
記住我
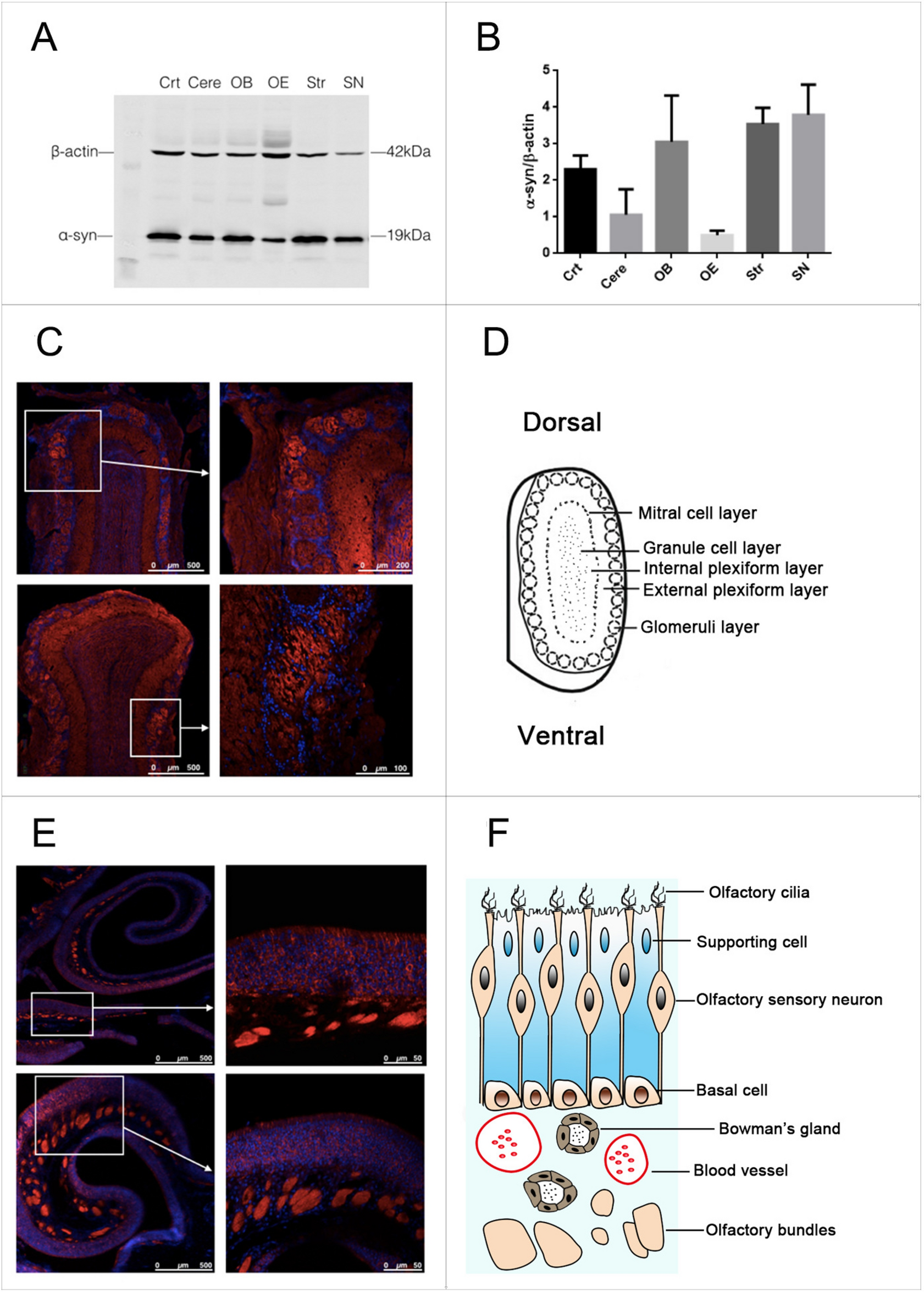


Firstly, RRM2 expression level in HCC was analyzed using the Cancer Genome Atlas (TCGA) and Gene Expression Omnibus (GEO) datasets. The results showed that the expression of RRM2 was upregulated in HCC tissues compared with noncancerous tissues (Fig. 1A–C). However, the methylation level of RRM2 decreased in 377 TCGA-LIHC patients, in contrast to the expression level (Fig. 1D). As illustrated in Fig. 1E, RRM2 overexpression was strongly linked with advanced patient cancer stages, with the highest RRM2 expression observed in stage 3, which corresponds to the degree of vascular invasion. Similarly, RRM2 expression increased as tumor grade increased. The highest RRM2 mRNA expression was seen in grade 4 tumors (Fig. 1F). HBx and RRM2 protein expression was observed by Immunohistochemistry (IHC) staining in 10 HBV-related cirrhosis tissues, with 151 pairs of HCC tumor tissues and adjacent paraneoplastic tissues (85 of which were HBV-related HCC). We discovered that RRM2 expression was significantly higher in HBV-positive HCC tissues compared to HBV-negative HCC tissues, and this tendency was also observed in the noncancerous tissues that correspond to cancerous tissues, which implies that HBV could promote the expression of RRM2 (Fig. 1G). In HBV-related cirrhosis, HBV-related HCC, and its corresponding noncancerous tissues, obvious HBx expression was displayed, and strong positive RRM2 staining was much deeper in HCC tissues than in cirrhosis tissues (Fig. 1H, I). In addition, the expression of RRM2 also depicted an upward trend between cirrhosis and HCC in GSE10143, GSE54236, GSE25097 and GSE17548 datasets from GEO database (Supplementary Fig. S1). Moreover, the RRM2 mRNA level was higher in HBV-related HCC tissues and its corresponding adjacent noncancerous than in normal liver tissues (Fig. 1J-L). These results suggested that RRM2 acted as a possible tumor promoter in HBV-related HCC.
Fig. 1
RRM2 is involved in HBV-related HCC. Boxplot (A) and Pairwise boxplot (B) of RRM2 expression between HCC tumor tissues (T) and pericarcinomatous tissues (P) using heatmaps from TCGA datasets. (C) The expression of RRM2 in HCC tumor tissues was compared with that in the pericarcinomatous tissues in the GSE25097 dataset. (D) RRM2 methylation between HCC tumor tissues and normal tissues in UALCAN dataset. (E) Association between RRM2 mRNA expression and specific cancer stages in HCC patients of UALCAN dataset. (F) Association between RRM2 mRNA expression and tumor grade in HCC