
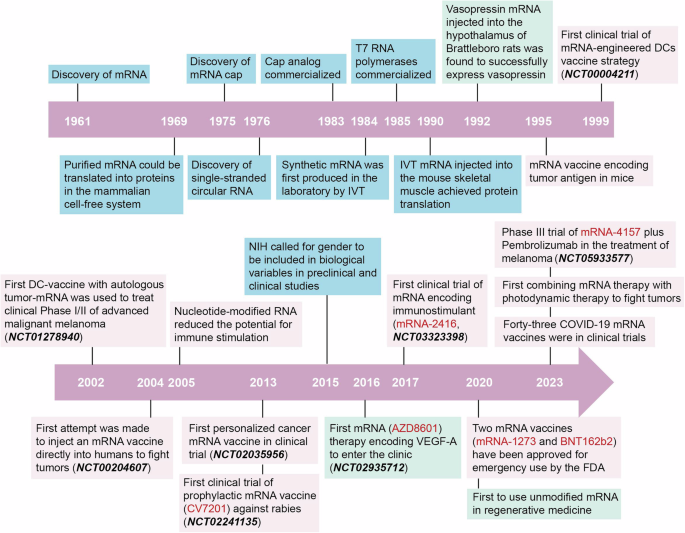
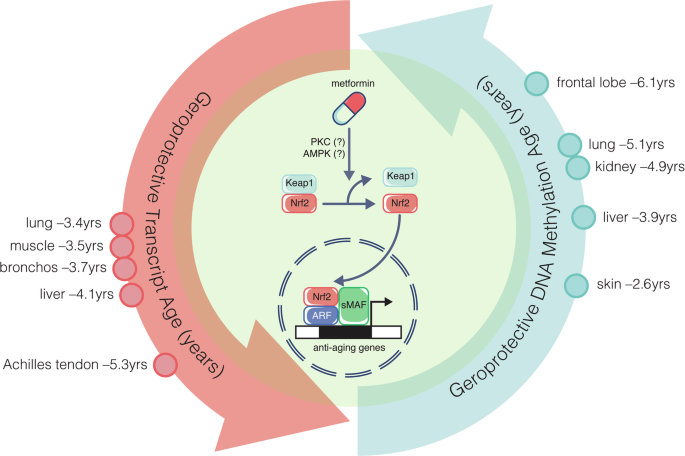
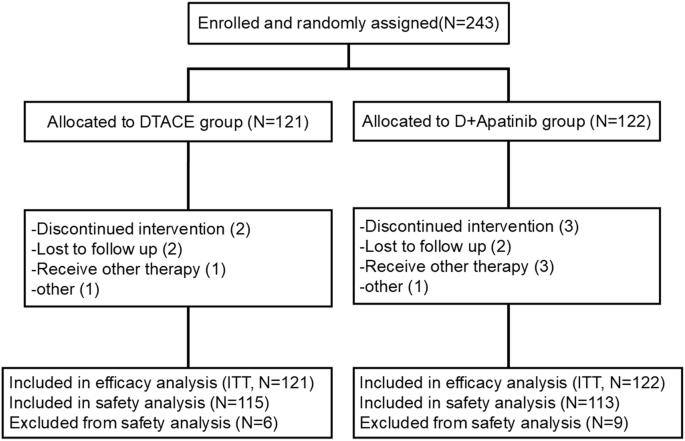
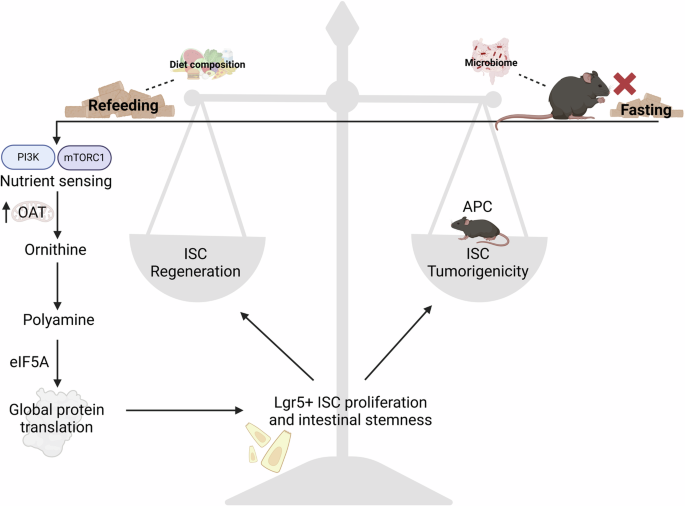
記住我
The study utilized GD2.28z, CD19.28z (19.28z), and CD19.4-1BBz (19.4-1BBz) CARs (Supplementary Fig. 1a). CAR tonic signaling gradually decreased across these 3 CARs, resulting from the difference in their scFvs and costimulation domains.6,10 Untransduced T (UTD) and 19.4-1BBz CAR-T cells expressed fewer inhibitory receptors (PD-1, TIM-3, and LAG-3) and activation markers (CD25 and CD69) and contained fewer effector memory CAR-T cells compared to 19.28z CAR-T cells (Fig. 1a–d). Using UTD cells and 19.4-1BBz CAR-T cells as controls, we first added trametinib, cobimetinib, and binimetinib, respectively, to 19.28z CAR-T cell culture at concentrations approaching their peak blood concentration for 9 days.1,20 All 3 MEKIs increased naive, reduced effector memory, and inhibited the expression of inhibitory receptors and activation markers (Fig. 1a–d). Among them, trametinib was the most potent because it could achieve the comparable effect of cobimetinib and binimetinib at the lowest concentration (Fig. 1a–d). Thus, we chose trametinib for further research. Compared to 7.5 nM and 30 nM, at 15 nM, trametinib demonstrated optimal efficacy by mildly inhibiting CAR-T cell expansion while effectively suppressing the activation, exhaustion, apoptosis, differentiation, CD8 decline, and ERK phosphorylation of CAR-T cells (Fig. 1e–i and Supplementary Fig. 1b–g). The above impacts of trametinib were more significant as the treatment time increased (Supplementary Fig. 1b–g). The roles of trametinib in reducing expression of exhaustion and activation markers and differentiation of CD4 and CD8 single-positive CAR-T cells were the same as its roles on bulk CAR-T cells (Supplementary Fig. 2a–d). A 9-day pretreatment with trametinib significantly reduced the release of TNF-α, IFN-γ, IL2, GzmB, CCL3, and CXCL8 by CD4 19.28z cells (Supplementary Fig. 2e). In contrast, the same pretreatment slightly inhibited CD8 19.28z CAR-T cells from releasing TNF-α, GzmB, CCL3, and CXCL8 while enhancing their release of IL2 and IFN-γ (Supplementary Fig. 2f). Consistently, a 9-day pretreatment with trametinib impaired the cytotoxicity of CD4 19.28z CAR-T cells but enhanced the cytotoxicity of CD8 19.28z CAR-T cells, while having no impact on the cytotoxicity of bulk 19.28z CAR-T cells (Fig. 1j and Supplementary Fig. 1h).
Fig. 1
