
記住我
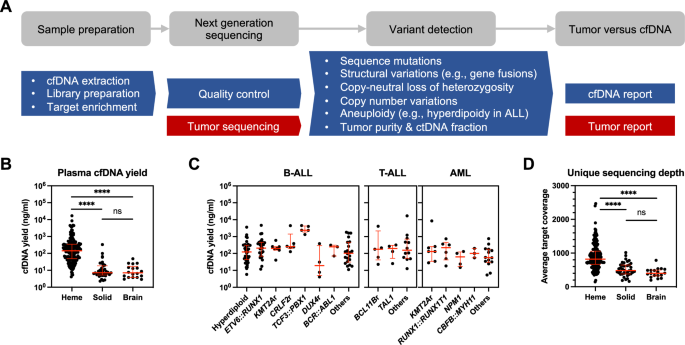
We analyzed Affymetrix GeneChip™ Human Genome U133 Plus 2.0 Array gene expression profiling results derived from mRNA/cDNA isolated from CD19+ flow sorted CLL samples from 117 CLL patients. Most CLL samples had been previously characterized for relevant CLL traits [7, 17]. Using supervised data analyses, we grouped probe level expression values by CLL del17p/TP53mut status, resulting in the comparison of N = 26 CLL with del17p/TP53mut (N = 23) or non-del17p/TP53mut (N = 3) with (N = 91) of CLL with varying genetic backgrounds and TP53 WT. The gene TP53 is located on chromosome 17p at 7.5 Mb and within del17p and was expressed at a log2-fold of −0.94 (211300_s_at) when comparing del17p/TP53mut with non-del17p CLL cases. Overall, 233 probe sets measured differential gene expression at log2-fold >1 or <−1, indicative of profound changes associated with del17p/TP53mut status in CLL. (Supplementary Fig. 1; Supplementary Table 3).
We detected much lower expression of the phosphatase LAIR1 in CLL with del17p/TP53mut status (LAIR1 210644_s_at: log2-fold change −2.35 at FDR 0), a gene previously identified as a negative regulator of BCR signaling [18]. We also identified substantially lower expression of the transmembrane phosphatase PTPRS, which is likely involved in fine tuning of CLL signal transduction pathways and the subject of ongoing experimental studies (PTPRS 229465_s_at: log2-fold change −2.45 at FDR 0 and PTPRS 226571_s_at: log2-fold change −2.26 at FDR 0, respectively).
The top ranked gene expressed at high levels in del17p/TP53mut CLL was the metabolic enzyme branched chain amino acid transferase BCAT1 (BCAT1 225285_at: log2-fold change 2.3 at FDR 0, and BCAT1 226517_at: log2-fold change 2.17 at FDR 0, respectively), which had not previously been identified in CLL and thus was selected for the studies described herein.
Aberrant expression of BCAT1 preferentially in CLL with del17p/TP53mut or trisomy 12Using purified CLL cells depleted of CD3+ and CD14+ cells and detergent cell lysates, we performed quantitative immunoblotting on 205 CLL and compared BCAT1 expression with the mean intrablot expression of BCAT2 (a universally expressed mitochondrial enzyme, which cross reacts with the BCAT1 antibody used) or in parallel with HSP90. Using mRNA/cDNA isolated/generated from flow-sorted CLL we measured BCAT1 expression by qRT-PCR compared with the housekeeping gene PGK1 in N = 269 CLL.
The immunoblotting data for BCAT1 revealed the complete absence of BCAT1 expression in ~70% of CLL and variable to high expression in the remainder of cases (Fig. 1A, B). We performed densitometric data quantification by deriving BCAT1/mean intrablot BCAT2 ratios and plotted such ratios over CLL genomic subtypes. We confirmed that 66% of CLL with del17p/TP53mut status expressed BCAT1, while the large majority of CLL with isolated del13q14 (85%) or normal FISH (71%) were BCAT1 expression null. Equally, 77% of CLL with trisomy 12 expressed BCAT1 and in this context it is relevant that BCAT1 is located on chromosome 12 (12p12.1; hg38; 24810022 – 24949459), indicating that the selected extra chromosome 12 permits augmented BCAT1 expression.
