
記住我



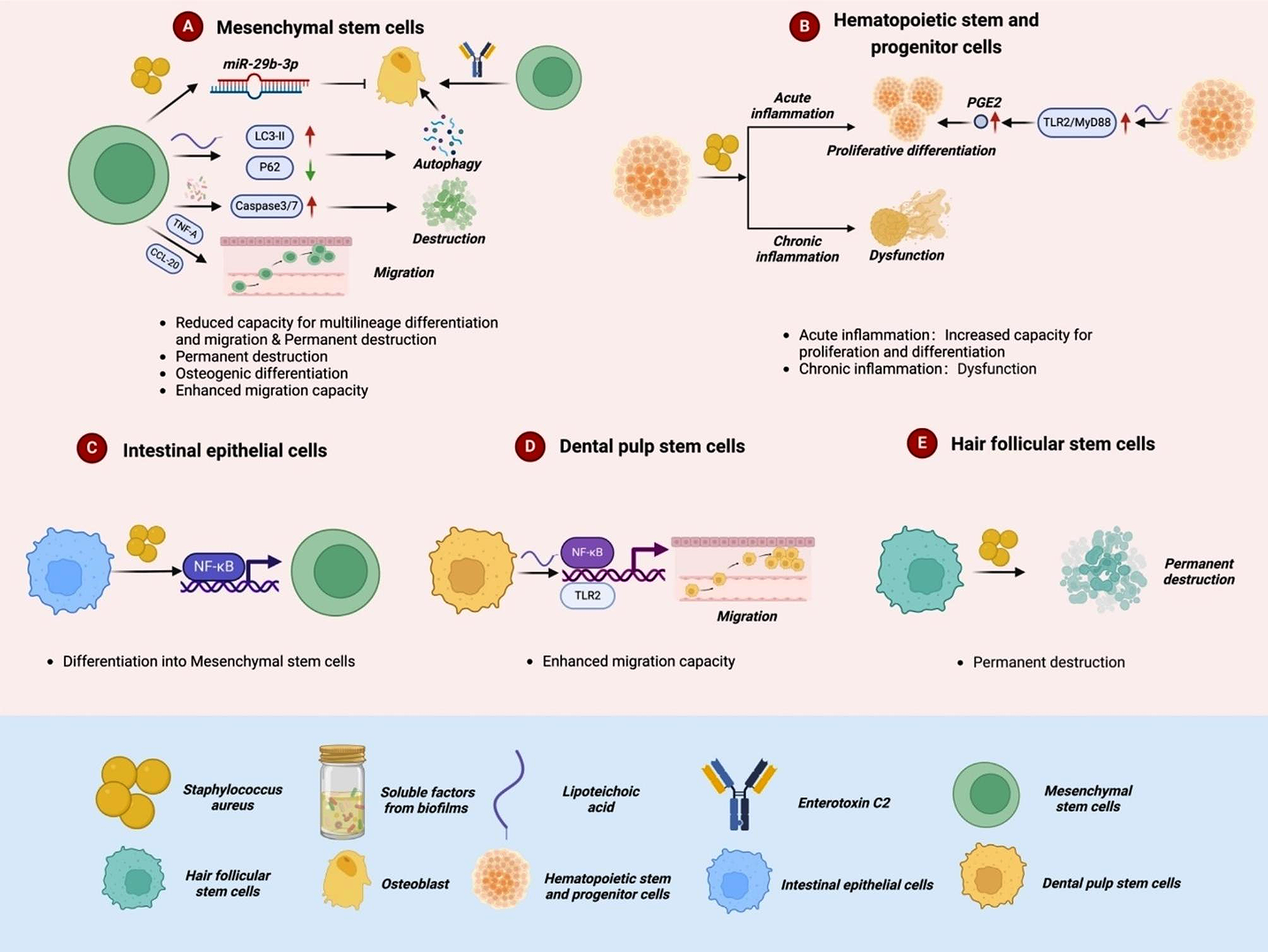

Based on our previous work demonstrating the efficient production of iPSC-derived macrophages [20, 23, 36], we here introduce a standardized, xeno-free, chemically defined and easy-to-use intermediate-scale benchtop bioreactor platform dedicated to producing iPSC-derived macrophages via 3D hematopoietic organoids, which we refer to as hemanoids.
