
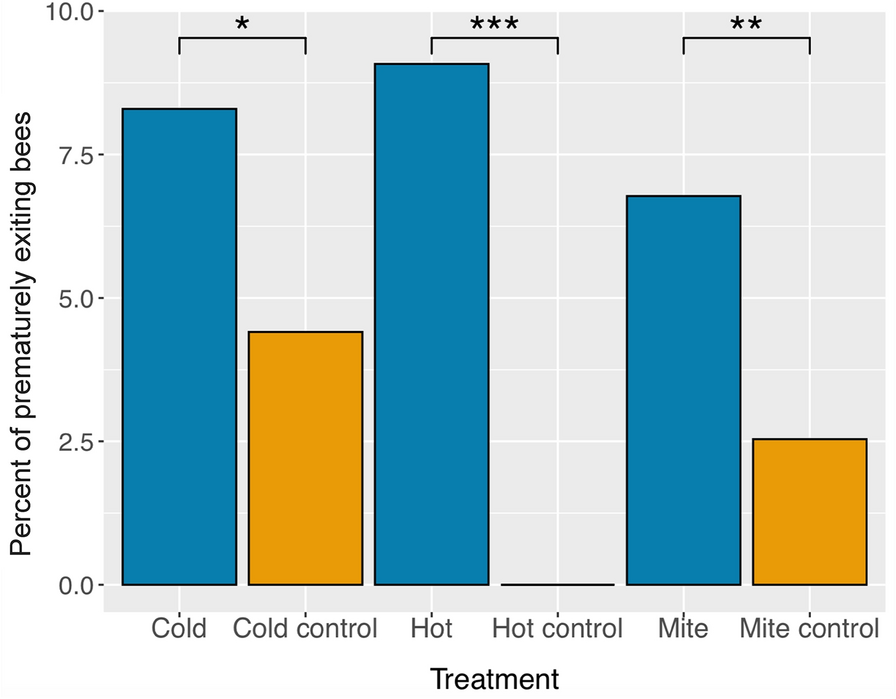
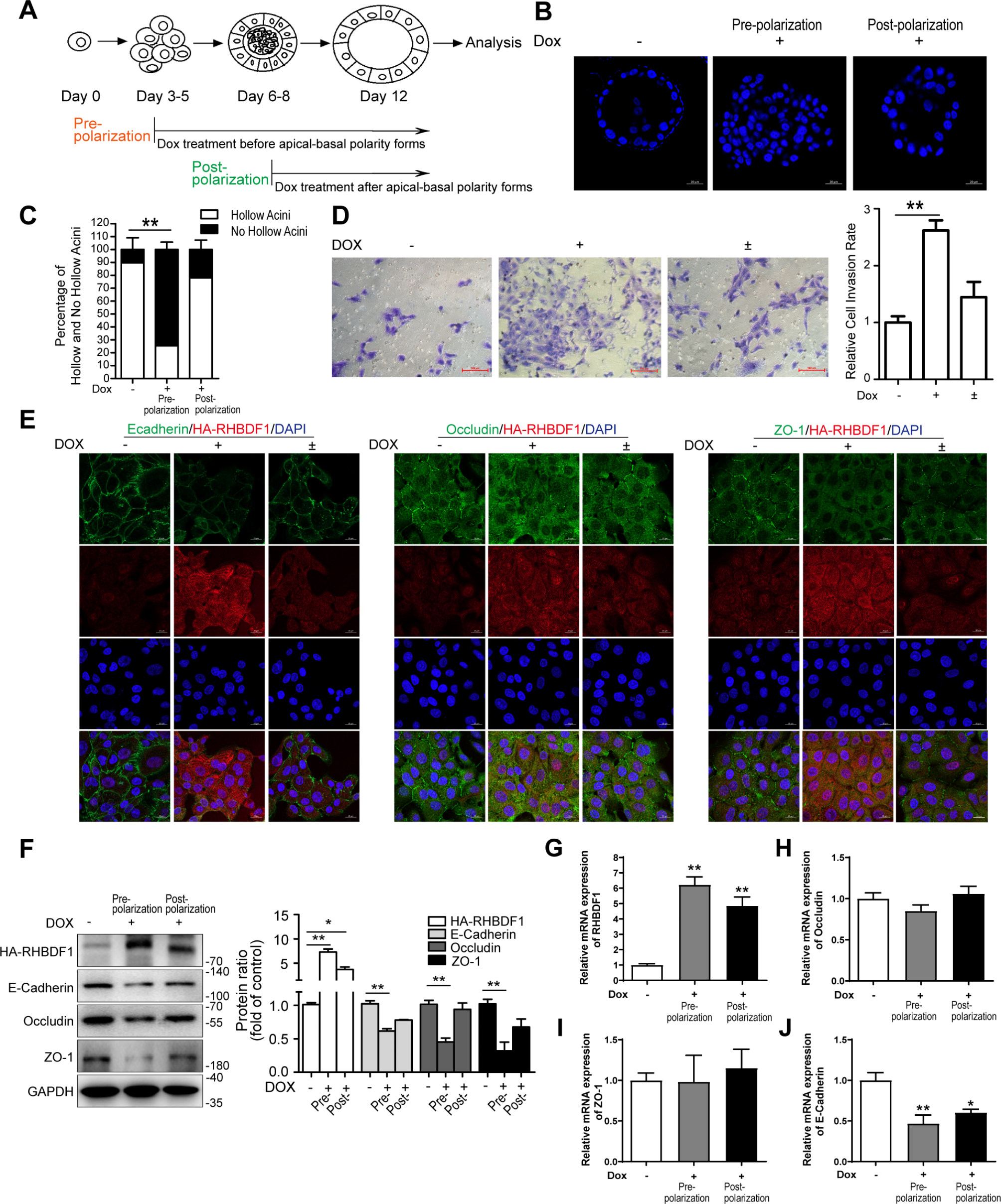
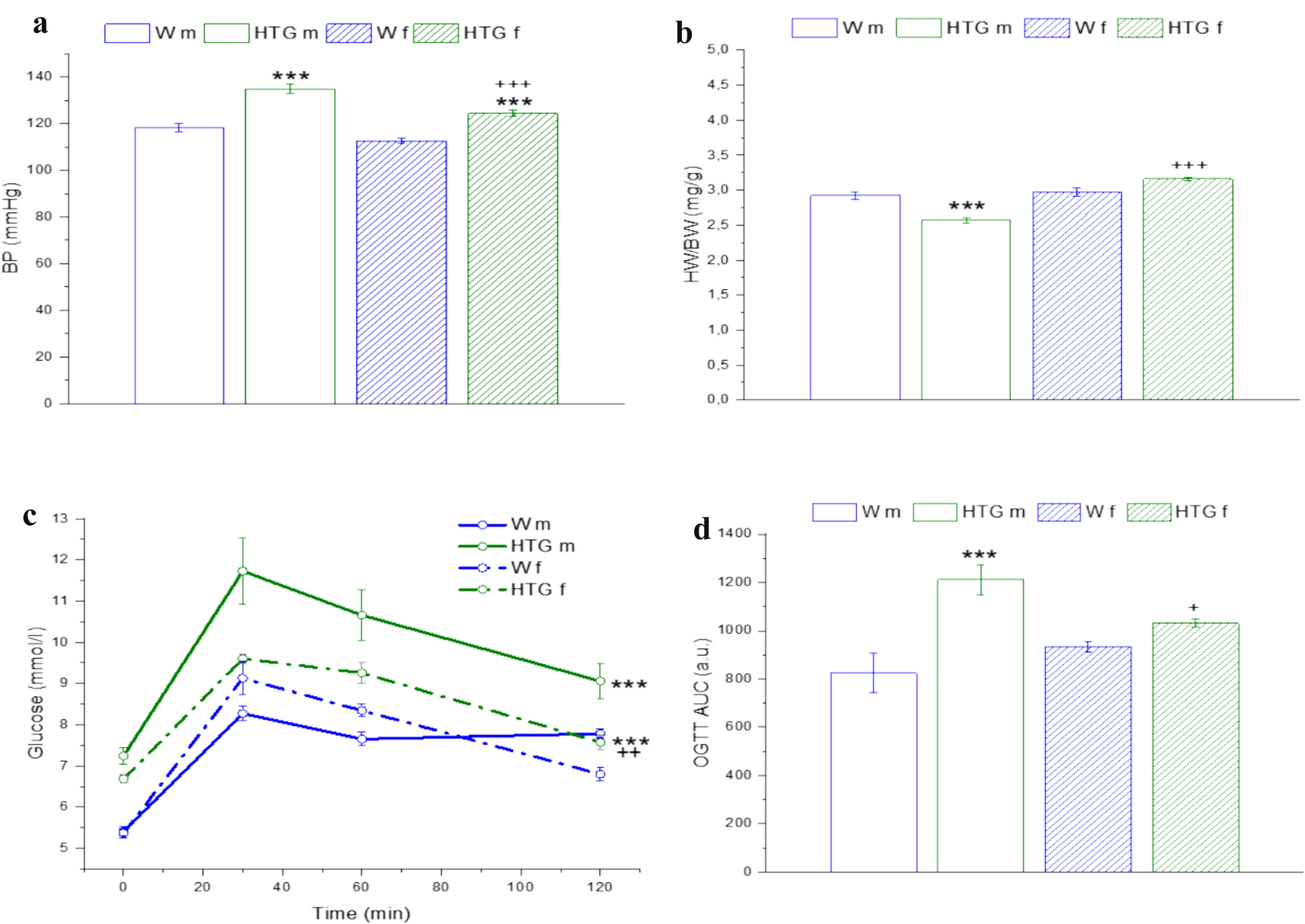
記住我



For the osteogenic differentiation of mesenchymal C3H10T1/2 cells, the literature commonly cites two primary components: β-glycerophosphate and ascorbic acid. A third component varies across studies [35]. In our experimental conditions, we utilized all-trans retinoic acid (ATRA) as this third variable component. We used the alkaline phosphatase staining method to confirm the differentiation of C3H10T1/2 cells into osteoblasts after 7, 14, and 21 days of this treatment (Fig. 1A). We observed a progressive increase in staining after 7 days of treatment. Additionally, we measured ALP activity at 1, 7, 14, and 21 days of osteogenic treatment (Fig. 1B). An increase in ALP activity was noted after 7 days, which remained consistent at 14 and 21 days. Additionally, we analyzed the expression at 1-, 7-, 14-, and 21-days post-induction of the early marker Alpl (alkaline phosphatase), the intermediate marker Col1a1 (Collagen Type 1 Alpha-1 chain), Runx2 transcription factor, and the late markers Opn (Osteopontin) and Ocn (Osteocalcin) (Fig. 1C and G). Alpl marker showed a rapid increase in expression, peaking on day 7, then declining by day 21 (Fig. 1C), compared with non-differentiated cells on day 1 (indicated by a horizontal line), taken as control cells. Col1a1 expression rose from day 1 and remained elevated throughout the differentiation process expression significantly (Fig. 1D), while Runx2 increased from day 7 onwards (Fig. 1E). Interestingly, although Opn is typically considered a late osteogenesis marker, it exhibited a peak in expression on the first day on the first day after induction