
記住我
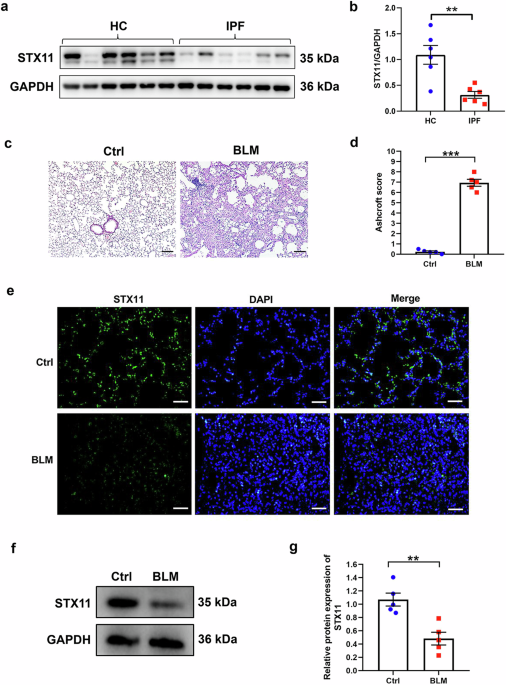
The B-cell linear antibody epitopes of the RBD proteins of some variants were analysed using IEDB.18 We found that the overall antigenic peptides of each RBD region appeared to be relatively similar. However, the antigenicity of some peptides seems to be different, indicating that some amino acid mutations may change the antigenicity of the whole peptide. For example, omicron variants (BA.1, BA.2, BA.3, and BA.4/5) showed an overall decrease in the antigenicity of RBD tail peptides (Supplementary Fig. 1). The key amino acid mutations in the RBD often cause immune escape of the virus. Mutations at E484 and F456 lead to a loss of neutralizing potency of sera from infected or immunized individuals. Moreover, mutations in G339, S371, K417, F486, L452, Q493, N501, and other sites may also reduce the antibody neutralization capacity. In addition, mutations in some of these sites directly reduce the receptor binding capacity of the RBD.19,20,21,22 At the same time, some conserved peptides are also critical for the ability of RBD to induce neutralizing antibodies.23 Here RBD sequences were designed according to the immune escape characteristics of each mutant RBD region and the antigenic characteristics of the RBD region of S protein (Fig. 1a). The antigen prediction results showed that the designed constructs did not change the overall antigen characteristics of the RBD (Fig. 1b–d).
Fig. 1
Design and preparation of the cRBD vaccine. a Schematic diagram of cRBD sequence design (adapted from “Multiple Sequence Alignment (Protein)”, by BioRender.com (2024). Retrieved from https://app.biorender.com/biorender-templates). The RBD region (319-541) on the S protein of the prototype strain was selected and specific mutation sites were added to obtain cRBD1-3. b–d Results of B-cell linear antigenic epitopes prediction for cRBD1-3. e SDS‒PAGE and Western-blot results of cRBD and BA.1 RBD proteins expressed by E. coli. The BA.2 RBD protein was a positive control in the Western-blot assay. As a eukaryote-expressed protein, it migrates as an ~36.55 kDa band in SDS‒PAGE under reducing conditions due to glycosylation. f–i Surface and cartoon representation of the interaction of hACE2 with BA.1 RBD and cRBD1-3. hACE2 is displayed in green, and cRBD or BA.1 RBD in orange. Surface plasmon resonance recorded the profile of a real-time affinity of hACE2 to BA.1 RBD and cRBD1-3
It has been reported that RBD protein obtained using prokaryotic expression system can induce an immune response after immunization in animals.24 Thus, we use the prokaryotic expression system to obtain the protein (the detailed steps are described in the “Methods” section). Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS‒PAGE) of the recombinant RBD proteins revealed a single band with a molecular weight of approximately 30 kDa (Fig. 1e), which is consistent with the theoretical value, indicating protein integrity and purity. Western-Blots verified that the purified proteins were indeed RBD proteins (Fig. 1e). Further, the structure of BA.1 RBD and cRBD proteins as well as their binding sites to human ACE2 (hACE2) were predicted using AlphaFold3 (Fig. 1f–i), and the results showed that the binding sites of the modified RBD protein with ACE2 have changed (Supplementary Fig. 2). Using surface plasmon resonance (SPR), we confirmed the binding and observed effective affinity between them (Fig. 1f–i). This represents that the proteins we obtained using the prokaryotic expression system have the correct conformation as well as biological activities. Moreover, the results of SPR showed that hACE2 had the highest affinity for BA.1 RBD and the lowest for cRBD3. The changes in the binding site and affinity of the modified RBD proteins for ACE2 might be responsible for the changes in the immune responses they induced. The adjuvant AddaS03 was mixed with protein to prepare a vaccine for subsequent experiments.