Fig. 1: Aberrant expression of BCAT1 in genomic CLL subgroups carrying del17p/TP53mut or trisomy 12.
A Displayed are immunoblot band density ratios of BCAT1: mean intrablot BCAT2 for detergent lysates from N = 205 primary CD19 + CLL purified via depletion of CD3+ and CD14+ cells categorized by indicated genomic status. B Representative BCAT1 immunoblot results for detergent lysates from primary CD19 + CLL. C Displayed are qRT-PCR based delta Ct values for BCAT1-PGK1 based on RNA/cDNA from N = 269 CD19+ flow sorted CLL categorized by indicated genomic status. A threshold delta CT value of 2.5 is highlighted above which no BCAT1 protein was detectable by immunoblotting. D Display of immunoblot band density ratios of BCAT1: mean intrablot BCAT2 (y-axis) and paired delta Ct values for BCAT1-PGK1 (x-axis) for individual CLL cases. A threshold delta CT value of 2.5 is highlighted above which no BCAT1 protein was detectable. BCAT1 branched chain amino acid transferase 1, BCAT2 branched chain amino acid transferase 2, CLL: Chronic Lymphocytic Leukemia, qRT-PCR quantitative Reverse Transcriptase-Polymerase Chain Reaction, Ct Cycle threshold, cDNA complementary DNA, PGK1 phosphoglycerate kinase 1.
The results for the BCAT1 qRT-PCR based mRNA expression analysis in CLL are very similar to the BCAT1 immunoblotting data showing preferential BCAT1 expression in CLL with del17p/TP53mut status and in CLL carrying trisomy 12 (Fig. 1C). Using a correlation analysis of BCAT1 protein expression (BCAT1/BCAT2 protein ratios) versus BCAT1 mRNA expression (delta Ct BCAT1-PGK1), we detected a clear cut-off in that no CLL samples with a delta Ct BCAT1-PGK1 ≥ 2.5 expressed BCAT1 protein (Fig. 1D).
We performed a similar analysis for BCAT2 protein expression (BCAT2/mean intrablot BCAT2) across the genetic CLL types and in contrast to BCAT1, detected no preferential expression (Supplementary Fig. 2A). In addition, BCAT2 was expressed in all CLL cases and at relatively uniform levels. Further, lower BCAT2 expression did not correlate with acquired high BCAT1 expression (Supplementary Fig. 2B).
Lack of expression of BCAT1 in human non-malignant lymph node derived B cellsGiven that ~30% of CLL cases expressed BCAT1 protein while ~70% did not, we wanted to ascertain if the BCAT1 expression is aberrant in CLL. Published RNA seq data (https://genome.ucsc.edu/) detected only minimal BCAT1 mRNA expression in spleen. We purified B cells by depleting T cells and macrophages from five non-malignant cryopreserved human lymph node biopsies and performed BCAT1 immunoblotting. We included the CLL-derived cell lines HG3, PGA-1 and C1 and two CLL samples and lysates from 293 T cells transiently transfected with a BCAT1 cDNA. The results overall confirmed absence of BCAT1 expression in human lymph node derived B cell populations and thus aberrant expression and a gain-of-function phenotype in CLL subsets (Supplementary Fig. 3).
Increased MTORC1 activity following BCR crosslinking in CLL with aberrant BCAT1 expressionMultiple catalytic reaction products (leucine, glutamine via glutamate, acetyl-CoA, and α-ketoglutarate) of BCAT1 are known positive regulators of MTORC1 [19,20,21,22]. To determine if the regulation of MTORC1 by BCAT1 is relevant in CLL, we treated purified CLL cells that were BCAT1 expressing or null (N = 10 cases each) in multiple separate experiments with anti-IgM for 10’, we made NP40 detergent cell lysates, and analyzed cellular protein for p-S6K, actin, BCAT1 and HSP90 by immunoblotting. We quantified the p-S6K signal across CLL samples using blotting standards run on each immunoblot. CLL cases comprised relevant genetic subtypes, including del17p, trisomy 12 and del13q and were also matched for IGHV status.