of differentiation. This was followed by a decline over the subsequent days, dropping below the levels observed in control cells by day 7, and then rising again by day 14 (Fig. 1F). Ocn reached maximum expression on day 14, maintaining elevated levels at day 21 (Fig. 1G), although its expression levels on days 1 and 7 were lower than control cells.
Fig. 1
Osteogenic differentiation in C3H10T1/2 cells induced by β-glycerophosphate, ascorbic acid, and all trans retinoic acid (ATRA). A Alkaline phosphatase staining illustrating the progression of osteoblastic differentiation in C3H10T1/2 cells over 1, 7, 14, and 21 days of induction. B ALP activity (U/L) in C3H10T1/2 cells over 1, 7, 14, and 21 days of osteogenesis induction. RT-qPCR analysis of the osteogenesis-specific marker in C3H10T1/2 cells. Relative mRNA expression levels of Alpl (C) Col1a1 (D), Runx2 (E), Opn (F), and Ocn (G) osteogenic markers were also measured in differentiated C3H10T1/2 cells over a period of 1, 7, 14, and 21 days of induction. Expression data were normalized against the constitutive ribosomal gene Rplp0. Expression levels are relative to day 1 in undifferentiated cells (set arbitrarily at 1) [horizontal line]. Absence of this line in some graphs is due to overlap with the horizontal axis due to scale adjustments. Data are presented as mean ± SD from a minimum of three independent assays, each performed in triplicate. Statistical significance was assessed using Student’s t-test (***p ≤ 0.001, **p ≤ 0.01, and *p ≤ 0.05)