MEK inhibition restrains the exhaustion and differentiation of CAR-T cells elicited by antigen-independent tonic signaling. a, b Flow cytometric analysis of exhaustion (PD-1, TIM-3, and LAG-3) and activation markers (CD25 and CD69) of CAR-T cells on day 15 of ex-vivo culture. One representative donor’s histograms (a) are shown. Bar graphs (b) are pooled from 3 donors. c, d Differentiation state of CAR-T cells on day 15 of ex-vivo culture. One representative donor’s pseudocolor plots are shown (c). CD62L and CD45RO define the differentiation state of CAR-T cells as the following combinations: naive T cells (TN, CD62L + CD45RO-), central memory T cells (TCM, CD62L + CD45RO + ), effector memory T cells (TEM, CD62L-CD45RO + ) and effector T cells (TEFF, CD62L-CD45RO-). Bar graphs (d) are pooled from 3 donors. e WB evaluates pERK versus total ERK of 19.28z CAR-T cells on day 15 of ex-vivo culture. Representative of 3 donors. f The bar graphs show the expansion fold of 19.28z CAR-T cells. Data are pooled from 3 donors. g Representative histogram of CellTrace Violet showing the cell proliferation state of 19.28z CAR-T cells after a six-day treatment with trametinib. N = 2 donors. h, i Quantification of apoptosis of 19.28z CAR T cells on day 15 of ex-vivo culture. One representative donor’s pseudocolor plots (h) are shown. Bar graphs (i) are pooled from 5 donors. j, k Cytotoxicity of CD4 and CD8 19.28z CAR-T cells (j), and CD4 and CD8 GD2.28z CAR-T cells (k) pre-treated with trametinib for 9 days. The assay was conducted in culture media without trametinib. Error bars represent means ± SD of triplicate wells. A representative donor from three donors. Error bars are means ± SEM and statistical test was paired one-way ANOVA with Dunnett’s multiple comparison test except (j, k). For (j, k), an unpaired student’s t-test was used. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, NS not significant, Tra trametinib, Cobi cobimetinib, Bini binimetinib, UTD untransduced T
To confirm whether the effects of MEKIs depend on the specific scFv and costimulation domain, we treated 19.4-1BBz and exhaustion-susceptible GD2.28z CAR-T cells6,10,22 with trametinib. Consistently, trametinib prevented them from activation, exhaustion, apoptosis, and differentiation (Supplementary Fig. 3a–j). Interestingly, trametinib mildly inhibited cell proliferation in the less-exhausted 19.28z and 19.4-1BBz CAR-T cells while slightly promoting proliferation in the more exhaustion-prone GD2.28z CAR-T cells6,10,22 (Fig. 1f and Supplementary Fig. 3l). Consistently, cell trace violet showed that trametinib had a minimal effect on the cell division of these three types of CAR-T cells (Fig. 1g and Supplementary Fig. 3k). Both methods confirmed that trametinib had a mild effect on CAR-T cell proliferation. Likely due to the higher exhaustion level in GD2.28z CAR-T cells, trametinib’s anti-apoptotic effect resulted in a slight increase in their cell quantity during manufacturing.