As can be seen in Fig. 2A–F, CLL cases expressing BCAT1 demonstrated an ~2-fold higher normalized p-S6K signals after BCR crosslinking when compared with BCAT1 null CLL, indicative of higher MTORC1 activity. The increased BCR-induced MTORC1 activity was not a result of unevenly distributed IGHV UM versus M CLL status as shown in the analysis displayed in Supplementary Fig. 4A.
Fig. 2: Elevated sIgM-crosslinking induced MTOR activation in CLL with aberrant BCAT1 expression.
A–E Primary CLL aberrantly expressing BCAT1 or BCAT1 null were purified via Miltenyi column-based depletion of CD3 (T-cells) and CD14 (macrophages) and subsequently incubated in serum supplemented RPMI medium in the presence of anti-IgM or not for 10’. Detergent cell lysates were fractionated by SDS-PAGE and protein prepared for immunoblotting with antibodies targeting the indicated epitopes. The CLL FISH and IGHV status is indicated. (1) diluted lysate from 293 T cells; (2) diluted lysate from 293 T cells transfected with a ATP6V1B2 cDNA [30]; (*) HG3 cell line lysate. F Displayed are immunoblot based band density ratios of normalized p-p70-S6K: normalized beta-actin for CLL with or without BCAT1 expression before and after anti-IgM treatment. Mann-Whitney test (unpaired, nonparametric), ***p < 0.001. sIgM surface immunoglobulin M, CLL Chronic Lymphocytic Leukemia, BCAT1 branched chain amino acid transferase 1, CLL FISH Chronic Lymphocytic Leukemia fluorescence in-situ hybridization, IGHV immunoglobulin heavy chain variable, S6K S6 protein kinase.
Next, we pretreated N = 4 purified CLL each that expressed BCAT1 or were BCAT1 null with the BCAT1 selective inhibitor compound #2 at 100 µM (a concentration previously studied via metabolomic analyses by McBrayer et al.) [13] for 1 h and repeated the above experiments. Under these conditions, the elevated BCR-induced MTORC1 activation was abolished directly implicating the BCAT1 catalytic activity and one of the BCAT1 metabolites in MTORC1 regulation (Supplementary Fig. 4B–D).
CLL-derived cell lines require BCAT1 for optimal growthWe generated isogenic CLL-derived cell lines (HG3, PGA1 and C1; see methods) via CRISPR-Cas9 targeting of BCAT1, or BCAT2, or in parallel the control locus AAVS. We confirmed complete loss of BCAT1 or BCAT2 expression via immunoblotting in pools of targeted cells (Fig. 3E, J) and used these cell pools for subsequent experiments.
Fig. 3: BCAT1 expression is required for optimal growth of CLL-derived cell lines.
The BCAT1 or BCAT2 gene or the AAVS locus (control) were targeted in the CLL-derived cell lines C1, PGA1 and HG3, with a pool of four specific guides using pLENTI-CRISPRv2-Cas9 followed by puromycin selection and verification via immunoblotting that resulting cell pools lacked BCAT1 or BCAT2 expression expression E, J. The BCAT1 or BCAT2 targeted and AAVS control cells were grown in fully supplemented RPMI1640 media or RPMI1640 media with 25% of BCAA. A, C 12-day cell growth curves (y-axis log2 scale), Black: AAVS (control) targeted cells; Red: BCAT1 knock-out cells. B, D cell counts on day 12 (y-axis log2 scale). Two-Way ANOVA with the Holm-Šídák test, *p < 0.05; **p < 0.01; ***p < 0.001. F, H 12-day cell growth curves (y-axis log2 scale), Black: AAVS (control) targeted cells; Red: BCAT2 knock-out cells. G, I cell counts on day 12 (y-axis log2 scale). Two-Way ANOVA with the Holm-Šídák test, *p < 0.05; **p < 0.01. BCAT1 branched chain amino acid transferase 1, BCAT2 branched chain amino acid transferase 2, CLL: Chronic Lymphocytic Leukemia, AAVS adeno-associated virus site, CRISPR clustered regularly interspaced short palindromic repeats, BCAA branched chain amino acid.
Next, we tested the requirement for BCAT1 expression for the growth of these cell pools. We cultured BCAT1 and AAVS (controls) targeted cell pools in fully supplemented RPMI1640 media or alternatively in media with experimentally reduced (25%) BCAA concentrations. We measured substantially and significantly slower cell growth in the BCAT1 null pools thus uncovering a physiological advantage conferred on CLL-derived cells by aberrant BCAT1 expression (Fig. 3A–D; see also CLL patient survival data below).