We also focused on analyzing the expression levels of endogenous Notch1-4 genes, their targets, Hes1 and Hey1, and non-canonical genes, Dlk1 and Dlk2, in both differentiated and non-differentiated C3H10T1/2 cells (Supplementary Fig. 1A–H). In undifferentiated C3H10T1/2 cells, we observed that the expression of all Notch genes increased with cell confluence from day 7, relative to the levels in non-differentiated cells on day 1 (horizontal line), which served as the control cells. Following osteogenic treatment, there was a decrease in Notch1 expression compared to the control cells (Supplementary Fig. 1A), but Notch2 (Supplementary Fig. 1B), Notch3 (Supplementary Fig. 1C), and Notch4 (Supplementary Fig. 1D) exhibited increased expression at all time points. In undifferentiated cells, Hes1 expression decreased after 7 days, correlating with increased cell confluence over time in culture (Supplementary Fig. 1E). However, in cells treated with osteogenic inducers, Hes1 expression levels increased, peaking at day 14 relative to the control cells. As for Hey1, its expression in undifferentiated cells rose with increasing cell confluence, reaching its maximum at day 14 compared to the control cells. Interestingly, upon osteogenic induction, Hey1 expression also increased, achieving its highest level on day 21 of differentiation, compared to the control cells (Supplementary Fig. 1F).
Finally, in undifferentiated cells, Dlk1 expression levels increased with cell confluence, peaking on day 14, compared to control cells (Supplementary Fig. 1G). However, when these cells were subjected to osteogenic inducers, Dlk1 expression remained lower than the control levels. Contrasting with Dlk1, Dlk2 expression was inhibited in undifferentiated cells on days 7, 14, and 21, despite increased cell confluence (Supplementary Fig. 1H). Notably, under osteogenic induction, Dlk2 expression significantly increased on day 1, compared to the control cells.