cRBDs induce strong immune responses in BALB/c miceAs shown in Fig. 2a, a three-injection strategy was used with a 21-day interval between each immunization, and for each injection, two different doses, namely, a low dose (5 μg/dose) and a high dose (25 µg/dose), were applied. To confirm whether the immune response could be activated after vaccination, we measured the expression levels of cytokines and chemokines in the serum at 6 h and 24 h (Fig. 2b). In addition to establishing the vaccine-immunized group, we also established an adjuvant group and a buffer group as control groups. Compared with those in the buffer group, the levels of many cytokines in the adjuvant and vaccine groups increased more obviously at 6 h post-injection than at 24 h post-injection. For example, IL-5, which is a critical cytokine for B-cell differentiation to antibody-secreting plasma cells in mice,25 and IL-6, which is important for B-cell proliferation and isotype switching.26 Furthermore, the production of MCP-1, MIP-1a, MIP-1b, and TNF-α, which are key chemokines for antigen-presenting cell activation and migration, was also induced at 6 h compared with 24 h.26,27 No such change was found in the citrate buffer injection group. These findings suggest that the vaccine induces an immune response and that this response is partly induced by the adjuvant.
Fig. 2
cRBDs induced strong immune responses in BALB/c mice. a The timeline of vaccine immunization and sampling (created with BioRender.com). The serum used for cytokine detection was collected at 6 h and 24 h after the first injection. Final blood collection was performed at 14 and 21 days (Days 14 (n = 3), 21 (n = 3), 35 (n = 5), 42 (n = 5), 56 (n = 5), and 63 (n = 5) in (a)) after each immunization to collect serum for detection of vaccine-induced production of binding and neutralizing antibodies. Spleens were collected simultaneously 21 days after the third injection (day 63) for an ELISPOT assay. b Fold change in the levels of inflammation-related cytokines in the serum at 6 h and 24 h after immunization (n = 5). The color of the circle represents the P-value. The smaller the P value is, the redder the color. The size of the circle represents the fold change (FC). FC = cytokine level at 6 h after immunization /cytokine level at 24 h after immunization. The greater the FC is, the larger the circle. c–f BA.2 RBD-specific binding antibody levels at each time point after immunization in each group. Data are presented as mean ± SD
Considering that most of the currently common variants evolved from BA.2, we measured BA.2 RBD-binding antibody levels on days 14 and 21 after each injection. On day 14 after the first immunization, BA.2 RBD-binding antibodies were detected in all groups except the cRBD2 low-dose and BA.1 RBD low-dose groups, and they were detected in all groups on day 21. Moreover, there were no differences between the high- and low-dose in cRBD1 and cRBD3 groups, while the cRBD2 and BA.1 RBD groups showed some differences only before the second immunization (higher levels of binding antibodies in the high-dose group). As the immunization process progressed, there was an overall trend toward increasing binding antibody levels. Antibody titers at the end of the immunization process (63 d) were ~1 × 105 in all groups, which indicates that all proteins have good immunogenicity.
cBRD3 elicits the production of antibodies with broadly neutralizing activity against SARS-CoV-2 variantsIt has been reported that higher levels of nAbs are associated with lower chances of immune evasion.28 Therefore, nAbs titers against different SARS-CoV-2 variants are key indicators for assessing the breadth of the protective capacity of a vaccine. We investigated whether the designed proteins could elicit a broadly neutralizing antibody response against a wide range of variants by authentic and pseudoviral neutralization experiments and with BA.1 RBD as a control. The immunization procedure was the same as that described above (Fig. 2a), but blood was collected only on days 35, 42, 56, and 63. We detected the variants that appeared before the sequence design, including the prototype, alpha, beta, delta, BA.1, BA.2, and BA.5, and the strains that appeared after the sequence design, including XBB.1.15, XBB.1.16, EG.5.1, and EG.5.1.1 (Supplementary Fig. 3, Fig. 3a–c). In addition, some strains lacking available authentic viruses were complemented by pseudoviruses, including BF.7, CH.1.1, BQ.1.1, BA.2.86, and JN.1 (Supplementary Fig. 4, Fig. 3d–f).
