
記住我



We have previously reported that FBLN2 is preferentially expressed at the terminal end buds (TEBs) of the pubertal mouse mammary gland and not in the ductal epithelium [14]. Using immunofluorescence, we here showed that ITGβ1 and ITGα3, markers of myoepithelial cells [37,38,39,40] exhibit similar patterns of expression to FBLN2 (Supplementary Fig. 1). To further assess their relative co-localization in the TEBs, we performed a co-immunofluorescence staining which showed that FBLN2 is exclusively expressed at the cap cells of TEBs, and not in luminal cells, co-localizing with ITGβ1 and ITGα3 (Fig. 1a, b and supplementary Fig. 2).
Fig. 1
FBLN2 is co-localized with ITGβ1 and ITGα3 in the terminal end buds of pubertal mouse mammary gland in mice. a Double staining of ITGβ1 and FBLN2 in TEBs of the mouse pubertal mammary gland showing co-localization of both markers at the cap cells, and not in the luminal cells (n = 3). Bars represent 50 μm. b Immunofluorescence staining of ITGα3 and FBLN2 in consecutive sections of TEBs of pubertal mouse mammary gland showing co-localization of both markers at the cap cells, and not in the luminal cells (n = 3)
TGFβ3 stimulates FBLN2 overexpression in mouse mammary epithelial cellsAs TGFβ3 is also mainly expressed in the cap cells of TEBs [41] and drives BM protein expression, we asked whether TGFβ3 treatment will stimulate FBLN2 expression in mammary epithelial cells. TGFβ3 treatment of the morphologically normal mouse mammary cell line induced the transformation to a spindle-shaped morphology, a characteristic of myoepithelial cells, (Fig. 2a). It also induced the expression of FBLN2 in a dose-independent manner, evident both on mRNA and protein levels (Fig. 2b and c); however, it was statistically insignificant. Further, TGFβ3 treatment of spontaneously immortalized mouse embryonic fibroblasts (MEFs) resulted in an equally dose-independent upregulation of the FBLN2 protein (Supplementary Fig. 3).
Fig. 2
Effect of TGFB3 treatment on FBLN2 expression in EpH4 cells. a Phase contrast microscopy of cells showing the transformation to a spindle-shaped morphology, reminiscent of myoepithelial cells, upon treatment of cells with TGFβ3. b Relative mRNA expression of Fbln2 upon treatment of EpH4 cells with different concentrations of TGFβ3 showing the upregulation of Fbln2 in a dose-independent manner. Error bars represent the standard error of three biological replicates. c Immunoblotting of FBLN2 expression upon treatment of EpH4 cells with ascending concentrations of TGFβ3, confirming the upregulation of FBLN2 after TGFβ3 treatment, relative to βactin expression (n = 3)
FBLN2 KD drives a luminal phenotype in mouse mammary cellsTo further examine the association of FBLN2 to basal phenotype in mammary cells, we assessed the expression of the myoepithelial/basal marker Krt14 [42, 43], and the luminal marker Krt18 [42, 44] in EpH4 cells in which FBLN2 was KD. Immunoblotting analysis of EpH4 cell lysates with different levels of FBLN2 KD (80, 28, and 18%) [14] showed a reduced expression of KRT14 and an increased expression of KRT18 upon FBLN2 KD (Fig. 3a). Flow cytometry analysis of one biological replicate confirmed this shift toward luminal phenotype upon FBLN2 KD (Supplementary Fig. 4). Immunofluorescence staining using monolayered and Matrigel-embedded cells also showed an increased expression of KRT18; however, the levels of expression among cells appeared heterogeneous, where cells with an increased expression of KRT18 being larger in size in monolayer cultures (Fig. 2c and 3b).