We performed similar longitudinal growth assays for pools of the BCAT2 disrupted cell lines HG3, PGA1 and C1 compared with AAVS targeted controls. We detected a reduction in cell growth at full amino acid concentrations but not at 25% BCAA (Fig. 3F–I). Of note, the loss of BCAT2 expression resulted in compensatory upregulation of BCAT1 expression (Fig. 3J) in these cell lines that likely rescued some of the growth defects.
Prompted by the latter finding, we correlated the expression of BCAT1 with BCAT2 in the 205 primary CLL discussed above. We detected no significant correlation and no upregulation of BCAT1 preferentially in BCAT2 low CLL (Supplementary Fig. 2B).
Next, we tested whether BCAT1 expression augments MTOR activation in HG3, PGA1 and C1 pools targeted for BCAT1 or AAVS (control). We measured a substantially higher induction of p-S6K phosphorylation following BCR crosslinking in CRISPR-Cas9 targeted control (AAVS) cell pools that express BCAT1, when compared with CRISPR-Cas9 BCAT1 targeted null cell pools (Supplementary Fig. 5A). In contrast, BCR crosslinking in CRISPR-Cas9 BCAT2 targeted HG3, PGA1 and C1 pools resulted in augmented p-S6K phosphorylation (Supplementary Fig. 5B), which in part may be due to the measured upregulation of BCAT1 in these cell lines (Supplementary Fig. 5C, D).
Results from untargeted steady-state metabolomics in purified primary CLL cells that expressed high BCAT1 or were BCAT1 nullThe BCAT1 enzyme catalyzes the conversion of BCAAs (leucine, isoleucine, and valine) to branched chain keto amino acid derivatives (α-ketoisocaproate, α-keto-β-methylvalerate, and α-ketoisovalerate), which are further metabolized through intermediates to succinyl-CoA or acetyl-CoA, the later a substrate for a variety of catabolic and anabolic reaction pathways. In the forward reaction, α-Ketoglutarate (αKG) is consumed (but regenerated in the TCA cycle), and glutamate produced. The BCAT1 reaction is bi-directional however and thus can also generate Leu, Ile and Val and α-ketoglutarate from the respective branched-chain keto acids and glutamate, which is generated via glutamine deamination.
We performed untargeted small molecule metabolomics on CD19+ purified CLL cells from four CLL samples each that expressed high BCAT1 or were BCAT1 null. The metabolomic analyses identified ~100 known metabolites directly and another ~1200 with high mass accuracy, including an assigned chemical formula and another ~4900 unknown metabolites. The data were corrected for the total mass of recovered metabolites, which was 1.26-fold higher in the combined BCAT1 null CLL samples compared with the BCAT1 expressing samples.
Focusing initially on the BCAAs, leucine levels were higher at 1.21-fold, isoleucine at 1.24-fold and valine at 1.36-fold in BCAT1 high versus null CLL. Multiple di-peptides (e.g., Ile-Val, Leu-Pro, and others) containing BCAA were also higher in BCAT1 expressing CLL (see Fig. 4 and Supplementary Table 4). The BCAT1 forward reaction product glutamate was largely unchanged (0.96-fold). A few N-acetyl-amino acids, like N-acetyl-alanine (1.87-fold), N-acetyl-glutamic acid (1.69-fold), or N-acetyl-methionine (1.77-fold), which are breakdown products of N-acetylated proteins were higher in the BCAT1 high CLL samples, but the biological significance of this finding is unclear. There were many other less well characterized metabolites with differential expression although these were not easily directly linked to BCAA metabolism.
Fig. 4: Results from untargeted metabolomics of primary human CLL with BCAT1 expression or lack of BCAT1 expression (N = 4 each).