To generate hemanoids, we initiated the differentiation process in a benchtop bioreactor by introducing a single-cell solution of hiPSCs along with mesoderm priming cytokines (SCF, BMP4, VEGF) in defined E8 medium for 24 h to allow for aggregate formation. Subsequently, the medium was changed to mesoderm priming medium 1b, and the aggregates were cultivated for an additional 3 days. On days 4 and 7, we performed another medium change for the developing hemanoids and added IL-3 along with the mesoderm priming cytokines. After 10 days of mesoderm priming, the medium was changed to hematopoietic differentiation medium containing only IL-3 and M-CSF, and the culture of the hemanoids was continued for several weeks for macrophage production (Fig. 1A).
Fig. 1
Organoid-based production of iPSC-Mac in intermediate-scale bioreactors recapitulates embryonic hematopoietic development. A Schematic representation of the manufacturing process in an intermediate-scale benchtop bioreactor to produce hemanoids and the continuous generation of iPSC-derived macrophages. B Immunohistochemical analysis of VE-Cadherin/CD144, VEGFR and CD45 expression in hemanoids derived from day 4, 7 and 10 of mesoderm priming as well as from hemanoids during hematopoietic differentiation after they initiated production of iPSC-derived Mac (day 7–10). Arrows indicate characteristic regions (scale bar = 200 µm, data shown for iPSC line#1, representative of n = 2). C Flow cytometric analysis of CD34, CD144, CD43 and CD45 expression to identify different hemato-endothelial progenitor populations. Hemanoids were dissociated and analyzed on day 4, 7 and 10 of mesoderm priming as well as during hematopoietic differentiation after they initiated production of hiPSC-derived Mac. Populations were pre-gated for single cells, CD34+ cells and viable cells. Subsequently, the frequency of CD144+/CD45− Hemato-endothelial progenitors, CD144-/CD43+ early hematopoietic progenitors and CD144-/CD45+ hematopoietic progenitors was analyzed in the CD34+ population (individual values with mean ± SD, iPSC line#1: dark blue dots, n = 2 and iPSC line#2: blue squares, n = 3. Gating strategy and representative plots can be found in Supplementary Fig. S1). D Gene expression analysis of key genes for pluripotency (POU5F1 (OCT4)), hemato-endothelial progenitors (SOX17), and hematopoietic progenitors (RUNX1) at different stages of differentiation as well as in hiPSC-derived macrophages (iPSC-Mac) by qRT-PCR. Values are represented as relative RNA expression to GAPDH (housekeeping gene) (individual values with mean ± SD, iPSC line#1: dark blue dots, and iPSC line#2: blue squares, n = 2–3 per line, n.d. indicates detection limit of the target gene)
To elucidate the developmental processes and hematopoietic commitment in the hemanoids during the mesoderm induction in a benchtop bioreactor, we analyzed the hemanoids derived from two iPSC lines: (our in-house, lenti-viral reprogrammed hiPSC line (hCD34iPSC11, iPSC line#1) [37] and an episomal reprogrammed, potential GMP-compatible hiPSC line (LiPSC-GR1.1, iPSC line#2) [38] at days 4, 7, 10 of mesoderm priming and after one week of hematopoietic differentiation, when the hemanoids began to produce iPSC-derived macrophages. In more detail, we dissected the cellular composition of our hemanoids and performed immunohistochemistry, immunophenotyping, and gene expression to determine the emergence of crucial markers and cell populations during the successive development of the hemanoids.
The early mesoderm marker VEGFR2/KDR displayed prominent expression at day 4 with gradual decrease at day 7 and pronounced reduction at day 10 and during hematopoietic differentiation as shown by immunohistochemistry and RT-qPCR (Fig. 1B, Fig. S1A). Additionally, we observed VE-Cadherin/CD144 expression (indicating the development of (hemato-endothelial cells) at low levels already at day 4, a clear increase at day 7 and gradually decrease at day 10 and during hematopoietic differentiation. Expression of CD45, demonstrating the emergence of hematopoietic cells was observed for the first time at day 10 and during hematopoietic differentiation (Fig. 1B, Fig. S1B–D).