Fig. 3
FBLN2 KD correlates with a reduction in the basal cell marker Krt14 and an increase in luminal cell marker Krt18. a Immunoblotting analysis of Krt18 and Krt14 in control and FBLN2 KD cells using three shr1 (70%), 2 (28%), and 3 (18%) FBLN2 expression cells. Expression was relative to βactin (n = 3). b Immunofluorescence staining of Krt18 and Krt14 in Matrigel-embedded control and FBLN2 KD cells [70% reduction in FBLN2 expression cells]. Scale bars are 50 µm (n = 3). c Immunofluorescence staining of Krt18 and Krt14 in monolayered control and FBLN2 KD cells (n = 3). Scale bars are 50 µm
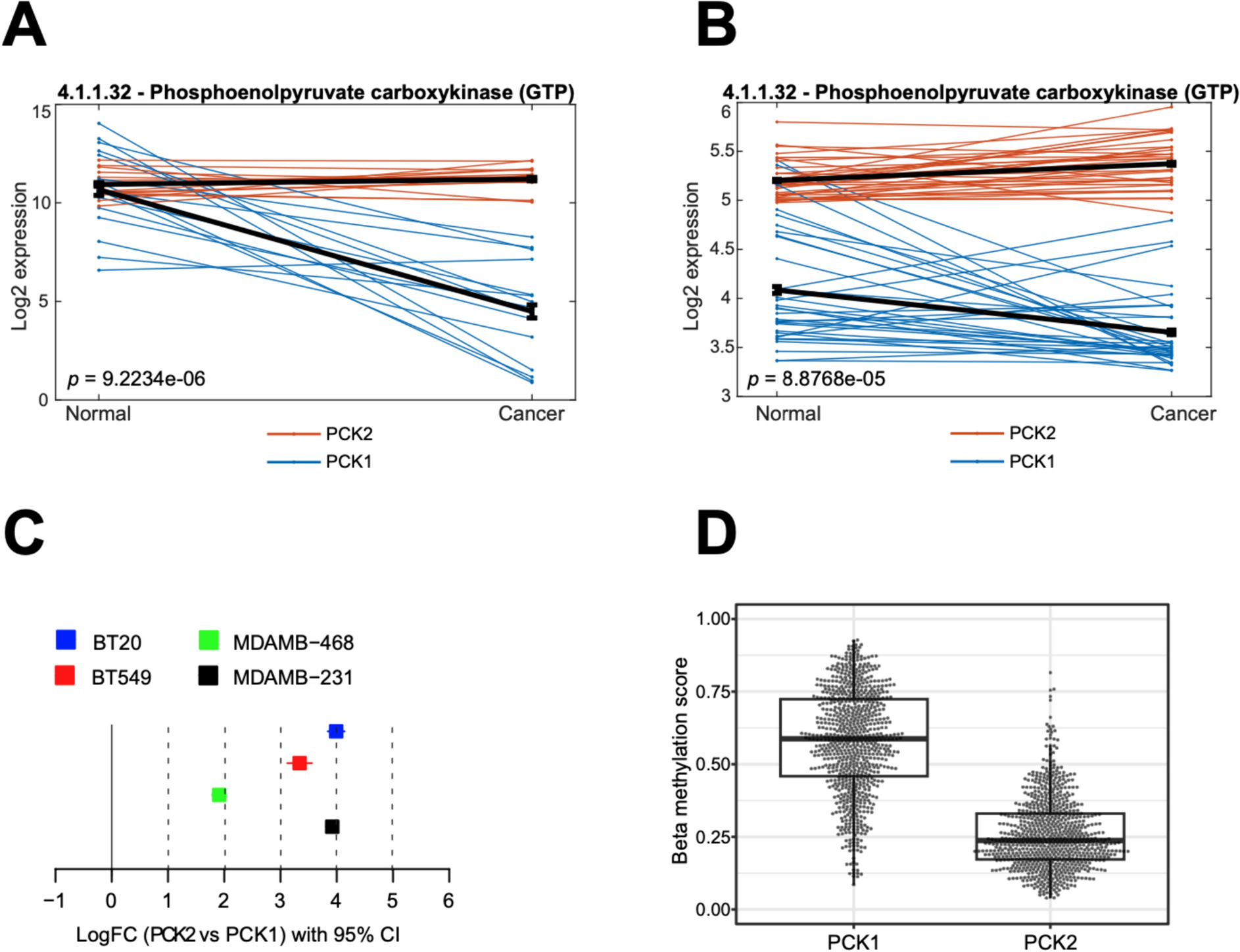
Fbln2 mRNA is preferentially correlated to tuman mammary basal markersTo investigate the preferential expression of FBLN2 in basal cells in human breast cancer, we retrieved online datasets from the Bioportal [28,29,30] (METABRIC dataset) and assessed the correlation between mRNA levels of Fbln2 and the available data on luminal, myoepithelial, EMT, stemness, and mesenchymal markers. Generally, METABRIC dataset analysis showed a negative correlation of Fbln2 with luminal markers (Krt18, Krt19, Ddr1, and Krt8) [37,38,39,40], and a positive correlation with myoepithelial cell markers (Krt14, Itgb1, ITtga3, Krt7, Krt5, Krt17, and Krt14) [37,38,39,40] and mesenchymal markers (Fn, Vim, Cdh2, and Cdh11) [45] in the majority of the molecular and clinical subtypes of breast cancer patients [including grouping based on hormonal receptors, LN status, tumor grade, growth factor receptor status, and tumor proliferation] (Fig. 4a). Markers of EMT (Sna12, Zeb1, Twist1, Zeb2,) [45] and markers of stemness (Itga6, Epcam, Prom1,Abcg2, Cd44, Aldh1a1, Sox2, Thy1, and Cd24) [46] showed different patterns of correlations with Fbln2 (Fig. 4a and Supplementary Fig. 5a). Of note, Fbln2 had significant positive correlations with Fbln1 and Fbln5 in all breast cancer subtypes (Fig. 4 and Supplementary Fig. 5). P values, and number of patients per group are summarized in Supplemental Table 1.
