Human subjects and ethics approval
Infertility patients diagnosed with the oocyte death phenotype were recruited from the Center of Reproductive Medicine, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology. Participants with normal oocytes and embryos in IVF/ICSI cycles were also recruited as the control group. All oocytes and embryos from controls and patients were obtained with written informed consent signed by the donor couples.
This study was approved by the ethics committee on human subject research at Tongji Hospital, Huazhong University of Science and Technology (TJ-IRB20220450). The animal experiments were approved by the Animal Welfare and Ethics Committee of Tongji Hospital (TJH-202,210,011).
Wholeexome sequencing (WES) and Sanger sequencing
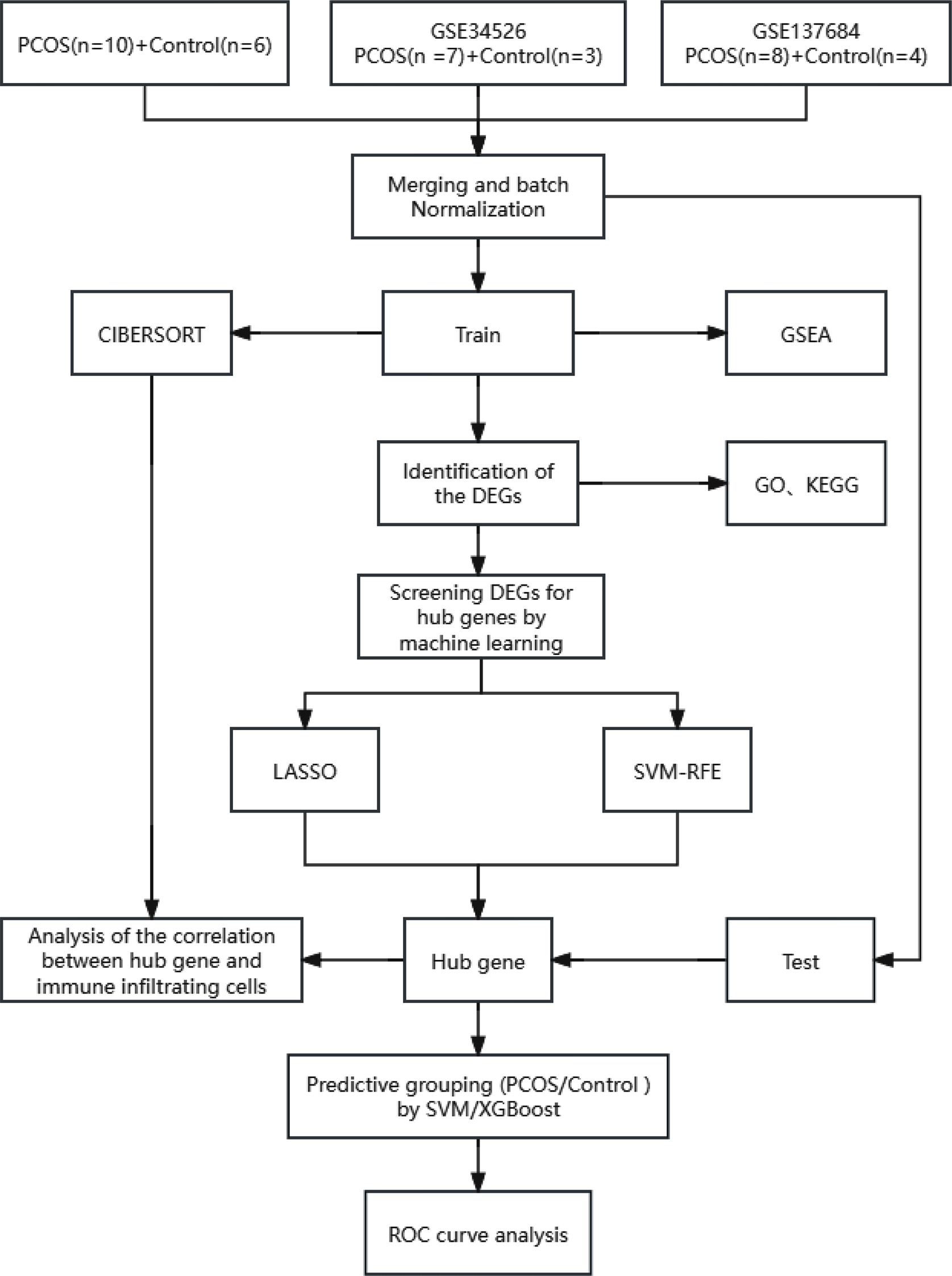
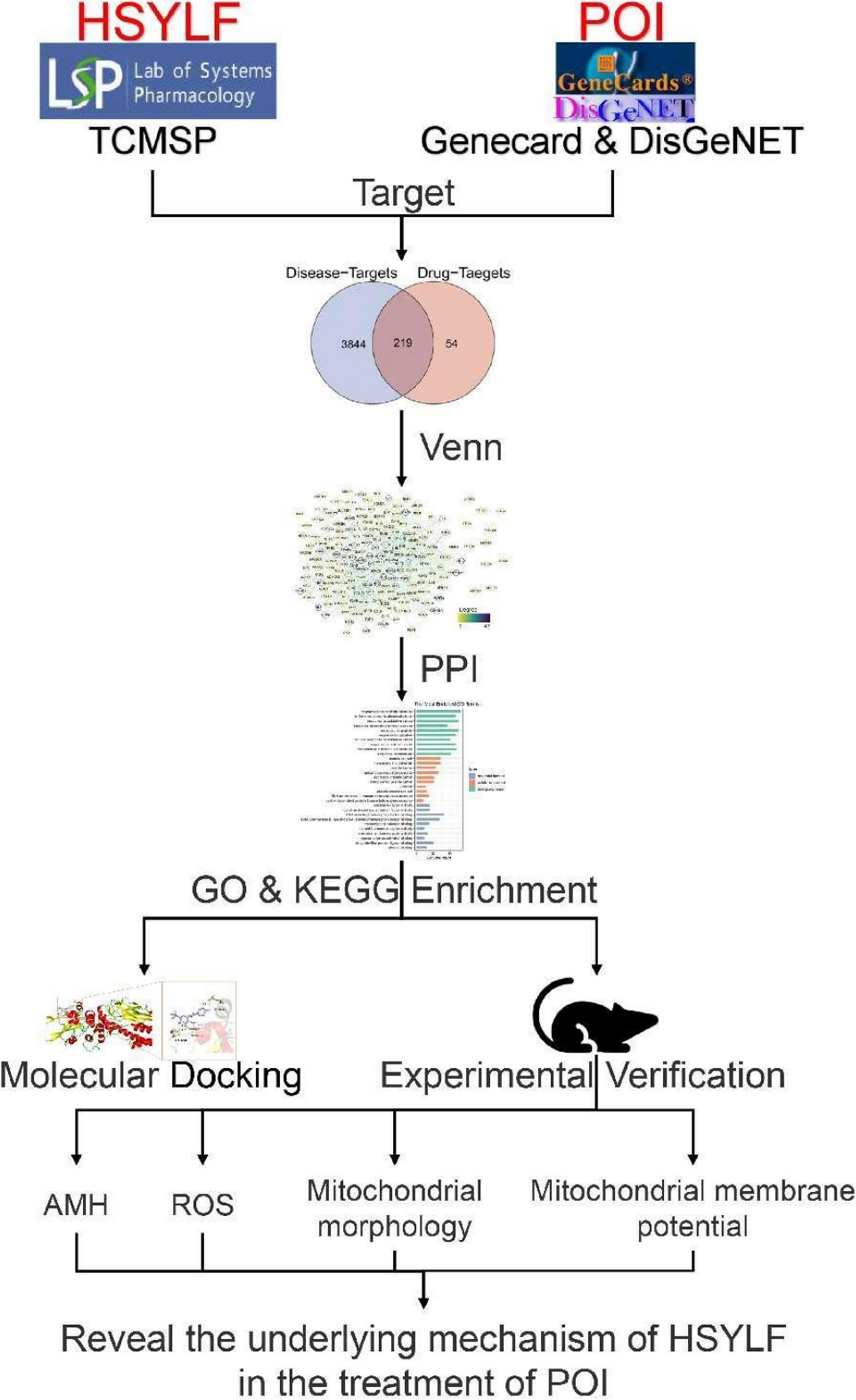
Genomic DNA was extracted from peripheral blood samples of the patients and their members for WES to identify potential disease-causing variants according to the manufacturer’s instructions. The details of the genetic analysis procedure have been well described previously [10, 22]. Candidate variants identified in the participants were validated by Sanger sequencing analyses conducted on ABI PRISM 3500 Genetic Analyzer (Applied Biosystems, Foster City, CA).