In line with the effect on 19.28z CAR-T cells, a 9-day trametinib pretreatment decreased the effector function of CD4 GD2.28z CAR-T cells while boosting the effector function of CD8 GD2.28z CAR-T cells (Fig. 1k and Supplementary Fig. 2g, h). Notably, trametinib had a more pronounced positive impact on the effector function of CD8 GD2.28z CAR-T cells compared to 19.28z CAR-T cells (Fig. 1j, k and Supplementary Fig. 2f–h) and even promoted the cytotoxicity of bulk GD2.28z CAR-T cells (Supplementary Fig. 1i).
We further investigated the in-vivo efficacy of trametinib pretreated CAR-T cells using a leukemia xenograft model in NCG mice (Fig. 2a). Trametinib pretreated 19.28z CAR-T cells demonstrated superior anti-leukemia activity compared to DMSO pretreated counterparts. That was evidenced by lower leukemia burden, longer mice survival, improved CD8 CAR-T expansion, increased CAR-T infiltration in bone marrow, more effector memory CAR-T cells, fewer effector CAR-T cells, and diminished expression of exhaustion and activation markers (Fig. 2b–h). We obtained similar results in xenograft leukemia model treated with 19.4-1BBz CAR-T cells (Supplementary Fig. 4a–h).
Fig. 2
Trametinib pretreatment during ex-vivo manufacturing enhances the in-vivo efficacy of 19.28z CAR-T cells. a 1.5 or 2 or 3 × 106 19.28z CAR-T cells manufactured in the presence of trametinib 15 nM or DMSO for 9 days were infused to NCG mice intravenously (IV) 6 days after engraftment of 1 × 106 Nalm-6-GL leukemia cells in 3 independent experiments. b–d Tumor growth was monitored by bioluminescent imaging. One representative experiment is shown, and 1.5 * 106 CAR-T cells were administrated (UTD cells: n = 3; DMSO pre-treated CAR-T cells: n = 6; trametinib 15 nM pre-treated CAR-T cells: n = 6). Each dot in (c) and each curve in (d) represents one mouse. D, day. e Kaplan–Meier analysis of survival of mice. Data are pooled from 3 independent experiments (UTD cells: n = 7; DMSO pre-treated CAR-T cells: n = 13; trametinib 15 nM pre-treated CAR-T cells: n = 14). f–h The number (f), subset composition (g), and positive rate of exhaustion and activation markers (h) of CAR-T cells in the bone marrow 8 days after CAR-T infusion. Data in (f) are pooled from 2 independent experiments, with 1.5 * 106 and 2 * 106 CAR-T cells administered in each experiment, respectively. Data in (g, h) are from 1 representative experiment, with 1.5 * 106 CAR-T cells administered in (g) and 2 * 106 CAR-T cells administered in (h). Each dot represents one mouse. N = 5 or 9. Error bars are means ± SEM. Statistical tests were paired two-tailed Wilcoxon test (c) and paired two-tailed student’s t-test (f–h). Survival curves were compared using a log-rank Mantel-Cox test. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. D day, Tra trametinib, UTD untransduced T
Overall, regardless of scFv and costimulation domain, trametinib treatment during ex-vivo manufacturing prevented CAR-T cells from overactivation, exhaustion, apoptosis, differentiation, and CD8 decline driven by tonic signaling, thus enhancing CAR-T cells’ in-vivo potency.