Primary human CLL cells were purified via Miltenyi column-based depletion of CD3+ T cells and CD14+ macrophages, cultured in RPMI1640 media with 10% FCS and pellets extracted with ice cold Methanol. Organic cell extracts were analyzed via mass spectrometry as detailed in the methods section. Displayed is the mean and individual measurements of selected aminoacids, dipeptides and N-acetylaminoacids in BCAT1 expressing versus absent CLL cases. The mean for metabolite measurements in CLL with absent BCAT1 expression was indexed at 1. Y-axis linear, dimension less. Mann-Whitney test (unpaired, nonparametric), *p < 0.05. BCAT1 branched chain amino acid transferase 1, BCAT2 branched chain amino acid transferase 2, CLL: Chronic Lymphocytic Leukemia.
Results from heavy isotope 13C-Leu metabolic labeling and tracings into TCA cycle intermediates in primary human CLLThe metabolism of Leu and Ile by BCAT1 in the cytoplasm and BCAT2 in mitochondria results in the generation of acetyl-CoA, which can be further metabolized via the tricarboxylic acid (TCA) cycle located in mitochondria (Fig. 5A). Alternatively, acetyl-CoA can be converted to various fatty acids or bioactive lipids. We employed heavy isotope labeled 13C-Leu to trace Leu metabolites via mass spectrometry in primary CLL cells. The 13C-Leu was added to RPMI media at equimolar concentrations to unlabeled Leu and cells incubated for 4 h. The CLL cells were selected to express high BCAT1 or no BCAT1 protein and aliquots of BCAT1 high cells were pre-incubated with the BCAT1 selective inhibitor compound #2.
Fig. 5: Aberrant BCAT1 expression accelerates primary CLL cell leucine consumption but does not change leucine conversion to TCA cycle intermediates.
Primary CLL cells aberrantly expressing BCAT1 or BCAT1 null were purified via Miltenyi column-based depletion of CD3 (T-cells) and CD14 (macrophages) and subsequently incubated in serum supplemented RPMI medium in the presence of 13C-Leu added at equimolar amounts to unlabeled Leu for 4 h in three separate experiments. An aliquot of BCAT1 expressing CLL cells was pre-incubated with 100 µM of the BCAT1 inhibitor compound #2. Cell pellets were prepared for mass spectrometric tracing as described in the methods. A Simplified schema of the BCAT1 reaction, carbon tracing and link to downstream TCA cycle intermediates. B Residual M + 5 leucine (input) in CLL cells after 4 h of cell culture. Increased Leu consumption in BCAT1 high cells is evident and is reversed by pre-incubation with a BCAT1 inhibitor. C–G The fractional 13C labeling of each indicated metabolite. One-Way ANOVA with the Dunnett test, **p < 0.01; ***p < 0.001. BCAT1 branched chain amino acid transferase 1, BCAT2 branched chain amino acid transferase 2, CLL: Chronic Lymphocytic Leukemia, TCA tricarboxylic acid cycle, Leu Leucine, ANOVA Analysis of Variance.
In preliminary studies using 13C-BCAA we noted that primary CLL cells without further metabolic activation were avid consumers of BCAA. We also tested anti-IgM and IL4 stimulation of CLL cells but detected no increased fractional TCA cycle metabolite labeling [23]. Using a labeling time of 4 h, we conducted three separate experiments using CLL samples expressing BCAT1 or not and carrying either del17p/TP53mut or trisomy 12. We measured higher fractional loss (consumption) of 13C-Leu in BCAT1 high CLL cells as compared with BCAT1 absent CLL or CLL with BCAT1 high expression that were pre-incubated with the BCAT1 inhibitor (Fig. 5B). These data show that BCAT1 contributed to Leu metabolism in BCAT1 high CLL. The labeling of glutmate was similar in BCAT1 expressing versus BCAT1 null CLL (Fig. 5C). We achieved substantial fractional labeling (5–10%) of the TCA cycle intermediates citrate, malate, succinate, and fumarate indicating highly active cellular uptake of Leu and metabolism via TCA cycle in primary CLL (Fig. 5D–G). However, the TCA labeling was not dependent on aberrant BCAT1 expression, as we measured similar fractional labeling in all three CLL groups (BCAT1-hi versus BCAT1 absent cells versus BCAT1-hi cells + BCAT1 inhibitor).