To delineate the successive development and emergence of hemato-endothelial progenitor cells (HEPs) CD34+/CD144+/CD45−, CD34+/CD144−/CD43+ cells and hematopoietic progenitors (HPs) CD34+/CD144−/CD45+ during hemanoid formation, we dissociated the hemanoids and analyzed the cells by flow cytometry. Among the CD34+ cells, HEPs were first detected at day 4 and 7 in our early hemanoids (74.5 ± 25.4%, 79.7 ± 6.0%, mean ± SD, n = 5, respectively), and a clear decrease at day 10 followed by gradual reduction during hematopoietic differentiation (47.0 ± 18.5%, mean ± SD, n = 5 and 31.8 ± 20.9%, mean ± SD, n = 4, respectively) was noted. Furthermore, we observed diminutive expression of CD34+/CD144−/CD43+ during earliest stage day 4 (1.0 ± 1.3%, mean ± SD, n = 5). The first appearance of HPs, more specifically, CD144−/CD43+ cells among the CD34+ cells was observed at day 7 and showed a gradual increase at day 10 and during hematopoietic differentiation (5.2 ± 3.4%, mean ± SD, n = 5 and 22.0 ± 22.8%, mean ± SD, n = 5, respectively). Interestingly, we observed CD34+/CD144−/CD45+ HPs at later stage than early CD34+/CD144−/CD43+ cells. The first appearance of CD34+/CD144−/CD45+ cells was observed at day 10 (20.2 ± 18.0%, mean ± SD, n = 5) and increased during further hematopoietic differentiation (4.1 ± 10.1%, mean ± SD, n = 4) (Fig. 1C, Fig. S1B).
Quantitative gene expression analysis of the entire hemanoids confirms a substantial early decrease in pluripotent marker OCT4, which subsequently decreased further at later stages. Transient expression SOX17, a transcription marker expressed in hemogenic endothelial cells, was noted in hemanoids at day 4–10. Additionally, expression of RUNX1 a marker required for the emergence of HPs during embryonic development and lymphoid/myeloid lineage maturation, appeared at day 7 and progressed during hemato-myeloid differentiation (Fig. 1D).
Taken together, we demonstrate that hiPSC-derived hemanoids undergo a stage-specific successive development, recapitulating key aspects of human embryonic hematopoiesis.
Phenotypic characterization of iPSC-Mac continuously produced in intermediate-scale bioreactorsTo test our differentiation process for robustness and efficiency of cell production, we subjected three different hiPSC lines to the recently developed, xeno-free, fully-defined and scalable protocol. After the formation of hemanoids in suspension culture by mesoderm priming, we observed the first production of macrophage-like cells released into the medium after approx. 10–14 days of cultivation in hematopoietic differentiation medium supplemented with IL-3 and M-CSF. In the first week of production, cells were harvested twice (harvest 1a and 1b), whereas in the consecutive weeks, cell harvest was performed along the media change on a weekly basis. Hemanoids derived from all three human iPSC lines continuously produced iPSC-Mac for 5–7 weeks (Fig. 2A) with comparable efficiencies of around 12 Million cells on average per harvest (iPSC line#1: 11.1 ± 9.6 Million cells, iPSC line#2: 11.9 ± 7.7 Million cells, iPSC line#3: 12.6 ± 16.6 Million cells, all mean ± SD, n = 14, 12 and 6 respectively). Importantly, harvested cells showed a classical and reproducible macrophage phenotype in brightfield as well as May-Grünwald/Giemsa stained cytospin images across the different harvests and the three hiPSC-lines (Fig. 2A, Fig. S2A). Additionally, immunophenotype analysis confirmed homogenous and characteristic surface expression of CD45 and CD11b (hematopoietic/myeloid), in addition to CD14, CD163, CD206 and CD86 (monocyte/macrophages) on iPSC-Mac derived from all three tested hiPSC lines, while CD66b, as a surface marker characteristic for granulocytes was absent (Fig. 2B, C, Fig. S2B, C). In this respect, iPSC-Mac from the consecutive weeks of differentiation showed a reproducible surface marker profile. Of note, we observed a lower expression of CD14 and more pronounced in CD163 positive cells in the first week of differentiation in comparison to later stages of differentiation (Fig. S2C).