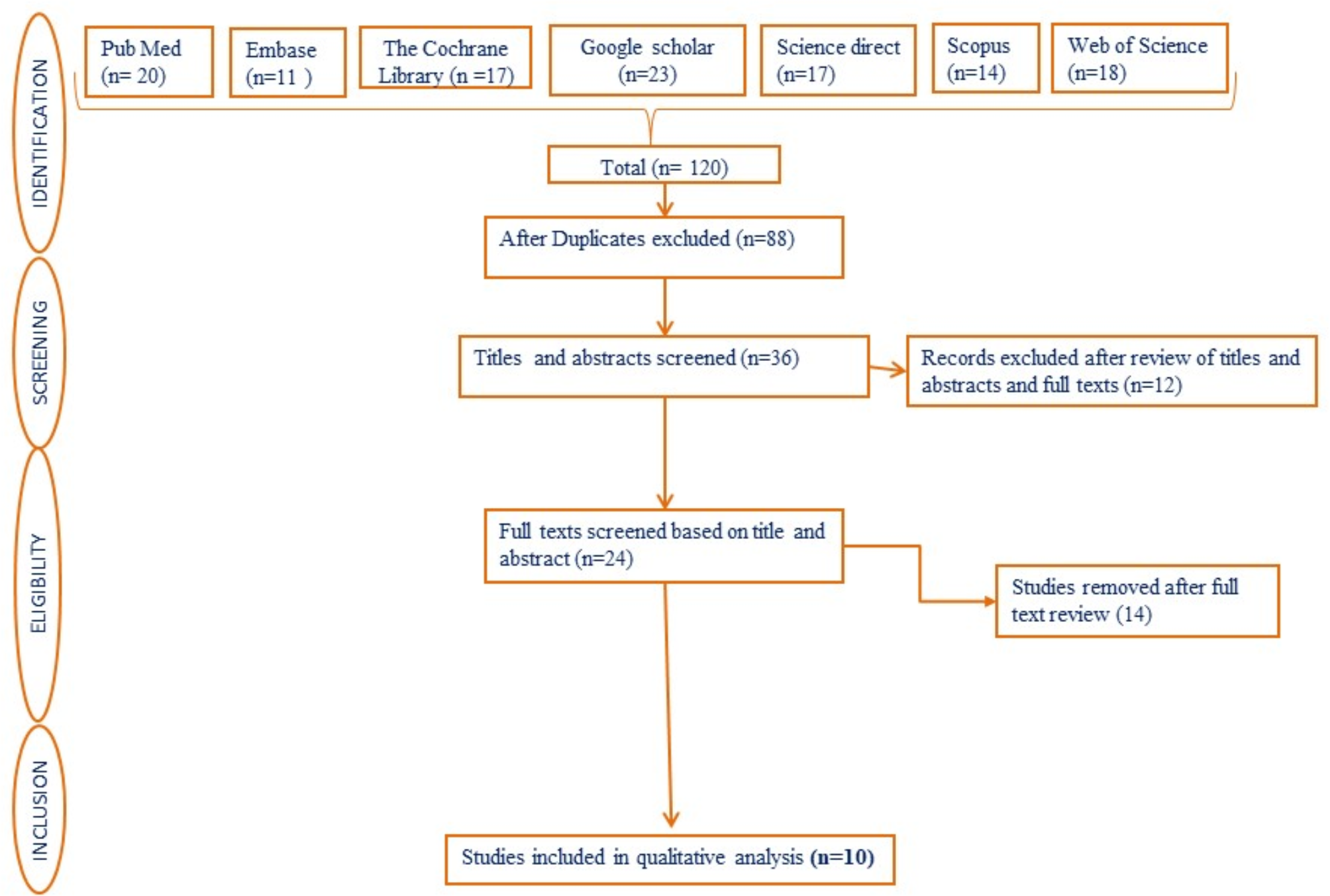
The specific filtering process of variant sites was as follows: (a) The variant sites in the 1000 Genomes database and Exome Aggregation Consortium (frequency > 0.01 in the population) were filtered, and the inter-individual diversity sites were removed to obtain rare variants that may actually cause disease; (b) The variant sites of exonic region or splicing site region (10 bp upstream/downstream) were retained; (c) Synonymous variants (without changing amino acids) were removed to obtain variants that affect gene expression products; (d) The variant sites predicted to have an impact on protein structure or function in more than 2 online variants prediction tools were retained; (e) Screen variations in genes associated with female infertility. At last, we could lock the variants consistent with clinical phenotype and genetic pattern. The details on variant selection were showed in the supplementary table.
Variants analysis and molecular modelling
Evolutionary conservation was assessed using Clustal Omega software (https://www.ebi.ac.uk/Tools/msa/clustalo/). The allele frequency of the variants in the general population was assessed using Genome Aggregation Database (GnomAD, http://gnomad.broadinstitute.org/). The pathogenicity of the variant was assessed using three online software: sorting intolerant from tolerant (SIFT, sift.jcvi.org), polymorphism phenotyping (PloyPhen2, genetics.bwh.harvard.edu/pph2), and mutation taster (http://www.mutaiontaster.org/). The structure model was built based on Cryo-EM structure of wild-type human pannexin1 channel (PDB ID, 6WBF/A) in the RCSB Protein Data Bank (https://www.rcsb.org/). PyMOL software (https://pymol.org/2/) was used to analyze the effect of the variants in PANX1 protein.
Expression vector construction
Wild-type (WT) human PANX1 and mutated PANX1 (p.Asn326del, p.Ser137Leu) were constructed and then recombined with the eukaryotic expression vector pcDNA3.1. A 3×FLAGtag was fused at the C-terminus of WT and mutated PANX1, respectively. The plasmids were constructed by OBiO Technology (Shanghai).
Cell culture and transfection
HeLa cells were obtained from Cancer Biology Research Center of Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology. Cells were cultured in Dulbecco’s Modifed Eagle Medium/Nutrient Mixture F-12 (DMEM/F-12, KeyGEN Bio TECH, Jiangsu, China) supplemented with 1% penicillin/ streptomycin (Servicebio, Wuhan, China) and 10% (v/v) fetal bovine serum (FBS, Wisten, Nanjing, China) in a humidified incubator with a 5% CO2 atmosphere at 37 °C. PANX1 WT and mutant constructs were transfected into HeLa cells using liposomal transfection reagent (Yeasen, Shanghai, China) according to the manufacturer’s instructions.