Thus, Leu flux into the TCA cycle in primary CLL cells was high indicating that BCAA are heavily metabolized by CLL cells. The expression of BCAT1 contributed to cytosolic consumption of labeled Leu but the flux of Leu into the TCA cycle was predominantly BCAT2 mediated.
Results from heavy isotope 15N-Leu metabolic labeling and tracings in primary human CLLIn three separate experiments, we labeled purified primary human CLL (BCAT1 high versus absent) with 15N-Leu for 1 h, 2 h or 4 h followed by mass spec-based quantification of 15N-containing molecules. Similar to the findings for 13C-Leu labeled CLL, we detected less 15N-Leu in CLL with BCAT1 high expression when compared with BCAT1 absent CLL cases confirming BCAT1 mediated Leu metabolism (Fig. 6A, B). The pre-incubation with the BCAT1 inhibitor partially restored this difference indicating BCAT1 dependent Leu consumption. We measured substantial 15N labeling of Ile and Val starting at 1 h and increasing with time in both BCAT1 expressing and null cells (Fig. 6C, D) reflective of a BCAT2 effect with a fractional contributions by BCAT1, together indicative of bidirectional BCAT1/2 reactions in CLL (15N-Leu is initially converted to 15N-glutamate resulting in 15N-Leu loss, followed by a reverse reaction/conversion of 15N-Glutamate to 15N-Leu, 15N-Ile, and 15N-Val). We measured substantial labeling of Alanine and Aspartate because of the actions of GOT2 and GPT2 in mitochondria confirming rapid conversion of Leu into these amino acids (Fig. 6E–G). We detected little labeling into glutamine indicating lack of conversion from glutamate in glutamine rich media (Fig. 6H).
Fig. 6: The BCAT1/2 reaction in primary CLL cells is rapidly bi-directional.
Primary CLL aberrantly expressing BCAT1 or BCAT1 null were purified via Miltenyi column-based depletion of CD3 (T-cells) and CD14 (macrophages) and subsequently incubated in serum supplemented RPMI medium in the presence of 15N-Leu added at equimolar amounts to unlabeled Leu for 1 h, 2 h or 4 h in three separate experiments. An aliquot of BCAT1 expressing CLL cells was pre-incubated with 100 µM of the BCAT1 inhibitor compound #2. Cell pellets were prepared for mass spectrometric tracing as described in the methods. A Simplified schema of the bi-directional BCAT1 reaction. B–H The fraction of each indicated metabolite labeled by 15N at various time points. Two-Way ANOVA with the Holm-Šídák test, *p < 0.05; **p < 0.01; ***p < 0.001. BCAT1 branched chain amino acid transferase 1, BCAT2 branched chain amino acid transferase 2, CLL: Chronic Lymphocytic Leukemia, Leu Leucine, ANOVA Analysis of Variance.
These data identify bi-directional BCAT1/2 reactions in CLL cells, resulting in the generation and interchange of multiple small molecules with MTOR activating properties. It is likely that the reversibility of the BCAT1/2 reaction confers metabolic flexibility onto affected CLL cells allowing for adjustments to the ambient nutritional milieu.
CLL with high aberrant BCAT1 protein expression have reduced sensitivity to Venetoclax-induced apoptosisWe treated purified CLL cells from four patients each that were BCAT1 protein expressing high or absent and matched for genomic characteristics with escalating Venetoclax concentrations ex vivo for 24 h and measured cell viability using CellTiter-Glo® based ATP assays (Supplementary Fig. 6A, B). We found that CLL with high BCAT1 protein expression were less sensitive to Venetoclax induced apoptosis than CLL with absent BCAT1 expression (IC50 values of ~3 nM and 1 nM, respectively; p = 0.03), and, of greater importance, a substantially higher fraction of BCAT1 high CLL survived even at the highest Venetoclax dose tested (cell fraction alive at 1 µM of Venetoclax of 19% and 7%, respectively; p < 0.001).
We treated isogenic cell line pools derived from HG3, C1 and PGA1 cells carrying BCAT1 disruption or AAVS disruption (controls) in two separate experiments per line (N = 6 total) with escalating doses of Venetoclax over 24 h, followed by CellTiter-Glo® based ATP cell viability assays (Fig. 6C–F). We found that BCAT1 expression associated with substantially reduced sensitivity to Venetoclax mediated cell death (pooled IC50 values of ~18 nM and 2 nM, respectively, p < 0.01). We measured the expression of anti apoptotic BCL2 family members in all lines via immunoblotting and detected no differences in expression (Fig. 6G).