Fig. 3
Neutralization activity of cRBD-induced antibodies against each strain. Geometric mean titer (GMT) of serum nAbs for authentic virus (a–c) (n = 5) and pseudovirus (d–f) (n = 3) 21 days after the third immunization (day 63 in Fig. 2a) for each group. Specific GMT values are presented in Supplementary Figs. 3–4. For ease of visualization, the cRBD1-3 results are plotted separately, and the results for the BA.1 RBD group are added as a control in each plot
Consistent with the results of the binding antibody assay, the level of nAbs against each strain increased as the immunization period progressed. cRBD1 induced a more rapid and stronger immune response, inducing the production of higher levels of nAbs at 14 days after the first immunization than in the other groups, regardless of the strain (Supplementary Fig. 3). Serum nAbs levels in mice immunized with BA.1 RBD were much lower than those in the mice immunized with the cRBD proteins, even that against BA.1 (Fig. 3a–c). nAbs against all tested strains were detected in cRBD-immunized mouse sera, even against the recently emerged BA.2.86 and JN.1 strains. However, some variants, such as CH.1.1, XBB.1.15, XBB.1.16, EG.5.1, and BA.2.86, were significantly less sensitive to neutralization by immunizing serum (Fig. 3a–c). Moreover, the levels of nAbs against variants were different among the three experimental groups. The neutralizing ability of the nAbs whose production was induced in response to each of the three constructs against the JN.1 variant was similar, but compared to the other groups, the cRBD3 high-dose group showed superior neutralizing activity against the BA.2.86 variant. Serum from cRBD2-immunized mice showed a more pronounced decrease in neutralizing ability against the XBB.1.5, XBB.1.16, EG.5.1, and EG.5.1.1 variants. Compared with serum from mice immunized with cRBD1, serum from mice immunized with cRBD3 had a slightly reduced ability to neutralize variants that emerged before the construct was designed, but its ability to neutralize new variants that emerged after the construct was designed was greater than that of serum from mice immunized with cRBD1. In general, cRBD3 induces a broader neutralization response. In addition, levels of nAbs against multiple variants could still be detected in the serum of mice 18 weeks or longer after completion of immunization, and antibody levels were relatively higher in the cRBD-immunized group, suggesting that cRBD-based vaccine induces long-lasting protection in mice after immunization (Supplementary Fig. 5).
To explore the ability of cRBD to induce cellular immunity, we collected the spleens of mice that had completed the full immunization schedule and isolated the lymphocytes. ELISPOTs were performed using the S protein of the BA.1.1, XBB, and EG.5 variants as stimulants (Fig. 4, Supplementary Fig. 6-8). Compared to the adjuvant control group, lymphocytes from mice immunized with either BA.1 RBD or cRBD proteins showed higher IFN-γ, IL-2, and IL-4 secretion than those from unimmunized or adjuvant-immunized animals. It suggests that the vaccine-immunized group induced specific T-cell responses to S proteins of BA.1.1, XBB, and EG.5. However, there was no significant difference between the BA.1 RBD- and cRBD-immunized groups, suggesting similar cellular immunity. The results showed that cRBD immunization could induce specific cellular immunity against XBB, EG.5, and BA.1.1 but not to a greater extent than BA.1 RBD.
Fig. 4
Statistical plots of the ELISPOT results. Three stimulants (BA.1.1, XBB, and EG.5 S protein) were used for detection. a–c The number of IFN-γ secreting cells in each group after stimulated (n = 3 in Control and AddaS03 group, n = 5 in other vaccinated groups). d–f The number of IL-2 secreting cells in each group after stimulated (n = 3 in Control and AddaS03 group, n = 3/4/5 in other vaccinated groups. g–i The number of IL-4 secreting cells in each groups after stimulated (n = 3 in Control and AddaS03 group, n = 3/5/5 in other vaccinated groups). The data are expressed as means ± SD. Statistical analysis was conducted using one-way ANOVA and Tukey’s multiple comparison test. The significance of differences between the experimental group and the unimmunized control group is indicated by black asterisks, the significance of differences between the experimental group and the adjuvant group is indicated by red asterisks, and no marker indicates no significance. *P < 0.05, **P < 0.01, ***P < 0.005
cRBDs protect BALB/c mice from challenge with live SARS-CoV-2Mice were immunized three times on days 0, 21, and 42, infected with SARS-CoV-2 on day 63, and dissected 5 days after challenge. Nasopharyngeal swabs and body temperature and weight data were collected daily during infection (Fig. 5a). After challenge with the EG.5.1 strain, the weights of the mice in all the groups tended to decrease, and the weight loss was the lowest in the cRBD3 high- and low-dose groups. Weight loss was most obvious in the control and adjuvant groups, except for the cRBD2 group (Fig. 5b). The nasopharyngeal swab loads of the control group and the vaccine group fluctuated greatly with time, but the swab load of some vaccine groups was still lower than that of the control group (Supplementary Fig. 9b, c). Moreover, the cRBD vaccine candidates significantly reduced the viral loads in the lungs of the infected mice (P < 0.05) (Fig. 5c). The pathological changes in the lungs of mice infected with the virus mainly manifested as pulmonary hemorrhage, inflammatory cell infiltration, vascular thrombosis, bronchial obstruction, and protein exudation (Fig. 5d, e). The degree of lung injury in the vaccine group was significantly lower than that in the control group, and the cRBD3 vaccine yielded the most robust lung protection (Fig. 5d, e). Although the BA.1 RBD was similar to the cRBDs in terms of ability to reduce the viral load, it was associated with worse lung pathology (Fig. 5d, e).