MEKIs modify CAR-T cell transcriptome during ex-vivo manufacturingTo explore the molecular mechanism underlying the enhanced persistence of trametinib-treated CAR-T cells, we performed transcriptional profiling of DMSO-treated UTD cells and DMSO or trametinib-treated 19.28z CAR-T cells (without antigen or tumor cell stimulation). 2128 DEGs among the three groups were identified, with 544 DEGs shared between CAR-T + DMSO vs. UTD + DMSO and CAR-T + DMSO vs. CAR-T + trametinib (Supplementary Fig. 5a).
GSEA showed DMSO-treated group downregulated naive-/memory-related genes and upregulated Myc- and pERK-regulated genes, and genes involved in T cell activation/effector/exhaustion, AP-1/NFAT/mTORC1 pathway, cytokine signaling and glycolysis/amino acid metabolism, compared to UTD and trametinib-treated group (Fig. 3a, b and Supplementary Fig. 5b, c). Consistently, KEGG pathways enriched in the DMSO-treated group relative to UTD and the trametinib-treated group included cytokine−cytokine receptor interaction, chemokine signaling pathway, and amino acids biosynthesis (Fig. 3c). The results of GSEA and KEGG were in accordance with the phenotypic change caused by trametinib during CAR-T manufacturing.
Fig. 3
MEK inhibition modifies the gene-expression profile of CAR-T cells during ex-vivo manufacturing. a Normalized enrichment score (NES) of significantly up- or downregulated gene sets in UTD + DMSO versus 19.28z CAR-T + DMSO (left panel) and 19.28z CAR-T + trametinib 15 nM versus 19.28z CAR-T + DMSO (right panel) as determined by GSEA using the MSigDB C7 gene sets. For all pathways, the false discovery rate (FDR) < 0.05. b Representative GSEA enrichment plot. c KEGG pathway enrichment analysis of differentially expressed genes (DEGs) among samples. d Heat map demonstrating the expression profiles of selected DEGs (FDR < 0.05) among the three groups. The AP-1 TFs are denoted in red. e Normalized RNA-seq counts of selected genes. Error bars are means ± SEM. Statistical test was unpaired one-way ANOVA with Dunnett’s multiple comparison test. For all analyses, n = 3 per group. ***P < 0.001, ****P < 0.0001. UTD untransduced T. C 19.28z CAR-T, D DMSO, Tra trametinib,TF transcription factor
Next, we examined specific gene alterations during the process. Naive-/memory-related TFs (TCF7, etc.)23 and MXD4, a suppressor of MYC,24 were upregulated with KIT and IL7R acquisition, whereas effector/exhaustion-related TFs (EGR1, TOX2 and NR4A1, etc.),25,26 AP-1 TFs (FOS and JUNB, etc.)22 and anergy-related TFs (EGR2, etc.)27 were downregulated with immune checkpoints (LAG3, etc.), effector molecules (CCL1, etc.) and pro-apoptotic genes (FAM162A, etc.) reduction in UTD and trametinib-treated group relative to DMSO-treated group (Fig. 3d, e, Supplementary Fig. 5d and Supplementary Table 1). Additionally, trametinib downregulated MAP kinase suppressors (DUSP1, etc.) and enzymes implicated in glycolysis (HK2, etc.) and amino acid metabolism (GPT2, etc.). The former probably served as a feedback mechanism,28 while the latter implied a more quiescent metabolic state in CAR-T cells following MEK inhibition (Fig. 3d, e, supplementary Fig. 5d and Supplementary Table 1).