patients of UALCAN dataset. (G) Positive area of RRM2 staining in HBV (-) or HBV (+) HCC tumor tissues and pericarcinomatous tissues, respectively and the overally. (H) Representative images of H&E staining, as well as HBx and RRM2 IHC staining in HBV-related liver cirrhosis tissues (n = 10), HBV-related HCC tissues and its corresponding pericarcinomatous tissues (n = 85). Scale bar: 100 μm. (I) Positive areas of RRM2 staining in specified tissues. (J) Relative RRM2 mRNA levels in human normal liver tissues (N), HBV-related HCC tissues (T) and its corresponding pericarcinomatous tissues (P) by qRT-PCR. n = 8 in each group. (K, L) RRM2 protein levels in indicated human liver tissues were determined by WB. ACTB was used as the loading control. n = 8 in each group. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
High RRM2 expression was responsible for a poor prognosis in HCC patientsSince RRM2 was involved in HBV-related HCC, we investigated the clinical significance of RRM2 in patients with HCC. On the RRM2 positive area revealed by immunohistochemical analysis, 148 HCC patients were separated into groups with low and high RRM2 expression. Kaplan–Meier survival analysis demonstrated that higher RRM2 levels were associated with a shorter Overall survival (OS) and Disease free survival (DFS) time than lower RRM2 levels (P = 0.004, P = 0.013, respectively; Fig. 2A, B). Identical outcomes were observed in the HCC group from TCGA dataset (P = 0.005, Fig. 2C). In addition, we conducted a time-dependent ROC curve analysis, which revealed that RRM2 was able to distinguish tumor samples from normal ones and had a satisfactory performance in predicting survival (Fig. 2D). Figure 2E and F depicted the forest plot of univariate and multivariate Cox regression, which indicated that higher RRM2 expression was linked with poorer survival in HCC patients from TCGA dataset as well. As illustrated in Fig. 2G, we created a nomogram to estimate and manage patient efficacy based on the chance of individual survival. The C-indices of the OS, DSS, and progression-free interval (PFI) nomograms were 0.711, 0.79, and 0.633, respectively, showing that the projected results approximated the observed results to a degree. The Calibration plot displayed an approaching straight line, showing a significant correlation between the actual and projected probability at 1.3 and 5 years (Fig. 2H). DCA found that the nomogram model had a better net benefit and a higher probability threshold, indicating that it had clinical usefulness (Fig. 2I).