Western blotting
HeLa cells were harvested 36 h after transfection and washed three times with cold phosphate-buffered saline (PBS, Servicebio, Wuhan, China). Cells were lysed in RIPA lysis buffer with 2% of protease inhibitor (Servicebio, Wuhan, China). After incubating on the ice for 20 min and centrifuging at 12,000 rpm at 4 °C for 20 min, protein lysates were collected in a new centrifuge tube. Protein concentrations were determined with a BCA protein quantitative detection kit (Servicebio, Wuhan, China). Then, cell extracts were mixed with 5×sodium dodecyl sulfate (SDS) loading buffer (Servicebio, Wuhan, China) and denatured by boiling at 100 ℃ for 10 min. Equal amounts of protein were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose filter membranes (Merck KGaA, Darmstadt, Germany). The membranes were blocked in 5% skim milk diluted in 1× Tris-buffered saline (Servicebio, Wuhan, China) with 0.1% Tween 20 (TBST, Servicebio, Wuhan, China) for 1 h and incubated at 4 °C overnight with rabbit anti-PANX1 (1:1000 dilution, Cell Signaling Technology, kind gifts from Lei Wang’s lab in Fudan University) or rabbit anti-vinculin (1:1000 dilution, ABclonal, Wuhan, China) antibodies. After incubation with the goat anti-rabbit IgG secondary antibodies (1:2000 dilution, Servicebio, Wuhan, China) for 1 h at room temperature on the secondary day, the membranes were detected by ECL chemiluminescence kit (Vazyme, Nanjing, China) and imaged on a chemiluminescent imaging system (GeneGnome XRQ, Syngene, England). For densitometric analyses, protein bands on the blots were measured by ImageJ software.
Complementary RNAs (cRNAs) transcription
WT and mutant PANX1 cRNAs (RNA transcripted from complementary DNA) was constructed by GenScript (Nanjing, China). Briefly, WT and mutant constructs were linearized by digestion with the AgeI restriction enzyme, then purified linearized DNA was used as a template to transcribe PANX1 cRNAs.
Collection and microinjection of mouse GV oocytes and fertilized oocytes
For GV oocytes collection, ovaries were isolated from 6 to 8-week-old Kun-Ming (KM) female mice (Biont, Hubei, China). Germinal vesicle (GV) oocytes were harvested by puncturing the large ovarian follicles and collected by mouth pipetting on the stage of a dissecting microscope. The GV oocytes were cultured in M2 medium (Aibei Biotechnology, Nanjing, China) under mineral oil (Servicebio, Wuhan, China) in an atmosphere of 5% CO2 at 37 °C.
For fertilized oocytes collection, the 6 to 8-week-old KM female mice were injected with 7.5 IU pregnant mare serum gonadotropin (PMSG, Aibei Biotechnology, Nanjing, China). 44–48 h after PMSG injection, each mouse was injected with 7.5 IU human chorionic gonadotropin (hCG, Aibei Biotechnology, Nanjing, China), and the mice were then euthanized after 14 h. Oviducts were removed and transferred to a pre-warmed human tubal fluid (HTF, Aibei Biotechnology, Nanjing, China) medium. Then cumulus oophorus masses (COMs) containing oocytes were removed by dissecting oviducts and transferred to the HTF medium. For sperm collection, 10 to 12-week-old male mice were euthanized, and the epididymal tails were taken out and transferred to a pre-warmed HTF medium. Scratches were made on the tails of the epididymis by an insulin needle to allow sperms to escape. Then they were incubated for 1 h to complete the capacitation. Finally, COMs were mixed with sperms in HTF for fertilization. Two pronuclei (2PN) zygotes were collected 6 h after fertilization.
The collected mouse GV oocytes and fertilized oocytes were microinjected with WT or mutant PANX1 cRNAs using a Narishige micromanipulator. About 5 to 10 pl of cRNA solution (250 to 1000 ng/µl) was microinjected into the cytoplasm of each mouse oocyte. Injected GV oocytes were matured in vitro in M2 medium (Sigma-Aldrich, USA) for 12 h, and the morphological changes of oocytes were recorded. Injected zygotes were cultured in KSOM medium (Aibei Biotechnology, Nanjing, China) for 12 h, and the morphological changes and embryo development were recorded. All oocytes and zygotes were cultured at 37 °C in an atmosphere of 5% CO2. For PANX1 channel inhibition experiments, carbenoxolone (CBX, MedChemExpress, Shanghai, China) was used at a concentration of 300 µM.
