen and mice lung tissues. The western blot showed reduced STX11 expression in human IPF lung tissues as compared to health control (HC) lung tissues (Fig. 1a, b). Meanwhile, we intratracheally instilled BLM to induce lung fibrosis in mice. H&E results showed damaged alveolar structure, accumulated collagen, thickened interlobular septa, increased Ashcroft score in BLM group, indicating that BLM indeed induced lung fibrosis in mice (Fig. 1c, d). We then examined its expression in mice lung tissues. Decreased expression of STX11 was observed in BLM group (Fig. 1e–g). Altogether, these results demonstrated reduced expression of STX11 in lung fibrosis, suggesting its possible involvement in the progression of lung fibrosis.
Fig. 1
The expression of STX11 in IPF and control lung tissues from the clinical specimen and mice. a, b The protein expression of STX11 in human IPF tissues and in normal human lung tissues was detected by western blot assay. c H&E staining was conducted on mice lung tissue sections from BLM group and control group. d The Ashcroft score was used to evaluate the degree of fibrosis in mice lung tissues. Immunofluorescence (e) and western blot (f, g) assays were applied to detect the expression of STX11 in mice lung tissues (original magnification ×200, scale bar: 100 μm). Data were expressed as mean ± SEM (n = 5 or 6). *p < 0.05; **p < 0.01; ***p < 0.001
STX11 expression is decreased in TGF-β1-induced fibroblast activationFibroblast activation plays a key role in the pathogenesis of IPF. In order to establish a cell model of fibroblast activation, we treated human lung fibroblasts (HLFs) with TGF-β1. The qPCR and western blot results showed enhanced expression of ACTA2 (gene encoding α-SMA), fibronectin (gene encoding fibronectin), and COL1A1 (gene encoding collagen I) in TGF-β1 group as compared with control group (Supplementary Fig. 2a–e). These data confirmed activation of HLFs by TGF-β1.
We then identified STX11 expression in HLFs treated with TGF-β1. The mRNA and protein expression of STX11 was significantly decreased in TGF-β1 group (Fig. 2a–d), suggesting that HLF activation downregulated STX11 expression and STX11 might be involved in fibroblast activation.
Fig. 2
The role of STX11 in fibroblast autophagy and activation. HLFs were treated with 10 ng/ml TGF-β1 for 48 h. a qPCR assay was used to detect the mRNA expression of STX11. b, c Western blot assay was used to detect the protein expression of STX11. d Immunofluorescence assay was used to detect the protein expression of STX11 (original magnification ×200, scale bar: 100 μm). HLFs were infected with lentiviruses harboring STX11 and treated with TGF-β1 for 48 h. qPCR (e) and western blot (f, g) assays were used to detect the expression of STX11. HLFs with STX11 overexpression were treated with 10 ng/ml TGF-β1 for 48 h. The expression of α-SMA, fibronectin, and COL1A1 (collagen I) was examined by qPCR (h–j) and western blot (k, l) assays. m, n Transwell assay was used to determine the migration ability of HLFs (original magnification ×100, scale bar: 200 μm). o–r Western blot assay was used to determine the expression of fibroblast activation markers (α-SMA, fibronectin, collagen I) and autophagy-associated markers (LC3II/I, P62) in HLFs with STX11 overexpression. s–v HLFs with STX11 overexpression were stimulated with chloroquine (CQ) at a final concentration of 10 mM for 48 h. Western blot assay was used to examine the expression of fibroblast activation markers and autophagy-associated markers in HLFs. GAPDH was used as an internal control. Data were expressed as mean ± SEM (n = 3 or 5). *p < 0.05; **p < 0.01; ***p < 0.001; NS, no significance
Overexpression of STX11 attenuates fibroblast activation via promoting autophagyTo reveal what role STX11 plays in fibroblast activation, we stably overexpressed STX11 in HLFs with lentivirus (LV). The mRNA and protein expression levels of STX11 were markedly upregulated in LV-STX11 group compared with LV-NC group (Fig. 2e–g). Forced expression of STX11 dramatically decreased TGF-β1-induced expression of α-SMA, fibronectin, and collagen I (Fig. 2h–l). In addition, we further identified the role of STX11 in HLF activaiton with the use of recombinant human STX11 protein and found that it also inhibited TGF-β1-induced expression of these fibroblast activation markers, indicating that STX11 inhibited TGF-β1-induced HLF activation (Supplementary Fig. 3). Meanwhile, we investigated the role of STX11 in migration of HLFs. As illustrated in Fig. 2m, n, TGF-β1 promoted migration of HLFs. And STX11 inhibited TGF-β1-induced migration of HLFs. We also explored the role of STX11 in HLF proliferation. Ki-67 protein has been widely utilized as a cell prolifertaion marker for decades. Therefore, to reflect the proliferation of HLFs, we decteded Ki-67 expression using immunofluorescence assay. pcDNA-STX11 plasmids were employed to overexpress STX11 in HLFs. As shown in Supplementary Fig. 4, TGF-β1 promoted proliferation of HLFs; whereas STX11 suppressed TGF-β1-induced HLF proliferation. In order to explore whether STX11 inhibited HLF activation via promoting autophagy, we examined the autophagy-related markers (LC3II/I and p62) and fibroblast activation markers (α-SMA, fibronectin, and collagen I) of HLFs in the absence or presence of an autophagy inhibitor (chloroquine, CQ). As shown in Fig. 2o–r, overexpression of STX11 resulted in decreased protein expression of α-SMA, fibronectin, collagen I, and p62 as well as increased ratio of LC3II/LC3I. To further determine the role of STX11 in autophagy, we transfected tandem-tagged mCherry-GFP-LC3 adenovirus into HLFs to monitor the subcellular localization of LC3. As shown in Supplementary Fig. 5, autophagosomes were increased in pcDNA-STX11 group, indicating that STX11 overexpression promoted autophagy of HLFs.