Finally, we investigated how trametinib affects the transcriptional profile of 19.4-1BBz CAR-T cells during their manufacturing process. Trametinib treatment led to 126 shared DEGs in 19.4-1BBz and 19.28z CAR-T cells. They shared downregulated genes, including exhaustion-related TFs (EGR1, TOX2 and NR4A1),25,26 AP-1 TFs (FOS and FOSL1),22 anergy-related TFs (EGR2 and EGR3),27 an immune checkpoint (CD276), effector molecules (CCL1, CCL3 and CXCL8, etc), a T cell activation marker (TNFRSF18) and negative regulators of MAP kinase (DUSP2, DUSP4, SPRED2) (Supplementary Fig. 5e). Of note, the shared upregulated genes included the stemness-related molecule KIT (Supplementary Fig. 5e)
MEKIs limit antigen-induced CAR-T cell exhaustion and differentiationTo evaluate trametinib’s protection against antigen-induced CAR-T cell exhaustion and differentiation, we conducted three in-vitro assays with 19.28z CAR-T cells (Fig. 4a and “Methods”). Whether added at the beginning of coculture or after Nalm-6 cells eradication, trametinib can reduce exhaustion, activation, and ERK phosphorylation, prevent differentiation, apoptosis, and CD8 decline, and promote proliferation of CAR-T cells (Fig. 4b–k and Supplementary Fig. 6a–i). CD25 MFI was lowered, but the change in CD25 positive rate pooled from 5 donors was statistically insignificant (Fig. 4b, c and Supplementary Fig. 6a, b). Again, 15 nM was the optimal concentration of trametinib for its relatively maximum effects in promoting CAR-T expansion and ameliorating exhaustion and differentiation (Fig. 4b–e, g–i and Supplementary Fig. 6a–g).
Fig. 4
MEK inhibition limits the exhaustion and differentiation of CAR-T cells triggered by target antigen. a Experimental design for target antigen stimulation. b, c Flow cytometric analysis of exhaustion and activation markers. The histograms (b) of one representative donor are shown. Bar graphs (c) are pooled from 4 to 8 donors. d, e Differentiation state of 19.28z CAR-T cells. Pseudocolor plots (d) of one representative donor are shown. Bar graphs (e) are pooled from 5 donors. (f) WB evaluates pERK versus total ERK. Representative of 3 donors. g, i Quantification of apoptosis of 19.28z CAR T cells. The bar graphs (g) are pooled from 6 donors and the pseudocolor plots (i) of one representative donor are shown. (h) The bar graphs show the expansion fold of 19.28z CAR-T cells. Data are pooled from 3 donors. j, k CD8 and CD4 composition in 19.28z CAR-T cells. Pseudocolor plots (j) of one representative donor are shown. Bar graphs (k) are pooled from 4 donors. l, m Cytotoxicity of 19.28z CAR-T cells cocultured with Nalm-6-GL cells for 18 h in the culture medium free of trametinib or DMSO. Error bars represent means ± SD of triplicate wells. A representative donor from four donors. n Proliferation curve of 19.28z CAR-T cells. Arrows denoted the time points when Nalm-6 cells were added. Data are pooled from 5 donors. o Cytotoxicity of 19.28z CAR-T against Nalm-6-GL cells (left panel) and GD2.28z CAR-T against 143B-GL cells (right panel) in the culture medium with trametinib or DMSO. Error bars represent means ± SD of triplicate wells. (19.28z: n = 4 donors; GD2.28z: n = 3 donors). The data in (b–k) were produced using CAR-T cells treated by protocols indicated in the upper panel of Fig. 4a. Error bars are means ± SEM unless indicated otherwise. Statistical tests were paired (c, e, g, h, k) or unpaired (m) one-way ANOVA with Dunnett’s multiple comparison test, and paired (n) or unpaired (o) students’ t test. Tests were two-tailed. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.NS not significant. C 19.28z CAR-T, D DMSO, Tra trametinib, N6 Nalm-6, UTD untransduced T
Probably due to insufficient one-time antigen stimulation, trametinib didn’t enhance CAR-T cells’ cytotoxicity after it was washed out (Supplementary Fig. 6j, k). Thus, we designed the repetitive antigen stimulation assay (Fig. 4a, the lowest panel). Expectedly, trametinib reduced CAR-T cells’ functional exhaustion and growth arrest under this situation (Fig. 4l–n).