Fig. 2
Increased RRM2 expression was related with a bad outcome in HCC patients. (A, B) Patients with high-expression levels of RRM2 had shorter OS and DFS times than low RRM2 expression patients in 148 samples. (C) Based on the TCGA dataset, the Kaplan-Meier survival curves depict the OS curves for HCC patients with high and low levels of RRM2 protein expression. (D) Time-dependent ROC curve analysis to assess the predictive efficacy of the prognostic signature. Patients with HCC from TCGA dataset were evaluated using univariate (E) and multivariate (F) Cox proportional hazards analysis to figure out the Hazard ratio (HR) of RRM2 for OS. (G) Nomogram for predicting the probability of 1-, 3-, and 5-year OS, DSS and PFI for HCC patients. (H) Calibration curve for predicting the probability of 1-, 3-, and 5-year OS for HCC patients. (I) DCA plots of the nomogram for predicting the probability of 1-, 3-, and 5-year OS for HCC patients by calculating the C-index
Then, we searched in the cBioPortal database to determine the types and frequencies of RRM2 mutations. Genes altered in 11.85% of 363 HCC patients, with mRNA upregulation occurring in 34 cases (9.37%), amplification occurring in 6 cases (1.65%), and multiple alterations occurring in 3 cases (0.83%; Fig. S2A). Thus, the observed genetic variation of RRM2 in HCC was primarily due to amplification. In comparison to the diploid group, the amplification group exhibited significantly higher levels of RRM2 expression (P < 0.001; Fig. S2B, C). Additionally, the Kaplan-Meier plot showed an association between RRM2 CNV and a shorter OS (log-rank test, P = 0.0156), a shorter DSS (log-rank test P = 0.0284), and a shorter progression free survival (PFS, log-rank test P = 6.241E-3) in HCC patients (Fig. S2D-F). These findings suggested that RRM2 and its genetic variants may also have a noticeable impact on HCC patients’ prognosis.
Analysis of RRM2 biological significance and potential molecular mechanism in HCCBased on the TCGA dataset, a total of 697 DEGs were identified as statistically different between HCC RRM2-high samples and low controls. Figure 3A depicted the relative expression values of each group’s top ten DEGs. As seen in Fig. 3B, the expression of RRM2 was positively associated with that of CCNB1 (R = 0.877), CDCA5 (R = 0.884), CCNB2 (R = 0.873), KIF4A (R = 0.909), KIFC1 (R = 0.877), NCAPG (R = 0.914), SPC25 (R = 0.0.897), TOP2A (R = 0.884), TPX2 (R = 0.903), ZWINT (R = 0.879) genes (all P < 0.001). The result of GSEA analysis exhibited that high expression of RRM2 was clearly involved in a multitude of carcinogenic signal, such as cell cycle, PLK1 pathways, FOXM1 pathways, and P53 pathways (Fig. 3C).
Fig. 3
Analysis of RRM2 biological significance and potential molecular mechanism in HCC. (A) Heat map of top 10 DEGs in RRM2-high expression samples and low samples groups. (B) Expression correlation between RRM2 and representative genes (CCNB1, CDCA5, CCNB2, KIF4A, KIFC1, NCAPG, SPC25, TOP2A, TPX2, ZWINT) of the top RRM2 positive correlated genes in TCGA projects as determined. (C) GSEA of RRM2 in HCC. The 8 significant pathways of RRM2 GSEA results in HCC
HBx promoted HCC cell tumorigenesis via autophagyTo confirm the oncogenic activity of HBx and its effects on HCC tumorigenicity, orthotopic xenograft tumor models were firstly developed in nude mice using Huh-7 liver cancer cells lines transfected with either LV-HBx or LV-NC. Six weeks following injection, all null mice developed orthotopic xenograft tumors at the injection site, which were collected. We found that HBx played a significant role in encouraging tumor growth, and the “primary tumors” were shown in Fig. 4A. The relative liver weights were significantly higher in the HBx-OE group than in the NC group (Fig. 4B). Moreover, the number of tumors was much lower in the NC group than in the HBx-OE group (Fig. 4C). Histopathological examination of tumor tissues indicated that they were hepatocellular carcinogenesis (Fig. 4D). WB analysis revealed that increased HBx expression was associated with decreased Bcl2 levels and increased Bax expression, indicating that apoptosis inhibition may be a potential cause of liver tissue carcinogenesis (Fig. 4F, G).