CQ can increase the pH value of acidic lysosomes and inactivate acidic hydrolases in lysosomes, and then inhibit the fusion and degradation of autophagic lysosomes within cells. Therefore, cells treated with CQ lead to the accumulation of LC3 II/LC3 I and p62, indicating blockage of autophagy. As expected, CQ treatment increased the protein expression of p62 and LC3 II/LC3 I, indicating that CQ can inhibit autophagy in HLFs (Fig. 2s–u). In addition, CQ treatment promoted the protein expression of fibroblast activation markers (Fig. 2s–v). These findings demonstrated that STX11 inhibited fibroblast-to-myofibroblast differentiation via activating autophagy.
STX11 interacts with SNAP25We employed STRING database to explore the molecules that interact with STX11 and the analysis indicated that STX11 might have a relationship with SNAP25, STXBP6, UNC13D, and FHL2 (Fig. 3a). We detected the mRNA expression of these genes in STX11-overexpressed HLFs. As shown in Fig. 3b–d, STX11 promoted SNAP25 expression at both mRNA and protein levels. Next, we determined STX11 expression in HLFs with SNAP25 knockdown or overexpression. Interestingly, we found that upregulation of SNAP25 also promoted STX11 expression (Fig. 3e–g); conversely, downregulation of SNAP25 inhibited STX11 expression (Fig. 3h–j).
Fig. 3
The relationship between STX11 and SNAP25. a The STRING online tool was used to identify the interaction protein with STX11. qPCR (b) and western blot (c, d) assays were used to detect the expression of SNAP25. qPCR (e) and western blot (f, g) assays were utilized to examine the expression of STX11 in HLFs transfected with pcDNA-NC or pcDNA-SNAP25. qPCR (h) and western blot (i, j) assays were conducted to explore the expression of STX11 in HLFs transfected with si-NC or si-SNAP25. k Immunofluorescence assay was performed to detect the expression of STX11 and SNAP25 in HLFs (original magnification ×200, scale bar: 100 μm). l Representative images showing STX11 (green), SNAP25 (red) and DAPI (blue) in HC and IPF lung sections by immunofluorescence staining (scale bar = 100 μm). m Immunofluorescence assay was performed to detect the expression of STX11 and SNAP25 in mice lung tissues (original magnification ×200, scale bar: 100 μm). n Reciprocal immunoprecipitation of STX11 and SNAP25 in HLFs. GAPDH was used as an internal control. Data were expressed as mean ± SEM (n = 5). **p < 0.01; ***p < 0.001
We further investigated whether STX11 interacted with SNAP25. First, we simultaneously detected the expression of STX11 and SNAP25 in the mouse lung tissue and HLFs by immunofluorescence. We found co-localized fluorescence between STX11 and SNAP25 in the human lung tissue, mouse lung tissue and HLFs (Fig. 3k–m), indicating a potential interaction between STX11 and SNAP25. Then, we conducted co-IP assay to validate whether STX11 could bind to SNAP25. The results showed that STX11 protein or SNAP25 protein could pull down each other, suggesting that STX11 and SNAP25 had an interaction and formed a complex (Fig. 3n).