We performed similar experiments in the BCAT2 disrupted HG3, C1 and PGA1 cell pools and detected no effect of BCAT2 expression on Venetoclax dose responses (Supplementary Fig. 7).
BCAT1 expression in CLL associates with substantially shortened patient survivalMultiple lines of experimental evidence suggested that BCAT1 facilitates CLL growth thus contributing to the progressive and aggressive clinical phenotypes displayed by CLL with del17p/TP53mut status or trisomy 12. To determine the effects of BCAT1 expression on CLL patients’ survival, we performed Kaplan Meier survival analyses on a CLL cohort with long clinical follow-up using overall survival as the main clinical outcome endpoint. We categorized patients in the CLL cohort into two groups: (i) BCAT1 protein expressing or (ii) BCAT1 protein absent (see Fig. 1B for examples of both patient categories). Using such a dichotomization there were 33% (53/160) of untreated CLL at CLL blood sample procurement with BCAT1 protein expression out of a total of 160 CLL with available BCAT1 protein expression data.
As can be seen in Fig. 7A, CLL patients with BCAT1 expression had a substantially worse clinical outcome than BCAT1 null CLL (median survival of 125 months for BCAT1 protein expressors versus 296 months for BCAT1 null CLL; p < 0.0001). Of further interest, the adverse survival effect of BCAT1 expression remained significant even after exclusion of all patients carrying TP53 mutations (median survival of 193 months for BCAT1 protein expressors versus 296 months for BCAT1 null CLL; p < 0.003; Fig. 7B). Overall, the adverse effect of BCAT1 expression was comparable to other well-defined adverse CLL biomarkers, including an unmutated IGHV status, elevated SNP 6.0 array based genomic complexity or selected FISH categories. The worst outcome was measured for CLL patients with TP53 mutations Fig. 7C–F.
Fig. 7: Reduced overall survival of CLL patients aberrantly expressing BCAT1 (Kaplan-Meier plots).
Survival analysis of CLL patients untreated at the time of enrollment stratified by biomarkers. Total number of patients with available data and positive or negative marker measurements is indicated. A BCAT1:BCAT2 pooled immunoblot protein ratios (any detectable BCAT1 protein in orange). B Same as in (A) but exclusive of TP53 mutated CLL. C TP53 mutated versus WT. D IGHV mutation status MUT versus UM. E. CLL interphase FISH findings prioritized 17p > 11q > t12 > 13q14>normal. F SNP 6.0 array based genomic acquired copy number aberrations (complexity) per case. BCAT1 branched chain amino acid transferase 1, BCAT2 branched chain amino acid transferase 2, CLL: Chronic Lymphocytic Leukemia, CLL FISH Chronic Lymphocytic Leukemia fluorescence in-situ hybridization, IGHV immunoglobulin heavy chain variable, MUT mutated, UM unmutated SNP single nucleotide polymorphism.
Next, we performed multi variate analyses incorporating BCAT1 protein expression (a BCAT1/BCAT2 protein ratio >0) together with IGHV status into a base model and alternatively added the adverse genomic traits TP53 mutations or SNP 6.0 array based elevated genomic complexity ( ≥ 3 acquired genomic copy number aberrations) into the models. Multi-variable survival analyses were conducted using proportional hazards regression (Cox) models, using the hazard ratio for each prognostic factor to assess its relationship to survival when controlling for other factors. BCAT1 expression independently associated with significantly elevated hazard ratios for death in both models (Model A: BCAT1 protein + HR 1.76, p = 0.05, IGHV UM HR 1.66, p = 0.08, TP53mut HR 3.55, p < 0.001. Model B: BCAT1 protein + HR 1.99, p = 0.04, IGHV UM HR 1.53, p = 0.17, SNP 6.0 aCNA ≥ 3 (genomic complexity), HR 2.08, p = 0.09; also see Table S5 in Supplementary Methods and Results).
In contrast, the protein expression levels of BCAT2 in CLL were not prognostic.
留言 (0)