Fig. 2
Phenotypic characterization of iPSC-derived macrophages continuously produced in intermediate scale bioreactors. A Number of viable cells harvested from the intermediate scale bioreactor for three different iPSC lines (iPSC line#1: n = 2, iPSC line#2: n = 2, iPSC line#3: n = 1, all individual values with mean +/− SD) over a time span of 7 weeks. Representative brightfield images and cytospin staining for iPSC-derived macrophages (iPSC-Mac) derived from harvest #1b, 3 and 5 for iPSC line#1. B Representative flow cytometry analysis of CD45, CD11b, CD14, CD163, CD206, CD86, HLA-DR and CD66b expression on iPSC-Mac from harvest #5. Histograms represent unstained iPSC-Mac (black line), and stained iPSC-Mac derived from iPSC line#1 (dark blue filled), iPSC line#2 (blue filled) and iPSC line#3 (light blue filled). Cells were pre-gated for viable cells according to FSC/SSC properties as well as single cells using SSC-A/SSC-H (see Fig S2B for gating strategy). C Frequency of CD11b+, CD14+ and CD163+ cells derived from different harvests/differentiations of the three different hiPSC lines as well as primary monocyte-derived macrophages. Individual values with mean ± SD, n = 5–15). Coefficient of variation (CV) is given for all values
When comparing our iPSC-Mac to primary macrophages derived from peripheral blood monocytes (MDM), we observed a much more homogenous and reproducible expression of CD14 and CD163, as important markers for macrophage differentiation on the hiPSC-derived cells (Fig. 2C). While CD11b and CD86 were also homogenously expressed on primary macrophages derived from different donors (Fig. S3), we observed a strong variability of CD14 and CD163 expression between the different donors/differentiations for primary cells, which is also indicated by a large coefficient of variation (CV) of 62.5% for CD14 and 70.6% CD163 expression compared to much lower CV values of 6.8–17.8% for iPSC-Mac derived from the different lines (Fig. 2C). Interestingly, we noted differences in the expression levels of the macrophage mannose receptor CD206 as well as HLA-DR between hiPSC-derived and primary macrophages. Here, our iPSC-Mac displayed lower expression of HLA-DR, but higher levels of CD206 expression, indicating a more anti-inflammatory phenotype upon harvesting (Fig. S2C).
Single cell transcriptomic analysis of iPSC-Mac from different hiPSC linesTo better define the iPSC-Mac and analyze the homogeneity of the population on a transcriptional level, we performed a scRNA sequencing analysis with iPSC-Mac from all three different lines, which were harvested at the same day from parallel running differentiations (all harvest #5).