Overexpression of SNAP25 attenuates fibroblast activation and proliferationWe detected SNAP25 expression in the lung tissues of mice with BLM-induced lung fibrosis and TGF-β1-activated fibroblasts. Decreased expression of SNAP25 was found in BLM group (Fig. 4a–c). Likewise, its expression in TGF-β1-activated group was lower than that in the control group (Fig. 4d–g). These results demonstrated that SNAP25 might be invloved in lung fibrosis and fibroblast activation.
Fig. 4
The role of SNAP25 in fibroblast autophagy and activation. a, b Western blot assay was used to detect the protein expression of SNAP25 in the mice lung. c Immunofluorescence assay was used to detect the protein expression of SNAP25 in the mice lung (original magnification ×200, scale bar: 100 μm). HLFs were treated with 10 ng/ml TGF-β1 for 48 h. qPCR (d) and western blot (e, f) assays were used to detect the expression of SNAP25 in HLFs. g Immunofluorescence assay was used to detect the protein expression of SNAP25 (original magnification ×200, scale bar: 100 μm). HLFs were transfected with pcDNA-NC or pcDNA-SNAP25 for 48 h. qPCR (h) and western blot (i, j) assays were conducted to identify the expression of SNAP25. HLFs were transfected with pcDNA-NC or pcDNA-SNAP25 for 24 h, and then stimulated with TGF-β1 for 48 h. qPCR (k–m) and western blot (n, o) assays were performed to determine the expression of α-SMA, fibronectin, and collagen I. p, q Transwell assay was used to determine the migration ability of HLFs (original magnification ×100, scale bar: 200 μm). r–u Western blot assay was used to determine the expression of fibroblast activation markers (α-SMA, fibronectin, collagen I) and autophagy-associated markers (LC3II/I, P62) in HLFs with enhanced expression of SNAP25. v–y HLFs with SNAP25 overexpression were stimulated with chloroquine (CQ) at a final concentration of 10 mM for 48 h. Western blot assay was used to detect the expression of fibroblast activation markers and autophagy-associated markers in HLFs. GAPDH was used as an internal control. Data were expressed as mean ± SEM (n = 3 or 5). *p < 0.05; **p < 0.01 ; ***p < 0.001
To assess the role of SNAP25 in fibroblast activation, we transfected pcDNA-SNAP25 or SNAP25 siRNA into HLFs. PcDNA-SNAP25 remarkably promoted SNAP25 expression in HLFs (Fig. 4h-j). As displayed in Fig. 4k–o, the expression of fibroblast activation markers was significantly reduced in SNAP25-overexpressed HLFs. Meanwhile, SNAP25 overexpression inhibited migration of HLFs (Fig. 4p, q). We also explored the role of SNAP25 in HLF proliferation. As shown in Supplementary Fig. 6, TGF-β1 promoted proliferation of HLFs; whereas SNAP25 overexpression inhibited TGF-β1-induced HLF proliferation, demonstrating that overexpression of SNAP25 impeded fibroblast activation, proliferation and migration.
In contrast, downregulation of SNAP25 by siRNA (Supplementary Fig. 7a–c) increased expression of markers for fibroblast activation as well as promoted HLF migration (Supplementary Fig. 7d–j). These results demonstrated that SNAP25 silencing promoted fibroblast activation and migration.
To explore whether SNAP25 inhibited HLF activation via promoting autophagy, we detected autophagy-related markers and fibroblast activation markers of HLFs in the absence or presence of CQ. As shown in Fig. 4r–u, overexpression of SNAP25 resulted in decreased protein expression of α-SMA, fibronectin, collagen I, and p62 and increased LC3II/LC3I. To further determine the role of SNAP25 in autophagy, we transfected tandem-tagged mCherry-GFP-LC3 adenovirus into HLFs to monitor the subcellular localization of LC3. As shown in Supplementary Fig. 8, increased autophagosomes were seen in pcDNA-SNAP25 group, indicating that SNAP25 overexpression promoted autophagy of HLFs.
CQ treatment increased the protein expression of p62 and LC3 II/LC3 I, indicating the blockage of autophagy in HLFs (Fig. 4v–x). In addition, the protein expression of fibroblast activation markers was increased post CQ treatment (Fig. 4v–y). These findings demonstrated that SNAP25 inhibited fibroblast-to-myofibroblast differentiation via activating autophagy.
Forced expression of STX11 attenuates fibroblast activation via upregulating SNAP25To determine whether STX11 inhibited fibroblast activation via upregulating SNAP25, we treated HLFs with lentivirus-STX11 and SNAP25 siRNA. As expected, upregulation of STX11 inhibited the expression of fibroblast activation markers, whereas this process could be reversed by downregulation of SNAP25, suggesting that STX11 inhibited fibroblast activation by upregulating SNAP25 (Fig. 5a, b).