These findings suggest that Dlk genes are expressed in a coordinated but inverse manner, exhibiting opposite patterns in undifferentiated cells and in cells undergoing osteogenic treatment.
The impact of DAPT, a γ-secretase complex inhibitor,on the osteogenic differentiation of C3H10T1/2 cellsWe first measured the activation of NOTCH1 and NOTCH2 receptors in differentiated cells by detecting the levels of their active intracellular domains, NICD1 and NICD2, respectively (Supplementary Fig. 2A and 2B). When global NOTCH signaling was inhibited using DAPT, we noted a reduction in the levels of both NICD1 and NICD2. These levels were normalized to the total NOTCH1 and NOTCH2 levels, respectively, and compared with non-differentiated cells on day 1 treated with DMSO (horizontal line), which was used as the control cells. The inhibitory effect in the presence of DAPT dissolved in DMSO was also observed when we analyzed the levels of global NOTCH signaling activation in C3H10T1/2 cells, as compared with cells in the presence of DMSO, as evidenced by luciferase assays (Supplementary Fig. 2C). A positive control of luciferase activity by transfecting cells with plasmid pNICD1, which express an active form of the NOTCH1 receptor, compared with cells transfected with an empty vector (V), is also shown (Supplementary Fig. 2D). We also evaluated the expression of two NOTCH receptor target genes, Hes1 and Hey1, in C3H10T1/2 cells undergoing osteoblast differentiation, both with and without DAPT treatment (Supplementary Fig. 2E and 2 F, respectively). Data normalization was performed against non-differentiated cells on day 1 treated with DMSO (horizontal line), and the expression levels of each gene were compared to those in similarly timed differentiated cells treated with DMSO. We found that the expression level of Hes1 increased on days 7 and 14 with DMSO compared to control cells (Supplementary Fig. 2C). However, Hes1 expression decreased in the presence of DAPT (days 1, 7, and 14), relative to control cells. Conversely, while Hey1 expression also increased on days 7 and 14 with DMSO (Supplementary Fig. 2D), DAPT treatment did not significantly alter Hey1 expression levels in differentiated cells, except for a noted inhibition on day 21 of treatment.
Then, we analyzed the impact of DAPT’s inhibitory effect on global NOTCH signaling, on the osteogenic differentiation of C3H10T1/2 cells by employing the alkaline phosphatase staining method. As depicted in Fig. 2A, adding DAPT to non-differentiated cells did not result in any significant change in alkaline phosphatase staining at any of the analyzed time points, compared to the control cells treated with DMSO. However, when DAPT was included in the osteogenic differentiation cocktail, a noticeable decrease in alkaline phosphatase staining was observed at all time points.
Fig. 2
Influence of DAPT, a γ-secretase complex inhibitor, on the expression of osteogenic markers during the differentiation of C3H10T1/2 cells . A Representative images from alkaline phosphatase staining depict the contrast in osteogenic activity of undifferentiated and differentiated C3H10T1/2 cells over 1, 7, 14, and 21 days [D], in both the presence and absence of the DAPT inhibitor. B–F The figure further includes a RT-qPCR analysis of the relative mRNA expression levels of osteogenic markers Alpl (B), Col1a1 (C), Runx2 (D), Opn (E), and Ocn (F) in C3H10T1/2 cells undergoing osteoblastic differentiation with or without DAPT treatment at the same time points. The RT-qPCR data were normalized against the mRNA levels of the constitutive ribosomal gene Rplp0, and expression levels were calculated relative to day 1 in undifferentiated cells treated with DMSO (horizontal line). The absence of the horizontal line in some graphs is due to its overlap with the horizontal axis because of the vertical axis scale. Data are presented as mean ± SD from at least three independent experiments, each performed in triplicate. Statistical significance was determined using Student’s t-test (***p ≤ 0.001, **p ≤ 0.01, and *p ≤ 0.05), with non-significant results indicated as ns
Finally, we evaluated the changes in the expression levels of osteogenic differentiation markers in C3H10T1/2 cells treated with DAPT (Fig. 2B–F). Data were again normalized to non-differentiated cells on day 1 of culture (horizontal line), and the expression of each marker was compared with values from DMSO-treated differentiated cells, serving as control cells. As shown in Fig. 2B, DAPT did not affect Alpl expression on days 1 and 7. However, its expression was significantly inhibited by DAPT on days 14 and 21, compared to control cells. The expression levels of Col1a1 were consistently lower in the DAPT-treated cells across all analyzed time points of osteogenic differentiation, indicating significant inhibition (Fig. 2C). The expression pattern of Runx2 (Fig. 2D) mostly mirrored that of the control group, except for a marked reduction on day 14. The expression of Opn was reduced during days 1, 7, and 14 of osteogenic treatment when compared with control cells (Fig. 2E). Lastly, Ocn expression levels were similar to control cells on days 1 and 7 but showed notable inhibition on days 14 and 21 (Fig. 2F).