Previous cytotoxicity assays used the culture media without trametinib. Given trametinib’s impact on reducing CAR-T cell activation, we investigated its influence on CAR-T cell effector function. 19.28z and GD2.28z CAR-T cells were cocultured with Nalm-6-GL and 143B-GL cells, respectively, in culture media containing DMSO or trametinib. Trametinib reduced the release of TNF-α, IFN-γ, IL2, GzmB, CCL3, and CXCL8 in both 19.28z and GD2.28z CAR-T cells, regardless of the CD4 and CD8 subsets (Supplementary Fig. 6l–o). However, trametinib didn’t impair 19.28z CAR-T cells’ cytotoxicity against Nalm-6 cells (Fig. 4o). Furthermore, trametinib showed direct killing against 143B cells, and when combined with GD2.28z CAR-T cells, trametinib and GD2.28z CAR-T cells showed synergistic killing in vitro (Fig. 4o).
We next investigated if trametinib in-vivo administration prevented CAR-T cells from exhaustion and differentiation. To exclude MEKIs’ potential suppression on 143B cells, we injected 143B-bearing mice with UTD cells or GD2.28z CAR-T cells and orally administrated mice with trametinib or vehicle (Fig. 5a). Likely due to the differences between in-vitro and in-vivo circumstances, unlike the in-vitro killing assay, after tumor engraftment, administrating trametinib alone following our protocol from day 8 to 18 didn’t mitigate tumor progression (Fig. 5b–e). Before day 15, CAR-T cells alone only slowed tumor growth in 1 out of 10 mice and didn’t significantly increase mice survival (Fig. 5b, c). In contrast, the combination of trametinib and CAR-T cells significantly delayed tumor progression and extended mice survival, with one mouse having no visible tumor at euthanasia (Fig. 5b–g). Tumor analysis showed an increase in total and CD8 CAR-T cells, as well as the sum of naïve plus central memory plus effector memory CAR-T cells (Fig. 5h, i), with lower PD-1, TIM-3, and CD25 expression (Fig. 5j). Trametinib reduced PDL1 expression in tumor cells, consistent with prior research showing MEK-mediated RAS signaling increased PDL1 expression29 (Fig. 5k). To investigate the optimal combination protocols of MEKIs and CAR-T cells, we further compared pulsatile vs continuous MEK inhibition on CAR-T cells using CAR-T cells from another donor (Supplementary Fig. 7a). Again, trametinib alone didn’t mitigate tumor progression (Supplementary Fig. 7b–f). GD2.28z CAR-T cells from this donor slightly slowed tumor progression and minimally extended mice survival (Supplementary Fig. 7b–f). Conversely, continuous MEK inhibition on CAR-T cells notably delayed tumor progression and extended mice survival in all five mice (Supplementary Fig. 7b–f). Although there was no statistically significant difference in tumor burden and mice survival between continuous and pulsatile MEK inhibition, the pulsatile group exhibited trends of higher average tumor burden and shorter median survival (49.5 vs 53 days) compared to the continuous group, indicating continuous MEK inhibition is the optimal combination ways of MEKIs and CAR-T cells. Together, these data implied that under antigen stimulation, MEKIs reduced CAR-T cell exhaustion and differentiation both in vitro and in vivo.