After combination of the three sample sets and dimensionality reduction, we observed 5 different clusters in the Uniform Manifold Approximation and Projection (UMAP) analysis. Importantly, all iPSC-Mac samples from the different iPSC lines show cells belonging to all 5 clusters, though with variable frequencies. Here, cells allocated to cluster 0 and 3 were most prominently in the iPSC-Mac derived from iPSC-line#2, whereas cells allocated to cluster 2 were mainly found in iPSC-Mac derived from iPSC line#1 and cells allocated to cluster 1 were derived from iPSC-Mac derived from iPSC line#3 (Fig. 3A). Irrespectively of these differences, we observed, that clusters 0–3 all demonstrate a clear macrophage phenotype indicated by the expression of ITGAM/CD11b, CD14, CD163, CD86, and MRC1/CD206, as also previously observed by flow cytometry. Also, the absence/very low expression of HLA-DR was confirmed. Here, only cells allocated to the small cluster 4 showed absence of these important macrophage genes, indicating some few contaminating cells (Fig. 3A, B). Similar results were obtained, when analyzing typical macrophage genes (conserved macrophage genes [40]). Also, here we noticed a strong expression of these key genes over clusters 0–3, whereas low to no expression was detected in cluster 4 (Fig. 3C). To confirm the cell identity, we performed logistic regression (LR) analysis using publicly available single cell transcriptomics data. Strikingly, we observed expression related to the class reference of macrophages and residual expression related to mono-DC precursor and monocytes. Within the macrophage reference class, the highest expression was observed in clusters 1, 2, 3, 0 and 4, respectively (Fig. 3D). Given the primitive fingerprint of several iPSC-derived progeny, which has also been reported for iPSC-derived macrophages [41, 42], we also analyzed the expression of genes associated with primitive yolk sac macrophages [36]. Indeed, we observed expression of LYVE1, NID1, LIN28B, IGF2BP1, CALD1, FGF13, SERPINH1, FERMT2 and PARD3 in clusters 0–3 (Fig. S4). To better understand the differences in clusters 0–3, we analyzed the expression patterns of genes, which are associated with macrophage activation/polarization. Here, we observed some differences between the clusters. While all of the clusters demonstrated expression of genes associated with pro- and anti-inflammatory activation, especially cluster 1 and 2 showed a more abundant and stronger expression of genes associated with a pro-inflammatory M1 macrophages activation status such as MX1, STAT1, or HIF1A. On the other hand, STAB1, a marker gene for anti-inflammatory or primitive/tissue resident macrophages [43] as well as RNASE1, which was recently described as a M2 macrophage marker [44] was highly expressed in cluster 0 and 3 (Fig. 3E–G). However, the clusters do not reflect a black and white picture of macrophage polarization, but demonstrate co-expression of key genes associated with the both directions. This is also indicated by a strong expression of APOE in cluster 1, which has been shown to induce an anti-inflammatory macrophage phenotype [45] and may indicate an auto-regulatory feedback loop.
Fig. 3
Single Cell transcriptomic analysis of iPSC-Mac from different hiPSC lines. A Uniform Manifold Approximation and Projection (UMAP) representing unsupervised clustering overlaying hiPSC lines #1, #2, and #3. Additional UMAP representation for each iPSC line. Bar chart representation demonstrating the ratios of each cluster in each line independently. B Violin plots demonstrating expression of ITGAM, CD14, CD163, CD86, MRC1/CD206, HLA-DRA for each cluster. C Expression of conserved lineage markers specific to macrophages (adapted from [40]) within the different clusters. D Heatmap representing the predicted probabilities of cell types. Cluster annotations were predicted using logistic regression classifiers trained on publicly available data [56]. E Gene list of macrophage polarization state M1, M2a, M2b and M2d (adapted from [57]). F Gene list of IFNγ fingerprint (adapted from [52]). G UMAP demonstrating global expression of pro and anti-inflammatory genes (APOE, MX1, RNASE1, STAB1) H Paga plot velocity graph annotate clusters M1, interim, and M2 cell states of macrophages. Bar chart representation demonstrating the ratios of M1, M2 and interim population produced from each line independently
These data point out, that indeed most of the harvested and non-purified cells, represent a core macrophage transcriptome (cluster 0–3). However, specifically looking at the different macrophage activation stages, the clusters differ in abundance and expression level of specific genes associated with pro- and anti-inflammatory macrophage functions. Given the different distribution of cells between the clusters in iPSC-Mac derived from the different iPSC lines, we observe a trend towards a slightly more pro-inflammatory activation in iPSC-Mac derived from hiPSC line#1 and 3 (cluster 1 and 2 show a higher frequency of cells), while iPSC-Mac derived from iPSC line#2 displayed more of anti-inflammatory signature (cluster 0 and 3 show a higher frequency of cells) (Fig. 3G). Given the observation of various polarization genes signatures, our aim was to analyze the clusters using RNA velocity and trajectory analysis to identify the various cell states. Notably, we observed directionality from cluster 2 toward cluster 3, then to cluster 0. Additionally, cluster 2 exhibited movement toward cluster 1, suggesting the presence of three distinct cell states. We noted higher expression of genes related to an M1-like signature in clusters 1 and 2. Conversely, expression of genes related to M2-like signatures in clusters 0 and 3 seemed to represent a combination of M2-like genes with lower expression of M1 genes and cluster 0 with the lowest expression level (Fig. 3E–G). Annotating the cell clusters based on gene expression and trajectory analysis, we identified cluster 0 as an M2-like phenotype, cluster 1 and 2 as M1-like, and cluster 3 as an interim between M1 and M2. We observed the highest M1-like cell frequency in Line#3 (86.3%) followed by Line#1 (77.7%) and Line#2 (9.64%). M2-like and interim cell frequencies was the highest in line#2 (59.5% and 28.6%) followed by line#1 (16.5% and 2.55%) and line#3 (8.6% and 1.46%), respectively (Fig. 3H).