Fig. 4
Fbln2 mRNA expression positively correlates with basal and mesenchymal markers in human breast cancer. a Correlation matrix of data retrieved from online datasets (METABRIC) for Fbln2 against luminal, myoepithelial, EMT, and mesenchymal markers. Numbers refer to the R coefficient of Pearson correlation between Flbn2 and each marker. b Correlation matrix generated by TNMplot.com for Fbln2 with luminal, myoepithelial cell markers, EMT, mesenchymal, stemness, and other Fblns in breast tumor. ER + Her2 − HP: ER + Her2 − High proliferation, ER + Her2 − LP: ER + Her2 − Low Proliferation, IDC: Invasive Ductal Carcinoma, ILC: Invasive Lobular Carcinoma, IMM: Invasive Mixed Mucinous, IMDLC: Invasive Mixed Ductal and Lobular Carcinoma, and IBC: Invasive Breast Carcinoma
Further, analysis of the retrieved dataset from TNMplot tool in KM database showed the same pattern of correlation with Fbln2 in breast cancer group (n = 7569) (Fig. 4b); however, myoepithelial markers showed a negative correlation with Fbln2, except for Itgb1, in the morphologically normal breast group (n = 242) (Supplementary Fig. 5b).
In addition, the relation of fbln2 to basal/mesenchymal and stemness phenotype within mammary gland was validated using the data extracted from Girardi’s datasets [47] (Supplementary Fig. 6).
Fbln2 mRNA has a distinct expression in different subtypes of human breast cancerWe have previously reported that FBLN2 contributes to BM integrity in mouse mammary gland and human breast cancer [14]. Retrospective analysis of a previously examined cohort of 65 human breast cancer patients [14] revealed that FBLN2 generally had a higher expression in control regions (safety margins) compared to DCIS and IDC. Further, Her2 + and triple-negative patients had a higher expression of FBLN2 compared to LumA and LumB at lesions of invasive ductal carcinoma (IDC), ductal carcinoma in situ (DCIS), and control regions (Supplementary Fig. 7). To test these observations in a larger dataset, we retrieved human breast cancer mRNA profiles from the datasets METABRIC and TCGA, registered at the Bioportal [28,29,30,31], with stratification of patients according to their molecular and clinical subtypes. In METABRIC dataset, Fbln2 was significantly upregulated in Claudin-low compared to LumA (P < 0.0001); however, it was upregulated in LumA when compared to Her2 + (P < 0.001) and LumB (P < 0.0001) (Fig. 5a).
Fig. 5
Preferential expression of FBLN2 in molecular subtypes of human breast cancer. a–j Fbln2 mRNA expression in different classification of human breast cancer [molecular subtypes, ER status, PR status, Her2 status, LN status, tumor grade, and tumor proliferation]. *P < 0.05, **P < 0.01, ***P < 0.001. G1: grade 1, G2: grade 2, G3: grade 3, ER + Her2− HP: ER + her2− High proliferation, ER + Her2− LP: ER + Her2− Low Proliferation, IDC: Invasive Ductal Carcinoma, ILC: Invasive Lobular Carcinoma, IMM: Invasive Mixed Mucinous, IMDLC: Invasive Mixed Ductal and Lobular Carcinoma, and IBC: Invasive Breast Carcinoma
In relation to hormonal and growth factor receptors, ER-patients showed a significant increase in Fbln2 expression when compared to ER + patients (P < 0.0001) (Fig. 5b); however, no preferential expression was observed based on PR and Her-2 status in this dataset (Fig. 5c and d). According to tumor grade, Fbln2 expression was significantly decreased in grade 3 compared to grade 1 (P < 0.01) and grade 2 (P < 0.05) (Fig. 5e). Fbln2 expression did not significantly change upon subgrouping based on LN status (Fig. 5f). Patients with higher tumor proliferation ER + Her2- showed a significant downregulation of Fbln2 compared to Lower proliferation group (P < 0.0001) (Fig. 5g). Based on pathological classification of patients, Fbln2 was significantly downregulated in IDC patients when compared to invasive lobular carcinoma (ILC) (P < 0.0001). Invasive mixed mucinous breast cancer showed a significant downregulation of Fbln2 compared to IDC (P < 0.001), ILC (P < 0.0001), mixed ductal carcinoma (P < 0.001), and invasive breast cancer (P < 0.05) (Fig. 5h).