Generation of stable transfectant pools for Dlk1 and Dlk2 in C3H10T1/2 cellsGiven their roles as NOTCH signaling inhibitors and their contrasting expression profiles during osteogenic differentiation in C3H10T1/2 cells, we aimed to generate stable transfectant pools of Dlk1 and Dlk2 genes in C3H10T1/2 cells that either overexpress or exhibit reduced expression levels of these genes (Fig. 3). We confirmed that the stable transfections resulted in the expected changes at both mRNA (Fig. 3A and C) and protein levels (Fig. 3B and D), in comparison to cells stably transfected with empty vectors, which served as control cells. These changes at the protein level aligned with the levels of activation of global NOTCH signaling in DLK transfected cells, as evidenced by luciferase assays (Fig. 3E).
Fig. 3
Characterization of Dlk1 and Dlk2 stable transfectant pools in C3H10T1/2 cells. RT-qPCR analysis of the relative mRNA expression levels of Dlk1 in DLK1S and DLK1aS stable transfectant pools (A), and Dlk2 in DLK2S and DLK2aS stable transfectant pools (B). C, D Representative Western blots and densitometric analyses illustrate DLK1 (50–60 kDa) (C) and DLK2 (40 kDa) (D) protein expression levels in these pools. E Global NOTCH signaling activity assessed by luciferase assay in DLK1 and DLK2 transfectants compared to control cells transfected with empty vector (V). The figure also presents RT-qPCR analysis of Hes1 (F) and Hey1 (G) mRNA expression levels in each stable transfectant pool. α-Tubulin was employed as a control for loading and sample quality in Western blot assays. Data from RT-qPCR assays were normalized against the constitutive ribosomal gene Rplp0, with expression levels calculated relative to cells stably transfected with the corresponding empty vector (set arbitrarily at 1) [horizontal line]. Data are presented as mean ± SD from a minimum of three independent assays performed in triplicate. Statistical significance was assessed using Student’s t-test (***p ≤ 0.001, and *p ≤ 0.05), and non-significant results are denoted as ns
We also observed notable effects of DLK protein expression levels on the endogenous expression of Hes1 and Hey1 (Fig. 3F and G), compared to control cells. Specifically, stable overexpression of Dlk1 led to a decrease in Hes1 expression, while Dlk2 overexpression did not significantly alter Hes1 levels (Fig. 3F). Conversely, when the expression of both Dlk1 and Dlk2 was downregulated, there was an activation of Hes1 expression (Fig. 3F). Interestingly, overexpressing DLK1 unexpectedly increased Hey1 expression, whereas cells overexpressing DLK2 showed a significant reduction in Hey1 levels (Fig. 3G). Additionally, a decrease in the expression of both Dlk genes surprisingly resulted in reduced Hey1 expression (Fig. 3G).
We also observed that the expression levels of DLK proteins in undifferentiated C3H10T1/2 cells influenced the endogenous expression of Notch (Supplementary Fig. 3A–D), in comparison to control cells. Specifically, the overexpression of Dlk1 or reduced expression of Dlk2 led to an increase in the expression of all four Notch genes, with a particularly notable effect on Notch1. Conversely, the overexpression of Dlk2 or reduced expression of Dlk1 elevated the expression levels of Notch2, Notch3, and Notch4 but no significative changes were observed in Notch1 expression.
Additionally, we observed that overexpression of Dlk1 in undifferentiated cells resulted in a decrease in Dlk2 expression, while reduced expression of Dlk1 led to an increase in Dlk2 expression (Supplementary Fig. 3E). In contrast, while overexpressing Dlk2 raised Dlk1 expression levels, decreasing Dlk2 expression did not significantly impact Dlk1 expression (Supplementary Fig. 3F).
Analysis of the expression levels of osteogenic markers, as well as Notch, Hes1, and Hey1 genes, in Dlk1 and Dlk2 stable transfectant pools of C3H10T1/2 cells induced to differentiate into osteoblastsWe conducted osteogenesis assays on Dlk stable transfectant pools of C3H10T1/2 cells, assessing first the level of osteogenic differentiation using the alkaline phosphatase staining method (Figs. 4A and 5A). Our observations revealed enhanced osteogenic differentiation in cells where DLK2 was overexpressed or DLK1 expression was reduced, in comparison to cells transfected with an empty vector or non-transfected C3H10T1/2 cells. These results indicate that DLK1 acts as an inhibitor, while DLK2 functions as an activator in the osteogenic differentiation process of C3H10T1/2 cells.