Fig. 5
Results of EG.5.1 variant challenge in Balb/c mice after immunization. a Schematic illustration of the timeline of BALB/c mice immunization and challenge with SARS-CoV-2 (created with BioRender.com). Mice that were not immunized were used as controls. The challenge experiment was performed 21 days after the third immunization. b Body weight changes after infection in each group (n = 10). c Viral gRNA levels in the lung at 5 days post-infection (dpi) in each group (n = 5). d Histogram of mouse lung pathology scores in each group (n = 5). e Histopathological examination of mice lung tissues. Scale bar, 100 μm. The data are expressed as means ± SD. Results of each group were compared with the control group in c and d. Statistical analysis was conducted using one-way ANOVA and Tukey’s multiple comparison test for bar graphs. ***P < 0.005. ns not significant
We also challenged the mice with the BA.5 variant after administering the cRBD1 vaccine and challenged mice immunized with the cRBD2 and cRBD3 vaccines with the XBB.1.5 variant (Supplementary Figs. 10–12). In the two challenge experiments, cRBD also showed great protective efficacy. The serum of mice immunized with cRBD showed low neutralizing activity against the strains that used in the challenge test (Fig. 3), and the level of neutralizing antibodies was positively correlated with the protective efficacy of the vaccine.28 Therefore, it can be speculated that the cRBD-based vaccine has a protective effect on all variants involved in neutralization test. The cRBD3 vaccine showed a broad-spectrum inhibitory effect against XBB.1.5 and EG.5.1 virus infection.
cRBDs protect K18-hACE2 mice from challenge with SARS-CoV-2In order to more realistically evaluate the protective effect of the cRBD vaccine, we evaluated the protective efficacy of the cRBD-based vaccine against SARS-CoV-2 using the K18-hACE2 model mice.29 As in the previous procedure, mice were immunized three times, and two weeks later the EG.5.1 attack experiment was performed (Fig. 6a). We detected similar levels of BA.2 RBD-binding antibodies in K18-hACE2 mice as in Balb/c mice during immunization, but lower levels of nAb and memory immune cells were induced in K18-hACE2 mice after vaccine immunization (Supplementary Fig. 13a–e). We measured the body temperature and body weight as well as the viral load of nasopharyngeal swabs of each group of mice during infection period, but no significant differences were observed between groups. (Supplementary Fig. 13f–h).
Fig. 6
Results of EG.5.1 variant challenge in K18-hACE2 mice after immunization. a Schematic illustration of the timeline of BALB/c mice immunization and challenge with SARS-CoV-2 (created with BioRender.com). b Changes in survival rate of mice in each group within 5 days of infection. c Body weight changes after infection in each group (n = 3). d Viral gRNA levels in the lung at 5 dpi in each group (n = 3). e Histogram of mouse lung pathology scores in each group (n = 3). f Histopathological examination of mice lung tissues. AddaS03 as adjuvant control. The data are expressed as means ± SD. Results of each group were compared with the AddaS03 group in d and e. Statistical analysis was conducted using one-way ANOVA and Tukey’s multiple comparison test for bar graphs. *P < 0.05, **P < 0.01, ***P < 0.005
Mice were dissected 5 days after infection. Death occurred in some immunized groups during infection, with mice in the cRBD1 low-dose group having the lowest survival rate at the end of the attack (Fig. 6b). The body weight of mice in all groups showed a decreasing trend after infection, with the greatest decrease in the adjuvant group and the lowest decrease in the cRBD3 high-dose group (Fig. 6c). In addition, cRBD-based vaccine immunization significantly reduced the viral load in the lungs of mice, with a significant reduction in viral load observed in the cRBD3 low-dose immunization group, suggesting that it exerted the most effective effect in up-suppressing viral replication (Fig. 6d). In addition, vaccine immunization significantly reduced lung pathological damage in all groups of mice, with the most significant reduction in lung pathological damage in the cRBD3-immunized group, suggesting that cRBD3 may play the most potent lung-protecting role in the challenge experiments (Fig. 6e, f). Therefore, we can speculate that the vaccine based on cRBD3 protein may exert the most obvious protective effect in the EG.5.1 challenge experiment.