These patterns of expression appeared to be conserved when analyzing the TCGA dataset (Molecular subtypes, ER status, PR status, and LN status); however, Her2− patients had a significantly lower expression of Fbln2 compared to Her2 + patients (Supplementary Fig. 8). Our examination of METABRIC dataset showed that the expression patterns of Krt14, Itgβ1, and Itgα3 were the same patterns as Fbln2 expression in molecular subtypes ensuring the association between fbln2 and basal markers (Supplementary Fig. 1). Interestingly, methylation status of the Fbln2 promotor across molecular subtypes supports the different expression patterns of Fbln2 (Supplementary Fig. 9), suggesting that expression of FBLN2 may be epigenetically regulated.
FBLN2 expression in human breast cancer cell linesTo investigate FBLN2 expression in the different molecular subtypes of human breast cancer, meta-analyses of publicly available datasets of breast cancer cell lines from Bioportal [28,29,30,31] and the Human Protein Atlas [35] revealed that FBLN2 had a lower expression in cell lines that represent LumA and LumB subtypes and had a higher expression in cell lines that represent Her2 + and triple-negative subtypes (Fig. 6).
Fig. 6
FBLN2 expression in human breast cancer cell lines. Heatmap of Fbln2 mRNA expression levels in different human breast cancer cell lines categorized based on their molecular subtypes and pathological classification. nTPM: Normalized transcript per million, rPKM: Reads per kilobase per million mapped reads, IDC: Invasive ductal carcinoma, ILC: Incasive Lobular Carcinoma, AC: Adenocarcinoma, MC: Medullary carcinoma, Sqc: Squamous cell carcinoma, infDC: Inflammatory ductal carcinoma, and AnC: anaplastic carcinoma
Higher FBLN2 expression is associated with better survival in less advanced bbreast cancerTo evaluate the prognostic potential of FBLN2 expression in breast cancer patients, we assessed the relevance of Fbln2 mRNA expression to patients’ survival status using data from the KM plotter and METABRIC. With multiple analyses in KM plotter dataset, higher Fbln2 expression was significantly associated with better OS in patients with Her2− status (P = 0.041, n = 531) following chemotherapy. RFS improved with higher Fbln2 expression in grade 2 patients (P = 0.04, n = 1177), unstratified breast cancer patients following chemotherapy (P = 0.047, n = 1935), Her2− patients following chemotherapy (P = 0.011, n = 1555), LN-patients following chemotherapy (P = 0.047, n = 1304), ER + patients following chemotherapy (P = 0.0043, n = 1229), grade 2 patients following chemotherapy (P = 0.0076, n = 580), and grade 2 patients following hormonal therapy (P = 0.018, n = 594). In contrast, lower Fbln2 expression was associated with better OS in grade 3 patients (P = 0.041, n = 586). RFS improved with lower expression of Fbln2 in Her2 + patients (P = 0.049, n = 695), ER-patients (P = 0.036, n = 1161), grade 3 patients (P = 0.033, n = 1300), normal following treatment (P = 0.014, n = 119), and Her2 + patients following treatment (P = 0.05, n = 695) (Fig. 7).
Fig. 7
Higher Fbln2 mRNA expression is associated with better survival in less advanced human breast cancer. KM plots for overall survival (a and b), and Relapse-free survival (c–m) based on Fbln2 expression levels, showing the stratification of breast cancer patients based on grade, Her2 status, LN status, and therapy regimen. High mRNA expression in red and low expression in black. The numbers below each plot are the numbers of individuals at risk in each group
In TCGA dataset, lower Fbln2 expression was associated with better OS in LumB patients (P < 0.05), and higher Fbln2 expression was associated with better OS in Her2 + patients (P < 0.01) (Supplementary Fig. 10).
留言 (0)