Fig. 4
Alkaline phosphatase staining and analysis of osteogenic marker expression levels in Dlk1 stable transfectant pools of C3H10T1/2 cells. A This part showcases representative images of alkaline phosphatase staining in cell culture wells. It includes both non-transfected and Dlk1 stable transfectant pools of C3H10T1/2 cells that have undergone osteoblastic differentiation. The images capture the staining results at 1-, 7-, 14-, and 21-days [D] post-induction of osteogenic differentiation. The cultures include C3H10T1/2 non-transfected cells, cells transfected with empty vector control [V], and cells from the DLK1S and DLK1aS transfectant pools, providing a comparative view of alkaline phosphatase activity across different genetic modifications and stages of differentiation. In this figure, we also show a RT-qPCR analysis of the relative mRNA expression levels of key osteogenic markers in Dlk1 sense (DLK1S) and antisense (DLK1aS) stable transfectant pools of C3H10T1/2 cells. The markers analyzed include Alpl (B), Col1a1 (C), Runx2 (D), Opn (E), and Ocn (F). The expression levels were measured in cells differentiated into osteoblasts over a period of 1-, 7-, 14-, and 21-days [D] post-induction of osteogenic differentiation. The RT-qPCR data were normalized against the mRNA levels of the constitutive ribosomal gene Rplp0, with expression levels calculated relative to day 1 in cells stably transfected with the empty vector (set arbitrarily at 1) [horizontal line]. The absence of the horizontal line in some graphs is due to its overlap with the horizontal axis resulting from the vertical axis scale adjustments. Results are shown as mean ± SD from a minimum of three independent assays, each performed in triplicate. Statistical significance was assessed using Student’s t-test (***p ≤ 0.001, **p ≤ 0.01, and *p ≤ 0.05), and non-significant results are denoted as ns
Fig. 5
Alkaline phosphatase staining and analysis of osteogenic marker expression levels in Dlk2 stable transfectant pools of C3H10T1/2 cells. A This part showcases representative images of alkaline phosphatase staining in cell culture wells. It includes both transfected and Dlk2 stable transfectant pools of C3H10T1/2 cells that have undergone osteoblastic differentiation. The images capture the staining results at 1-, 7-, 14-, and 21-days [D] post-induction of osteogenic differentiation. The cultures include C3H10T1/2 non-transfected cells, cells transfected with empty vector control [V], and cells from the DLK2S and DLK2aS transfectant pools, providing a comparative view of alkaline phosphatase activity across different genetic modifications and stages of differentiation. In this figure, we also show a RT-qPCR analysis of the relative mRNA expression levels of key osteogenic markers in Dlk2 sense (DLK2S) and antisense (DLK2aS) stable transfectant pools of C3H10T1/2 cells. The markers analyzed include Alpl (B), Col1a1 (C), Runx2 (D), Opn (E), and Ocn (F). The expression levels were measured in cells differentiated into osteoblasts over a period of 1-, 7-, 14-, and 21-days [D] post-induction of osteogenic differentiation. The RT-qPCR data were normalized against the mRNA expression levels of the ribosomal gene Rplp0. Expression levels for each marker were compared with values from cells stably transfected with the empty vector on day 1, set as a baseline (horizontal line). The absence of the horizontal line in some graphs is due to its overlap with the horizontal axis, resulting from the scaling of the vertical axis. Data are presented as mean ± SD from at least three independent assays, each performed in triplicate. Statistical significance was evaluated using Student’s t-test (***p ≤ 0.001, **p ≤ 0.01, and *p ≤ 0.05), and non-significant results are indicated as ns
We further assessed the expression levels of osteogenic markers in Dlk stable transfectant pools by using RT-qPCR (Figs. 4B and F and 5B and F), at 1, 7, 14, and 21 days following the induction of osteogenic differentiation. Data normalization was performed against values from non-differentiated cells on day 1 of culture (horizontal line), and expression levels of each marker were compared with those from differentiated cells stably transfected with the corresponding empty vector, serving as control cells.