Fig. 4
HBx promoted HCC cell tumorigenesis and autophagy. (A) Images of the tumors of nude mice from the NC or HBx-OE group. (B) The average liver weight as a percentage of total body weight. (C) The average tumor number of liver tissues as quantified. (D) Examples of liver tissues from the NC group or HBx-OE group stained with H&E. Scale bars: 100–200 μm. (E) Representative images of SQSTM1 staining in liver tissues from two groups. Scale bars: 100–200 μm. (F, G) HBx, SQSTM1, BCL2 and Bax protein expression in liver tissues from two groups were determined by WB. Quantification of proteins was shown in the right panel. **P < 0.01, *** P < 0.001, ****P < 0.0001
Autophagy is a crucial process that breaks down cellular components to maintain balance in HCC. It serves various purposes, but the exact way it responds to the tumor-inducing effects of HBx is not yet understood [29]. So, we firstly proved that HBx could promot autophagy in HCC. Following that, we investigated the role of autophagy in HBx-induced growth of tumors by examining the effects of autophagy on the proliferation and apoptosis of LO2 and Huh-7 cells transfected with LV-HBx or LV-NC. WB analysis showed the relative SQSTM1 protein expression level in the HBx-OE group was reduced compared with that of the NC group (Fig. 4F, G). Consistently, IHC analysis further showed the same results as depicted in Fig. 4E. Electron microscopy showed that the number of typical autophagosomes with double membranes was significantly increased in HBx-expressing LO2 (LO2-HBx) and HBx-expressing Huh-7 cells (Huh-7-HBx) (Fig. 5A, B). additionally, the HBx overexpression increased the conversion of LC3-I into LC3-II and decreased SQSTM1 protein, an autophagic flux marker, in LO2 and Huh-7 cells (Fig. 5C, D). Using a EdU incorporation assay, we analyzed the impact of autophagy on the proliferation of Huh-7-HBx cells and LO2-HBx cells. As demonstrated in Fig. 5E-G, HBx overexpression stimulated the proliferation of LO2 and Huh-7 cells. Whereas, treatment with the autophagy inhibitor 3-MA inhibited the HBx-induced proliferation. Multiple investigations have demonstrated that autophagy may precede apoptosis [23]. To determine whether HBx-related autophagy was associated with apoptosis in HCC cells, we measured the level of apoptosis in LO2-HBx cells and Huh-7-HBx cells using flow cytometry with Annexin V and PI staining after autophagy inhibition. As depicted in Fig. 5H-J, the HBx overexpression group experienced a significantly lower apoptosis rate than the control group. However, when HBx-overexpressed cells were treated with 3-MA to inhibit autophagy, the rate of apoptosis rose. Taken together, these findings indicated that HBx induced autophagy, which promoted the formation liver cancer.