Fig. 5
In-vivo administration of trametinib enhances the antitumor activities of CAR-T cells by offsetting CAR-T cells exhaustion and differentiation. a Mice treated with GD2.28z CAR-T cells or UTD cells were dosed with trametinib or vehicle once daily. b Kaplan–Meier analysis of survival of mice. Data are pooled from two independent experiments (UTD + vehicle: n = 3; UTD + trametinib: n = 3; GD2.28z + vehicle: n = 10; GD2.28z + trametinib: n = 10). c, e Tumor growth was monitored by bioluminescent imaging. Each curve in (c) represents one mouse. D day. d Growth curve of tumor size. Each line represents one mouse. The dotted lines indicate the endpoint where tumor volume reached 1.5 cm3, and the asterisks indicate mice died before reaching the endpoint. f Picture of tumors on day 19 after CAR-T infusion. The arrow indicates a tiny tumor, and the asterisk indicates a missing macroscopic tumor in one of the mice in GD2.28z + trametinib group. N = 5 mice per group. One of two independent experiments is shown. g Bar plots of tumor volume and tumor weight of mice from (f). N = 5 mice per group. h Tumor-infiltrating total GD2.28z, CD8 GD2.28z, and CD4 GD2.28z CAR-T cell counts normalized to tumor weight of mice from (g). GD2.28z + vehicle: n = 5. GD2.28z + trametinib: n = 4 for one mouse in GD2.28z CAR-T + trametinib group didn’t have a macroscopic tumor. i The percentage of CD8 and CD4, along with the total percentage of naïve plus central memory plus effector memory in tumor-infiltrating GD2.28z CAR-T cells of mice from (g). j The positive rate of PD-1, TIM-3, and CD25 in tumor-infiltrating GD2.28z CAR-T cells of mice from (g). k The MFI of PDL1 in tumor cells of mice from (g). Error bars are means ± SEM unless indicated otherwise. Statistical tests were unpaired (g, i, j, k) students’ t test, Mann–Whitney test (h), and log-rank Mantel-Cox test (b). All tests were two-tailed. *P < 0.05, **P < 0.01, ***P < 0.001, NS not significant, Tra trametinib, UTD untransduced T, N naive, CM central memory, EM effector memory
MEKIs alter antigen-stimulated CAR-T cells transcriptome to a naive-/memory-like stateWe analyzed the transcriptome of Nalm-6-stimulated CAR-T cells treated with DMSO or trametinib and CAR-T cells without Nalm-6 stimulation (defined as “resting” below). After antigen stimulation, the transcriptomic differences among the three groups differed from the “resting state.” 5337 DEGs among the three groups were identified, with 1070 DEGs shared between CAR-T + Nalm-6 + DMSO vs. CAR-T + DMSO and CAR-T + Nalm-6 + DMSO vs. CAR-T + Nalm-6 + trametinib (Supplementary Fig. 8a), suggesting the alteration of MEKIs on CAR-T cell transcriptome is magnified under antigen exposure (Supplementary Figs. 5a, 8a). Of the 1576 DEGs in CAR-T + Nalm-6 + DMSO vs. CAR-T + Nalm-6 + trametinib, 270 overlapped with CAR-T + DMSO vs. CAR-T + trametinib (Supplementary Fig. 8b). Trametinib-treated groups shared downregulated genes, including exhaustion/anergy-related and AP-1 TFs (FOS and JUNB, etc.), effector molecules, glycolysis/amino acid metabolism-related enzymes, and pro-apoptotic genes. They also shared upregulated genes, including naive-/memory-related genes, KIT, and MXD4 (Supplementary Fig. 8b).
Apart from the differential pathways found in the “resting state”, GSEA revealed Nalm-6-stimulated group upregulated several novel pathways, including oxidative phosphorylation, fatty acid/nucleotide/polyamine metabolism, and apoptosis compared to trametinib-treated and resting groups (Fig. 6a, b and Supplementary Fig. 8c, d). The newly-found KEGG pathways enriched in Nalm-6-stimulated group relative to trametinib-treated and resting groups included T cell receptor signaling pathway and fatty acid metabolism (Fig. 6c). Given the similarity between CAR signaling pathway and TCR signaling pathway,19,30 these results confirmed that under antigen stimulation, MEK inhibition not only suppresses genes associated with the CAR signaling pathway but also appears to attenuate the metabolic activities of CAR-T cells.