Mouse oocyte ATP measurements
Mouse GV oocytes were cultured in 80 µl of M2 medium containing 300 µM ARL 67,156 trisodium (Sigma-Aldrich, USA) with or without 300 µM CBX, and collected at 6 h after injection with WT or mutant PANX1 cRNAs. The relative ATP content was determined with the Enhanced ATP Assay Kit (Beyotime, Shanghai, China). For extracellular ATP measurements, a mixture of 50 µl of culture medium and 50 µl of ATP assay reagent was assayed using a luminometer (Servicebio, Wuhan, China). For intracellular ATP measurement, 20 GV oocytes were lysed with 100 µl of ATP assay lysis buffer for 10 min and centrifuged at 4000 g at room temperature for 30 s. A mixture of 50 µl of supernatant and 50 µl of ATP assay reagent was assayed using the luminometer. The relative ATP concentration was expressed as a ratio of all values with respect to the WT group.
Mouse oocyte cell membrane potentials measurements
Mouse GV oocytes were cultured in M2 medium with or without 300 µM CBX, and collected at 6 h after injection with WT or mutant PANX1 cRNAs. Oocytes were first incubated with Hoechst33342 (Servicebio, Wuhan, China) for 5 min and washed three times with M2 medium. Oocytes were then incubated with M2 medium containing 5 µM cell membrane potential sensitive fluorescent dye DiBAC4 [3] (MedChemExpress, Shanghai, China) and immediately examined and photographed under confocal laser scanning microscope (Zeiss LSM 510 META, Germany). The relative fluorescence intensity was measured by ImageJ and expressed as a ratio of all values with respect to the WT group.
Microinjection of human oocytes
Human GV oocytes were collected from ICSI patients on the day of oocyte retrieval after denudation. And 3PN fertilized oocytes were collected from IVF patients one day later after fertilization ascertainment. All patients in our center donating oocytes for research were informed and signed written consent. The microinjection procedure was similar to that of mice. Briefly, about 8 to 10 pl of cRNA solution (1000 ng/µl) was microinjected into the cytoplasm of each human GV oocyte and pronucleus oocyte. Then the injected oocytes were cultured in a time-lapse incubator with 6% CO2 and 5% O2 at 37 ℃. The morphological changes of oocytes and subsequent embryonic development were recorded accordingly.
Immunofluorescence staining
The oocytes and embryos were collected and fixed with 4% (w/v) paraformaldehyde in PBS for 1 h at room temperature, and then were permeabilized in PBS containing 0.1% (w/v) Triton X-100 and 1% BSA for 30 min at room temperature. After washing three times with PBS containing 1% BSA, oocytes and embryos were incubated in blocking solution (PBS containing 3% BSA) for 1 h at room temperature. Then oocytes and embryos were incubated at 4 ℃ overnight with rabbit anti-PANX1 (1:100 dilution, Sigma-Aldrich, kind gifts from Lei Wang’s lab in Fudan University) antibody. On the second day, oocytes and embryos were incubated with FITC-labeled secondary antibody (1:100 dilution, Servicebio, Wuhan, China) for 1 h. After washing three times, oocytes and embryos were incubated with Tubulin-Tracker Red (Beyotime, Shanghai, China) for 30 min at room temperature. Then oocytes and embryos were incubated with 4′,6-diamidino-2-phenylindole (DAPI, Servicebio, Wuhan, China) for 20 min at room temperature. Finally, the samples were mounted on glass slides and examined with a confocal laser scanning microscope (Zeiss LSM 510 META, Germany).