Fig. 5
HBx promoted HCC tumorigenesis via autophagy. (A, B) Autophagosomes and autolysosomes were observed in LO2 and Huh-7 cells transfected with HA-HBx plasmid or empty NC. Scale bar, 200 nm. (C) The expression of the autophagy-associated proteins, LC3B, and SQSTM1 in LO2 and Huh-7 cells were detected using the corresponding antibodies with ACTB as the control by WB. (D) Relative expression levels of the indicated proteins in different groups (n = 3). LO2 or Huh-7 cells transfected with the HA-HBx construct or empty NC were treated with/without 3-MA (5 mM) for 24 h. (E) After treatment, the EdU incorporation assay was used to examine cell proliferation. Scale bar: 200 μm. (F, G) Quantification of positive cells in the EdU assay. (H-J) Using Annexin V-FITC/PI assay, the apoptotic index of cells subjected to diverse treatments was determined. Flow cytometry was used to examine the results (n = 3). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
HBx stimulated RRM2 expression in HCCTo investigate further the potential role of HBV on RRM2, the expression of RRM2 was analyzed in LO2, HepG2, and Hep2.2.15 cells. The results showed that RRM2 was significantly elevated both at the mRNA level and the protein level in HepG2 cells compared with LO2 cells, suggesting that RRM2 is most likely an oncogene that contributes to hepatocarcinogenesis. In the meantime, we compared the expression in HepG2 of hepatocellular carcinoma cells from the same source but with varying infection status. Compared to HepG2, the mRNA level or protein expression level of RRM2 was found to be even higher in HepG2.2.215, suggesting that HBV infection promotes RRM2 expression to some degree (Fig. 6A, B). Immunofluorescent double labeling of HBx and RRM2 showed that both proteins were expressed in numerous common locations in human HCC tissues (Fig. 6C). Moreover, we discovered that RRM2 mRNA and protein expression was markedly increased following transfection of LO2 and Huh-7 cells with the HBx plasmid (Fig. 6D, E). Meanwhile, the expression of RRM2 in the liver tissues of HBx-OE mice group were elevated to a greater extent than that of control mice group (Fig. 6G, F, I). Then, the interaction between HBx and RRM2 was analyzed in LO2-HBx cells and Huh-7-HBx cells by co-immunoprecipitation, and the results revealed that RRM2 was coupled with HBx (Fig. 6H).
Fig. 6
HBx stimulated RRM2 expression in HCC. (A) RRM2 mRNA expression levels in LO2, HepG2, and HepG2.2.15 cells (n = 3). (B) Analysis of RRM2 protein levels in the indicated cells using WB analysis (n = 3). (C) HCC tumor tissues underwent double immunofluorescence staining for HBx and RRM2. The nuclei were dyed blue with DAPI. Scale bar: 200 μm. (n = 6). (D) The protein expression levels of RRM2 in LO2 and Huh-7 cells transfected with either LV-HBx or LV-NC plasmids were detected using the corresponding antibodies with ACTB as the control by WB. (E) Quantification of the relative RRM2 protein expression in cells (n = 3). (F, G) HBx and RRM2 protein expression in liver tissues of nude mice from the NC and HBx groups were determined by WB (n = 6). (H) Anti-HBx antibody was used to perform immunoprecipitation on the cell lysates. Anti-RRM2 antibody was used to identify endogenous RRM2 that was co-immunoprecipitated. (I) Representative images of RRM2 staining in liver tissues from the NC and HBx groups (n = 6). *P < 0.05, **P < 0.01
Blockade of RRM2 strongly suppressed HBx-induced autophagosome formationOur preceding data demonstrated that HBx promotes the expression of RRM2 and induces autophagy in HCC. However, the link between autophagy and RRM2 remains obscure. To further investigate the influence of RRM2 on the HBx-induced autophagic response, siRNA targeting RRM2 (si-RRM2) was transfected into LO2-HBx cells and Huh-7-HBx cells. The efficacy of RRM2 knockdown was detected 48 h after transfection (Fig. 7A). The transient transfection of si-RRM2 inhibited the HBx-induced increase in LC3B expression (Fig. 7B). In order to further investigate the impact of RRM2 on the autophagic response triggered by HBx, we utilized a stubRFP-sensGFP-LC3 reporter system for observing alterations in autophagic flux. Autophagosomes exhibited both GFP and RFP simultaneously, however LC3 in autolysosomes only showed red fluorescence due to the acidic environment in the lysosome lumen, which affected the GFP signal. The merged picture of Huh-7 cells infected with HBx lentivirus showed a clear and noticeable increase in red and yellow puncta. Nevertheless, the use of small interfering RNA (si-RRM2) to knock down RRM2 resulted in the disappearance of fluorescent puncta, particularly those that were yellow. This indicated that RRM2 knockdown hindered the flow of autophagy and primarily hindered the creation of autophagosomes (Fig. 7C, D). Similarly, LO2-HBx-siRRM2 cells and Huh-7-HBx-siRRM2 cells contained less GFP-LC3B puncta formation and lipidation than LO2-HBx cells and Huh-7-HBx cells, respectively (Fig. 7E, F). Ultrastructural analysis demonstrated typical autophagosomes in LO2-HBx cells and Huh-7-HBx cells, but autophagosomes were scarce after the transient transfection of si-RRM2 (Fig. 7G, H). All these results indicated that RRM2 blockade impaired the autophagy process induced by HBx.