Generated iPSC-Mac demonstrate important pro-inflammatory functionalityAfter showing broad phenotypic similarities between the iPSC-Mac derived from different harvests, differentiations, and hiPSC lines, we next aimed to characterize their functional properties. Given the important role of macrophages in cellular host defense, we analyzed their potential to phagocytose bacterial particles, to produce reactive oxygen species (ROS) and secrete Interleukin 6 (IL-6) in response to a pro-inflammatory stimulus.
After 2-h incubation with pHrodo™ Red E. coli BioParticles, we observed a strong phagocytic activity of iPSC-Mac with 93.4 ± 8.2% pHrodo™ Red positive cells for iPSC line#1, 83.0 ± 11.1% for iPSC line#2 and 90.0 ± 4.9% cells for iPSC line# 3 (n = 6–15). In contrast, we only observed a phagocytosis rate of 71.7% for MDMs, again with a strong variability indicated by a SD of 36.4% (n = 5) and a CV of 50.75% (Fig. 4A, Fig. S5A).
Fig. 4
iPSC-Mac demonstrate important pro-inflammatory functionality. A Phagocytosis of pHrodo™ Red E. coli BioParticles. Different iPSC-Mac as well as primary Mac were incubated with pHrodo™ Red E. coli BioParticles for 2 h at 37 °C. Subsequently, phagocytosis was evaluated by the induction of a red fluorescent signal after acidification of the pH-sensitive pHrodo™ Red in the phagolysosome. Left: Representative fluorescence microscopy of iPSC-Mac derived from iPSC line#1 incubated for 2 h with pHrodo™ Red E. coli BioParticles (fluorescence only, brightfield as well as overlay, scale bar = 100 µm). Right: Frequency of pHrodo Red+ cells derived from different harvests/differentiations of the three different hiPSC lines as well as primary monocyte-derived macrophages analyzed by flow cytometry (Individual values with mean ± SD, n = 5–15). B Production of reactive oxygen species (ROS) by macrophages from the different sources. Different iPSC-Mac as well as monocyte derived macrophages (MDM) were incubated with PMA for 5 min and subsequently stained with Dihydrorhodamine (DHR). Left: Representative flow cytometry data for iPSC-Mac derived from the different iPSC lines#1–3 as well as primary Mac (grey: unstimulated; stained and colored filled: respective macrophages stimulated with PMA and stained). Right: Fold change of Rhodamine mean fluorescence intensity (MFI) for iPSC-Mac derived from different harvests/differentiations of the three different hiPSC lines as well as primary monocyte-derived macrophages analyzed by flow cytometry (Individual values with mean ± SD, n = 4–11, dotted line indicates “1”), C Secretion of IL-6 after stimulation with Lipopolysaccharide (LPS). Different iPSC-Mac as well as primary Mac were stimulated with 500 ng/ml LPS for 4 h and supernatants were analyzed for secretion of IL-6 by ELISA. Left: IL-6 levels secreted by iPSC-Mac from the three different iPSC lines for the individual harvests. Right: IL-6 secretion for iPSC-Mac derived from different harvests/differentiations of the three different hiPSC lines as well as primary monocyte-derived macrophages (Individual values with mean ± SD, n = 3–15). Statistical analysis was performed using one-way ANOVA with Tukey’s multi comparisons test (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, ns denotes not significant)
Stimulation with phorbol 12-myristate 13-acetate (PMA) lead to the production of ROS, indicated by an increase in Rhodamine mean fluorescence intensity (MFI) in both, iPSC-Mac and MDMs. While all different macrophages responded to PMA stimulation with an increased Rhodamine signal/ROS production, there was a general trend towards a higher induction of ROS production in the iPSC-Mac, specifically in hiPSC line#1 and 3 that showed significantly higher induction of ROS when compared to MDMs (Fig. 4B, Fig. S5B).