Statistics
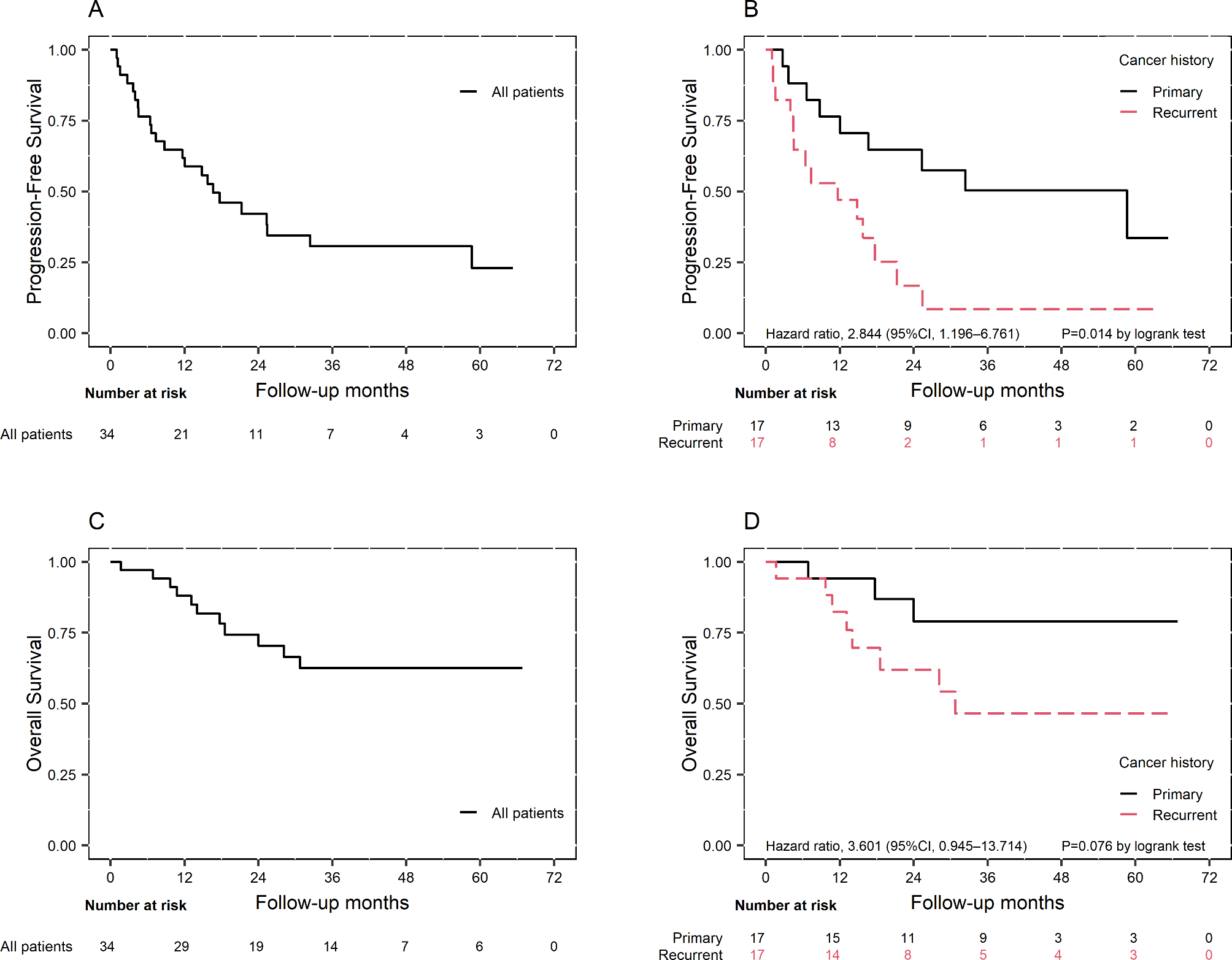
All data are representative of three independent experiments. GraphPad Prism (version 9.0) was used to perform the statistical analysis. Values were analyzed by Student’s t-tests when comparing experimental groups. Wald p-values were two-sided; p < 0.05 was considered to be statistically significant.




留言 (0)