Fig. 5
The effect of STX11 and SNAP25 on the PI3K/AKT/mTOR pathway. a, b HLFs with STX11 overexpression were transfected with si-NC or si-SNAP25 for 24 h, and then stimulated with TGF-β1 for 48 h. Western blot assay was performed to determine the expression of α-SMA, fibronectin, and collagen I. c–f The expression of the PI3K/AKT/mTOR pathway related proteins was detected by western blot in HLFs with STX11 overexpression. g–j HLFs with forced expression of STX11 were pretreated with 100 ng/ml IGF-1 for 1 h, and exposed to TGF-β1 for 48 h. Western blot assay was utilized to detect the protein expression in HLFs. k–n The expression of the PI3K/AKT/mTOR pathway related proteins was detected by western blot in HLFs with SNAP25 overexpression. o–r HLFs with forced expression of SNAP25 were pretreated with 100 ng/ml IGF-1 for 1 h, and exposed to TGF-β1 for 48 h. Western blot assay was utilized to detect the protein expression in HLFs. s–v HLFs with SNAP25 silencing were pretreated with 10 μM LY294002 for 1 h, and exposed to TGF-β1 for 48 h. Western blot assay was utilized to detect the protein expression in HLFs. Data were expressed as mean ± SEM (n = 3 or 5). *p < 0.05; **p < 0.01 ; ***p < 0.001; NS, no significance
STX11-SNAP25 complex attenuates fibroblast activation via blocking the PI3K/AKT/mTOR signaling pathwayTo illustrate the mechanism underlying suppressed activation of HLFs by STX11-SNAP25 complex, we explored the relevant signaling pathways. The PI3K/AKT/mTOR pathway plays a vital part in autophagy.16 Hence, we explored whether it was involved in the inhibition of fibroblast activation by STX11 and SNAP25. Western blot showed that this pathway was activated by TGF-β1, whereas suppressed by the enhanced expression of STX11 (Fig. 5c–f). In order to further determine whether STX11 exerts its effect on fibroblast activation in a PI3K/AKT/mTOR signaling pathway-dependent way, we treated HLFs with IGF-1, a PI3K/AKT/mTOR activator. The alterations in fibroblast activation markers and relevant proteins of the PI3K/AKT/mTOR pathway caused by STX11 overexpression could be reversed by IGF-1 (Fig. 5g–j). Collectively, these results suggested that STX11 inhibited fibroblast activation by suppressing this pathway.
We also examined the relationship between SNAP25 and the PI3K/AKT/mTOR pathway. As expected, forced expression of SNAP25 inhibited this pathway (Fig. 5k–n). However, the decreased expression of phospho-AKT, phospho-mTOR, a-SMA, fibronectin, and collagen I by forced expression of SNAP25 was elevated again after treating with IGF-1 (Fig. 5o–r). In contrast, downregulation of SNAP25 activated the pathway, which was reversed by the pathway inhibitor LY294002 (Fig. 5s–v). Taken together, these data indicated that SNAP25 suppressed fibroblast activation by blocking the PI3K/AKT/mTOR signaling pathway.
Enhanced expression of STX11 attenuates BLM-induced pulmonary fibrosis in miceTo explore the role of STX11 in BLM-induced lung fibrosis, we intratracheally instilled AAV-STX11 into the mice lung. H&E and Masson staining revealed that lung fibrosis developed in the BLM group (Fig. 6a, b). After AAV-STX11 instillation, STX11 expression was obviously elevated, which accompanied with decrease of lung fibrosis (Fig. 6a–c, g–h). Moreover, the expression of those fibroblast activation-related markers was increased in BLM and BLM + AAV-NC group as compared with Ctrl group (Fig. 6d–f, g, i–k). Whereas, the above-mentioned markers were reduced in BLM + AAV-STX11 group compared with BLM + AAV-NC group (Fig. 6d–f, g, i–k). These results indicated that overexpression of STX11 alleviated BLM-induced lung fibrosis in mice.
Fig. 6
Overexpression of STX11 inhibits BLM-induced pulmonary fibrosis in mice. a Representative images of H&E and Masson staining in the lung sections of mice. b Ashcroft score based on H&E staining was determined. c–f The mRNA expression of STX11, α-SMA, fibronectin, and COL1A1 was detected by qPCR. g–k The protein expression of STX11, α-SMA, fibronectin, and collagen I was detected by western blot. GAPDH was used as an internal control. Data were expressed as mean ± SEM (n = 8). *p < 0.05; **p < 0.01; *** p < 0.001; NS, no significance
留言 (0)