Fig. 6
MEK inhibition alters the gene profile of antigen-stimulated CAR-T cells to a naive-/memory-like state. (a) NES of significantly up- or downregulated gene sets in 19.28z + DMSO versus 19.28z + Nalm-6 + DMSO (left panel) and 19.28z + Nalm-6 + trametinib 15 nM versus 19.28z + Nalm-6 + DMSO (right panel), as determined by GSEA using the MSigDB C7 gene, sets. For all pathways, the FDR q < 0.05. b Representative GSEA enrichment plot. c KEGG pathway enrichment analysis of DEGs among samples. d Normalized RNA-seq counts of selected genes. Statistical test was unpaired two-tailed one-way ANOVA with Dunnett’s multiple comparison test. Error bars are means ± SEM. e Heat map showing the expression profiles of selected DEGs (FDR < 0.05) among the three groups. The AP-1 TFs are denoted in red. For all analyses, n = 3 per group. C 19.28z CAR-T, N6 Nalm-6, D DMSO, Tra trametinib., TF transcription factor
Besides the naïve/memory-related and exhaustion/effector/anergy/glycolysis/amino acid metabolism/apoptosis-related DEGs found in “resting state”, a suppressor of PI3K/Akt/mTOR pathway, PIK3IP131 was upregulated, whereas genes participating in mitochondria oxidative phosphorylation (NDUFAF2, etc.), polyamine synthesis (ODC1, etc), nucleotide metabolism (AK4, etc.), and translation (EIF4A1, etc.) were downregulated in trametinib-treated and resting groups relative to DMSO-treated group, indicating a more dramatic metabolic alteration of MEKIs to antigen-stimulated CAR-T cells (Fig. 6d, e, Supplementary Fig. 8e and Supplementary Table 2). Consistent with upregulation of PIK3IP1 and downregulation of “mTORC1 signaling” gene sets, trametinib downregulated genes (PIK3CB, etc.)32 in PI3K/Akt/mTOR pathway, implying a crosstalk between MAPK and PI3K/Akt/mTOR pathway33 (Fig. 6e and supplementary Table 2).
Single-cell transcriptome analysis of CAR-T cells after MEK inhibitionTo elucidate MEKIs’ impacts on individual CAR-T cell transcriptome, we performed single-cell transcriptome analysis on flow-sorted resting CAR-T and Nalm-6-stimulated CAR-T cells treated with DMSO or trametinib. DEGs were identified among the three groups, strongly correlating with bulk RNA-seq findings (Supplementary Fig. 9a). Next, eight clusters were identified and visualized by uniform manifold approximation and projection (UMAP) (Fig. 7a), with full names and identifying markers shown in Methods and Fig. 7b.
Fig. 7
Single-cell transcriptome analysis reveals an increase of memory CAR-T cells while a decrease of effector/exhausted CAR-T cells after MEK inhibition. a The UMAP visualization of 35469 cells from all the samples. 8 clusters are indicated by different colors. b Dot plot illustrating the expression of the marker genes in different clusters. c PAGA analysis shows the potential developmental connectivity among all eight clusters. d, e The UMAP and bar plots show the constitution of the eight clusters in the 3 samples. f, g The UMAP and bar plots show the relative frequency of 19.28z CAR-T cells in each phase of the cell cycle. h The violin plots depict the single-cell expression of the selected genes in each cluster of the three samples. C 19.28z CAR-T, N6 Nalm-6., D DMSO, Tra trametinib. Q, quiescent. Statistical test was a two-tailed Wilcoxon test. *P< 0.05, **P <0.01, ***P < 0.001, ns: not significant
Partition-based graph abstraction (PAGA)34 constructed developmental trajectories for these clusters, revealing that qCD4 TCM and qCD8 TCM were at the start of CD4 and CD8 CAR-T cells’ developmental trajectories (Fig. 7c), respectively, and their percentage was the most in resting CAR-T cells (Fig. 7c–e). After antigen stimulation, qCD4 TCM could differentiate into cycling CD4 TCM and cycling CD4 TEM, indicating the self-renewal and committed differentiation ability of TCM35 (Fig. 7c), and then cycling CD4 TEM would differentiate into CD4 TE and CD4 TEXH. CD8 CAR-T cells’ developmental trajectories were similar to CD4 CAR-T cells, both involving the development of TCM to TEM to TE/EXH, with the only difference that there was no cycling CD8 TCM in CD8 CAR-T cells’ developmental trajectories (Fig. 7c).T cell exhaustion and glycolysis scores, AP-1 TFs22 (BATF, BATF3, JUNB, and IRF4), and glycolysis-related enzymes (HK2 and ENO1, etc.) upregulated as CAR-T cells developed into effector/exhausted state (Fig. 7b and Supplementary Fig. 9b), demonstrating effector/exhausted T cells’ metabolic shift to glycolysis35 and the role of AP-1 TFs in driving T cell exhaustion.22 Cell division scores revealed qCD8 TCM and qCD4 TCM were the most quiescent with the lowest score, while cycling CD8 TEM and cycling CD4 TEM were the most active in cell division with the highest score (Supplementary Fig. 9b). The variation tendency of exhaus
留言 (0)