Among the transfectant pools, DLK1aS and DLK2S showed notable increases in the expression of Alpl, compared to control cells, with peak expressions on days 7 (Fig. 4B) and 14 (Fig. 5B) of induction, respectively. The DLK2aS transfectant pool (Fig. 5B) displayed a slightly higher expression of Alpl than control cells, reaching its maximum on day 7. In contrast, the DLK1S transfectant pool (Fig. 4B) consistently exhibited lower Alpl expression levels compared to control cells throughout the differentiation process. The DLK1S pool (Fig. 4C) consistently exhibited Col1a1 expression levels that were equal to or lower than those of control cells throughout the differentiation process. A similar trend was seen in the DLK2aS pool (Fig. 5C), except for a significant increase on day 1 of differentiation. On the other hand, both the DLK1aS (Fig. 4C) and DLK2S (Fig. 5C) transfectant pools showed higher Col1a1 expression levels compared to control cells, except on day 14, where DLK1aS exhibited similar expression levels to the controls. Notably, the DLK1aS (Fig. 4D), DLK2S (Fig. 5D), and DLK2aS (Fig. 5D) transfectant pools exhibited a significant increase in Runx2 expression. Among these, the DLK1aS pool showed the highest expression levels, peaking on day 14 post-induction. The DLK2aS pool reached its maximum Runx2 expression earlier, on day 7 of differentiation, whereas DLK2S attained its peak on day 14. In contrast, the Runx2 expression level in the DLK1S transfectant pool (Fig. 4D) was consistently similar to or even lower than that observed in control cells. Regarding the Opn marker, DLK1S and DLK2aS pools consistently demonstrated lower expression levels than control cells, while DLK2S and DLK1aS pools experienced a significant increase, especially noticeable in the DLK1aS pool (Figs. 4E and 5E). The highest expression of Opn in the DLK2S pool occurred on day 14 of differentiation, and in the DLK1aS pool, it happened on day 21. We found that the DLK2S, DLK1aS, and DLK2aS stable transfectant pools exhibited high expression levels of Ocn, with DLK1aS cells showing particularly elevated values (Figs. 4F and 5F). The DLK2S and DLK2aS pools (Fig. 5F) reached their peak Ocn expression on day 21 of differentiation, whereas the DLK1aS pool (Fig. 4F) displayed increased levels starting from day 7. In contrast, the DLK1S pool (Fig. 4F) showed Ocn expression levels similar to those of control cells. These results lead us to conclude that while both DLK1 and DLK2 proteins are inhibitors of NOTCH receptor signaling, DLK1 inhibits and DLK2 potentiates osteogenesis.
Given the contrasting effects of DLK1 and DLK2 proteins on the osteogenesis process in C3H10T1/2 cells, despite both proteins are NOTCH signaling inhibitors, we were motivated to investigate how varying levels of DLK1 and DLK2 in these transfectant pools influence the expression of Notch genes and their targets, Hes1 and Hey1, during osteoblast differentiation (Supplementary Figs. 4 and 5). We normalized data against the values from non-differentiated cells on day 1 of culture (horizontal line). For each marker, expression levels were compared to those of differentiated cells stably transfected with the corresponding empty vector, which served as control cells.
Notably, all transfectant pools experienced a marked decrease in Notch1 expression on the first day of osteogenic differentiation, particularly evident in the DLK1aS (Supplementary Fig. 4A) and DLK2S (Supplementary Fig. 5A) pools. Regarding Notch2, the DLK1S and DLK1aS pools (Supplementary Fig. 4B) showed the highest expression levels compared to controls. The DLK2S and DLK2aS pools (Supplementary Fig. 5B) also had elevated Notch2 expression, with the DLK2aS pool being notably higher. For Notch3, all transfectant pools exhibited expression levels equal to or surpassing those of control cells (Supplementary Fig. 4 C and 5 C). The DLK1aS pool had the highest expression, closely followed by DLK2S. The DLK1S and DLK2aS pools showed Notch3 levels similar to controls. Finally, in the case of Notch4, the DLK1aS (Supplementary Fig. 4D) and DLK2S (Supplementary Fig. 5D) pools demonstrated higher expression than control cells, with DLK2S having the most pronounced increase. The DLK1S and DLK2aS pools displayed Notch4 levels equal to or lower than control cells.
As indicated in Supplementary Fig. 4E and 5E, the Hes1 expression level increased during osteogenic differentiation across all transfectant pools compared to control cells, with the DLK2S pool (Supplementary Fig. 5E) showing the highest Hes1 expression. Conversely, Hey1 expression was highest in the DLK1S cells, increasing during osteogenic differentiation, while it was lower in DLK1aS cells compared to controls (Supplementary Fig. 4F). In the DLK2S and DLK2aS pools, Hey1 expression was lower than in control cells (Supplementary Fig. 5F).
These findings suggest that DLK1 and DLK2 have differential effects on the expression of Notch genes and their targets during osteogenic differentiation. This difference may partially account for the opposing impacts of DLK1 and DLK2 on osteogenesis as observed in our study.
Analysis of kinase signaling pathways involved in the osteogenesis of C3H10T1/2 cellsWe explored potential interactions between DLK proteins and some key kinase signaling pathways involved in osteogenesis. We first conducted osteogenesis assays in non-transfected C3H10T1/2 cells and used the alkaline phosphatase staining method to assess the effects of inhibiting ERK1/2 MAPK, PI3K/AKT, mTOR (mammalian Target of Rapamycin), and p38 MAPK signaling pathways on osteogenic differentiation at 1, 7, 14, and 21 days (Supplementary Fig. 6). Our findings revealed that the addition of U0126, an ERK1/2 MAPK inhibitor, led to a significant reduction in osteogenic activity compared to control cells, as indicated by alkaline phosphatase staining. Interestingly, treatment with the mTOR inhibitor rapamycin resulted in greater staining than control cells. The PI3K/AKT pathway inhibitor LY294002 yielded staining similar to controls. Lastly, inhibiting p38 MAPK signaling with SB203580 significantly reduced staining compared to controls. Our findings demonstrated that ERK1/2 MAPK and p38 MAPK pathways enhance osteogenic differentiation in C3H10T1/2 cells, while mTOR kinase serves as an inhibitor and PI3K/AKT may be less involved.