After stimulation with Lipopolysaccharides (LPS) for 4 h, we observed a significant induction of IL-6 secretion, indicating an efficient pro-inflammatory response of the iPSC-Mac. While we observed some differences in the level of IL-6 secreted after LPS stimulation of the iPSC-Mac derived from the different hiPSC-lines, iPSC-Mac from the different harvests of the same differentiation responded in a reproducible manner towards this stimulus. In general, we observed a higher induction of IL-6 secretion in iPSC-Mac derived from all the three lines when compared to primary macrophages (Fig. 4C).
Taken together, all iPSC-Mac displayed typical macrophage functionality and important anti-bacterial functions when compared to primary cells. Moreover, iPSC-Mac derived from the different harvests or differentiations produced on the intermediate scale, continuous production platform showed a high-quality reproducibility of phenotype and function.
iPSC-Mac can be polarized into different pro- and anti-inflammatory activation stagesGiven the diverse function of macrophages in innate immunity as well as tissue homeostasis and repair, a crucial feature is their responsiveness to different pro- or anti-inflammatory stimuli and a change of their activation status. Thus, we next analyzed the potential of our iPSC-Mac to adopt pro- or anti-inflammatory activation stages by stimulation with IFNy (M1), IL-4 (M2a) and IL10/TGFb (M2c) (Fig. 5A). Indeed, all iPSC-Mac derived from the three hiPSC lines responded to a pro-inflammatory IFNy stimulation with a specific up-regulation of Fc receptor CD64 as well as increased HLA-DR expression (Fig. 5B, Fig. S6, Supplementary Table 1). The anti-inflammatory stimulation with IL-4 resulted in a profound up-regulation of the T cell co-stimulatory molecule CD86 as well as the macrophage mannose receptor CD206 (Fig. 5B, Fig. S6, Supplementary Table 1), which are both associated with an anti-inflammatory activation stage. The alterations in the surface marker profile after different stimuli were associated with the induction of cytokine secretion (Fig. 5C). Here, we observed a characteristic pattern of cytokine induction across the iPSC-Mac from all hiPSC lines: IFNy induced strong induction of C-X-C motif chemokine ligand 10 (CXCL10)/ Interferon gamma-induced protein 10 (IP-10) secretion. IL-4 stimulation resulted in the specific up-regulation of Chemokine (C–C motif) ligand 12 (CCL12), and IL-4 as well as IL10/TGFb stimulation lead to the up-regulation of IL10 secretion. Whereas the pattern of cytokine induction was the same for all different lines, we observed differences in the levels of cytokine secretion. The iPSC-Mac derived from hiPSC line#1 and 3 showed only low background levels of IP10 secretion in the non-stimulated controls (558.12 ± 376.78 pg/ml, mean ± SD, n = 4 and 657.62 ± 413.87 pg/ml, mean ± SD, n = 3, respectively). In contrast, iPSC-Mac from line#2, even under steady-state conditions exhibited profound levels of IP10 secretion of 3350.21 ± 2271.28 pg/ml (mean ± SD, n = 3). Additionally, variations in CCL17 secretion levels were observed after IL-4 stimulation among the different lines. Specifically, iPSC-Mac from iPSC line#1 and 3 revealed CCL17 levels ranging from 200 to 260 pg/ml, while iPSC-Mac from line#2 demonstrated a higher secretion level of 1015 pg/ml (Fig. 5C).