Differences in the B-cell receptor repertoire induced by cRBDsTo further explore the reasons for the broad-spectrum differences after vaccination from the molecular aspect, we collected whole blood of mice in the cRBD high-dose group 21 days after completing three immunizations for transcriptome and B-cell receptor (BCR) sequencing. Another set of untreated mice was used as a negative control (NC) group.
In terms of the transcriptome, Gene Ontology (GO) enrichment of the differentially expressed genes (DEGs) (|log2FC| > 2, P < 0.05) was performed between the cRBD groups and NC group, and each group exhibited enrichment for immune- or inflammation-related pathways (Supplementary Fig. 14a–c). This finding suggested that immunization with these vaccines affects the immune system. Using the Reactome database,30 we identified genes related to innate and adaptive immunity pathways and analysed their expression among the groups. The expression of most of the IFN-γ pathway genes increased in the cRBD1 immunization group, followed by the cRBD3 immunization group, while the expression of most of the IFN-γ pathway genes tended to decrease in the cRBD2 immunization group (Supplementary Fig. 14d). The expression of genes complexed with MHCI and MHCII, which promote antigen presentation to CD4 and CD8 molecules, were also examined in each experimental group. CD8+T cells mainly participate in cellular immunity, while CD4+T cells mainly promote the proliferation and differentiation of B cells and participate in humoral immunity. Similar to that of IFN-γ, most of the gene expression in the cRBD1 group was induced (Supplementary Fig. 14e, f). This finding suggested that cRBD1 immunization may induce stronger humoral and cellular immunity. In our previous experiments, we confirmed that cRBD1 induces faster and stronger humoral immunity after immunization, as determined by serial detection of nAbs, and that higher levels of antibodies can be detected as early as 14 days after a single immunization (Supplementary Fig. 3).
For BCR sequencing, we first compared the abundances of different immunoglobulin types in the NC and vaccine-immunized groups, and the abundances of IgM and IgD were greater in all groups (Fig. 7a). Moreover, the diversity of IgM and IgD was much greater than that of the other immunoglobulins (Fig. 7b). Therefore, we focused on IgD and IgM and further assessed the use of V-J genes in the IgD and IgM heavy chain proteins of each group. The abundance of the top 10 V-J gene pairs with the highest frequency of use in each group for the IgM and IgD isoforms is shown in Fig. 7c and, e. Among the IgM subtypes, v2-2_J4, v2-9_J4, v2-9_J3, and v1-4_J4 showed abnormally high abundances in the cRBD3-immunized group (Fig. 7c). Further statistical analysis revealed that among the top ten most abundant V-J pairs, those in the cRBD3 group had significantly greater abundances than did those in the other groups (Fig. 7d). Similarly, for IgD, we observed the same difference in V-J usage (Fig. 7f). We hypothesized that this change in antibody abundance may be the reason why cRBD3 induced a better broad-spectrum response.
Fig. 7
Analysis of BCR sequencing results. a Cumulative histogram of the proportion of each isoform of IGH in each group. b Statistical plot of the diversity of IGH isoforms in each group, using D50 as an indicator, with larger D50 values indicating greater diversity of IGH in the samples. c Expression levels of top-10 used V-J pairs in IgM. d Statistical analysis of the expression levels of the top 10 V-J genes associated with IgM in each group. e Expression levels of the top 10 V-J pairs used for IgD. f Statistical analysis of the expression levels of the top 10 V-J genes associated with IgD in each group. Statistical analysis was conducted using multiple tests and corrected for FDR, with Scheffe post hoc tests. *P < 0.05, **P < 0.01, ***P < 0.005. ns no significant. n = 3 in each group
留言 (0)