We observed that phosphorylation of ERK1/2 increased throughout the osteogenic process of non-transfected C3H10T1/2 cells compared to the baseline on day 1 in undifferentiated cells, which we used as control cells (Supplementary Fig. 7A). The peak phosphorylation level was reached on day 14 post-induction of osteogenic treatment. We next analyzed the phosphorylation levels of this kinase in DLK1 and DLK2 stable transfectants during osteogenic differentiation. Data were normalized against non-differentiated cells on day 1 of culture (horizontal line), and phosphorylation levels were compared with those of differentiated cells stably transfected with the corresponding empty vector, serving as control cells.
We then analyzed the phosphorylation levels of ERK1/2 MAPK in all DLK protein stable transfectants throughout the osteogenic differentiation process (Fig. 6A and B). In the early phase (days 1 and 7), in DLK2 transfectants, and on day 7, in the DLK1aS transfectant, ERK1/2 MAPK phosphorylation levels decreased compared to controls. DLK1S transfectants showed similar levels to controls during these days. However, by day 14, an increase in ERK1/2 MAPK phosphorylation was noted in both DLK1aS and DLK2S transfectants, while DLK1S transfectant did not exhibit significant variations. The DLK2aS transfectant displayed lower phosphorylation levels than control cells. By day 21, most transfectants showed no significant changes in phosphorylation levels compared to controls, except for the DLK1aS transfectant, which maintained elevated phosphorylation levels. These findings suggest that the ERK1/2 MAPK kinase is actively involved in the osteogenic process of these cells and its activity may be modulated by the levels of DLK proteins.
Fig. 6
ERK1/2 MAPK phosphorylation dynamics in Dlk1 and Dlk2 stable transfectant pools during osteogenic differentiation. A Representative Western blot images displaying the phosphorylation levels of ERK1/2 MAPK (42–44 kDa) in Dlk1 and Dlk2 stable transfectant pools of C3H10T1/2 cells at various stages of osteogenic differentiation (1, 7, 14, and 21 days [D]). B Densitometric analysis quantifying ERK1/2 MAPK phosphorylation levels in Dlk1 and Dlk2 transfectants during osteogenic differentiation. The data are normalized to total ERK1/2 MAPK expression, serving as control for loading and sample integrity. The baseline phosphorylation level was set using data from non-differentiated cells stably transfected with the empty vector on day 1 (indicated by the horizontal line). The densitometric results are presented as the mean ± SD from at least three independent experiments, each performed in triplicate. Statistical analysis was conducted using Student’s t-test to compare each transfectant pool at different time points against the day 1 baseline, with significance levels marked as ***p ≤ 0.001, **p ≤ 0.01, and *p ≤ 0.05. Results not reaching statistical significance are denoted as ns
Our findings also indicated a progressive increase in the phosphorylation of p38 MAPK, peaking on day 7 of differentiation during the osteogenic differentiation of non-transfected C3H10T1/2 cells (Supplementary Fig. 7B). We then analyzed the phosphorylation levels of p38 MAPK in all DLK protein stable transfectants throughout the osteogenic differentiation process (Fig. 7A and B). For consistency, data were normalized against non-differentiated cells on day 1 of culture (horizontal line), and phosphorylation levels were compared to those in differentiated cells transfected with the corresponding empty vector, used as control cells. Our results revealed that in transfectants with decreased Dlk1 expression, there was an increase in p38 MAPK phosphorylation across the entire differentiation period. In contrast, the other DLK stable transfectants, including those overexpressing Dlk1 and Dlk2, did not exhibit significant differences in p38 MAPK phosphorylation levels during osteogenic differentiation.
Fig. 7
p38 MAPK phosphorylation dynamics in Dlk1 and Dlk2 stable transfectant pools during osteogenic differentiation. A Representative Western blot images showcase the phosphorylation levels of p38 MAPK phosphorylation in Dlk1 and Dlk2 stable transfectant pools of C3H10T1/2 cells at various stages of osteogenic differentiation (1, 7, 14, and 21 days [D]). B Densitometric analysis provides a quantitative assessment of p38 MAPK phosphorylation in DLK1 and DLK2 transfectant pools. The data are normalized to total p38 MAPK expression, serving as controls for loading and sample integrity. The baseline phosphorylation level was set using data from non-differentiated cells stably transfected with the empty vector on day 1 (indicated by the horizontal line). The densitometric results are presented as the mean ± SD from at least three independent experiments, each performed in triplicate. Statistical analysis was conducted using Student’s t-test to compare each transfectant pool at different time points against the day 1 baseline, with significance levels marked as ***p ≤ 0.001, and **p ≤ 0.01. Results not reaching statistical significance are denoted as ns
留言 (0)