Fig. 7
HBx-induced autophagosome production was inhibited by RRM2 blockade. LO2 or Huh-7 cells transfected with either LV-HBx or LV-NC plasmids were treated with/without siRRM2 for 48 h. (A) After treatment, RNA samples were then collected and relative gene expression ratios of RRM2 in cells were determined using RT-PCR analysis (n = 3). (B) Cell lysates were collected and analyzed by immunoblotting using anti-LC3B antibody. Quantification of the relative LC3B protein expression in cells (n = 3). (C) Huh-7 cells that had been genetically modified to express HBx were exposed to stubRFP-sensGFP-LC3 lentivirus, and then treated with si-RRM2 or si-NC for a period of 48 h. The nuclei are labeled with DAPI staining. Scale bar: 20 μm. (D) Measurement of the number of LC3 puncta in each field. (E) LC3 immunofluorescent staining in cells. Scale bar: 200 μm. (F) The mean number of LC3 puncta per cell was computed (n = 50 cells). (G, H) With transmission electron microscopy, double membrane autophagosome structures were identified in cells, as indicated by the arrows. Scale bar: 200 nm. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
HBx promoted HCC cell tumorigenesis via RRM2In many varieties of cancer, autophagy is inextricably linked to the cell cycle, whereas abnormal cell proliferation results from cell cycle changes. Since RRM2 is an intermediate mediator of HBx-induced autophagy, does RRM2 partake in HBx-regulated cell cycle regulation by affecting the cell cycle? For this reason, we utilized flow cytometry to examine the alterations in the cell cycle following RRM2 expression interference. Our data demonstrated that HBx-expressing cells rapidly exited the G0/G1 phase, entered the S phase, and nearly tripled their proliferation rate compared to HBx-deficient cells, whereas the si-RRM2 treatment inhibited the effect of HBx on proliferation (Fig. 8A-C). The results of EdU staining confirmed this conclusion. EdU experiments demonstrated that HBx overexpression boosted cell proliferation, but this effect was reversed after RRM2 interference (Fig. 8D-F). In addition, RRM2 knockdown downregulated PCNA, a factor related to DNA replication, indicating that HBx-induced stimulation of hepatocyte proliferation was dependent on RRM2 (Fig. 8G, H). Meanwhile, to determine whether RRM2 was required for HBx-induced inhibition of hepatocyte apoptosis, we examined the level of apoptosis in LO2-HBx and Huh-7-HBx cells after RRM2 knockdown using flow cytometry with Annexin V and PI staining. As depicted in Fig. 9A-C, more significant apoptosis was observed in LO2-HBx and Huh-7-HBx cells treated with si-RRM2. As depicted in Fig. 9D and E, overexpression of HBx enhanced the protein expression level of BCL2 and decreased the protein expression level of Bax, whereas treatment with si-RRM2 attenuated this effect. The aforementioned findings suggest that HBx promotes hepatocarcinogenesis via RRM2, with the mechanism involving the regulatory role of RRM2 in cell proliferation and apoptosis.
Fig. 8
HBx promoted HCC cell proliferation via RRM2. LO2 or Huh-7 cells transfected with either LV-HBx or LV-NC plasmids were treated with/without siRRM2 for 48 h. (A-C) Analysis of cell cycle distribution among different groups by flow cytometry. (D) EdU incorporation assay was used to assess cellular proliferation following treatment. Scale bar: 200 μm. (E, F) Quantitative analysis of EdU-positive cells. (G) WB analysis of PCNA protein expression in LO2 or Huh-7 cells with the above treatments. (H) Quantitative of the relative PCNA protein expression in cells (n = 3). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
Fig. 9
HBx promoted HCC cell apoptosis via RRM2. LO2 or Huh-7 cells transfected with the HA-HBx construct or empty NC were treated with/without siRRM2 for 48 h. (A) Analysis of apoptotic cells by flow cytometry. Representative Annexin V-FITC/PI-stained flow cytometry plots for apoptosis. (B, C) The proportion of apoptotic cells was displayed on the right panel. (D) WB analysis of BCL2 and Bax protein expression in LO2 or Huh-7 cells with the above treatments. (E) Quantitative of the relative BCL2 and Bax protein expression in cells (n = 3). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
留言 (0)