Fig. 5
Polarization of iPSC-Mac into different pro- and anti-inflammatory activation stages. A Schematic representation of the experimental layout. iPSC-Mac from the three iPSC-lines were polarized in vitro by the stimulation with 25 ng/ml IFNy into pro-inflammatory M1(IFNy) macrophages or with 10 ng/mL IL-4 or IL-10/TGFb into anti-inflammatory M2(IL-4) or M2 (IL10/TGFb) iPSC-Mac. B Changes in surface marker expression of CD64, HLA-DR, CD86 and CD206 24 h after polarization analyzed by flow cytometry. Values are given as fold change in the median fluorescence intensity (MFI) compared to non-stimulated cells (Individual values with mean ± SD, n = 4 for iPSC line#1, n = 3 for iPSC line#2 and 3). Statistical analysis was performed using one-way ANOVA with Tukey’s multi comparisons test (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, ns denotes not significant). C Secretion of IP10, CCL17 and IL10 24 h after polarization analyzed by Legendplex technology (Individual values with mean ± SD, n = 4 for iPSC line#1, n = 3 for iPSC line#2 and 3, dotted lines depict upper (IP10) or lower (CCL17) detection limits)
Thus, all iPSC-Mac showed characteristic changes after exposure to pro- and anti-inflammatory stimuli and the adaptation of M1 or M2 phenotypes. However, the iPSC-Mac from the different lines displayed some variability in the levels of cytokine secretion.
iPSC-Mac as a model system for the testing of immune-modulatory drugsEnsuring the validity and reproducibility of model systems is crucial for testing the efficiency of novel immunotherapeutic strategies. As an alternative to artificial cell line models or primary cells, we evaluated the potential of our iPSC-Mac to show a dose-dependent response to LPS stimulation. More importantly, we examined the effect of the classical anti-inflammatory drug dexamethasone on the LPS-induced secretion of IL-6.
All iPSC-Mac exhibited a dose-dependent response to increasing concentrations of LPS, as indicated by increasing secretion levels of IL-6 (Fig. 6). However, variations in the overall levels of secreted IL-6 were also observed. When stimulated with 500 ng/ml LPS, iPSC-Mac derived from line#2 showed lowest IL-6 levels with 1122.34 ± 67.54 pg/ml, iPSC-Mac from line#1 levels of 2352.83 ± 52.79 pg/ml and iPSC-Mac from line#3 the highest levels with 2989.56 ± 641.52 pg/ml. In comparison, primary macrophages displayed secretion levels of up to 1885.19 ± 1033.05 pg/ml. Irrespectively of these different levels of IL-6 secretion, all hiPSC lines showed a significant suppressive effect of dexamethasone on IL-6 secretion in a specific dose range of 10–100 ng/ml LPS stimulation (p = 0.0004 iPSC line#1 at 100 ng/ml LPS stimulation, p = 0.0032 iPSC line#2 at 10 ng/ml LPS stimulation and p = 0.0017 iPSC line#3 at 10 ng/ml LPS stimulation). Furthermore, we noted a significant effect of dexamethasone at 100 ng/ml LPS stimulation in MDMs, albeit with a larger p value of 0.0225 (Fig. 6).
Fig. 6
iPSC-Mac as a model system for the testing of immunodulatory drugs. Different iPSC-Mac as well as primary Mac were stimulated with increasing concentrations of LPS (0, 1, 10, 100 and 500 ng/ml) with or without the simultaneous addition of 1 ug/ml Dexamethasone for 4 h. Levels of IL-6 secretion were determined in supernatants using ELISA. (Individual values with mean ± SD, n = 3. Statistical Analysis was performed using two-way ANOVA with Sidak’s multi comparisons test (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, ns denotes not significant)
In conclusion, these data illustrate that iPSC-Mac serve as a suitable model system for studying the effect of immune-modulatory drugs. Nevertheless, these findings highlight variations between the hiPSC lines in the optimal dose of stimulation.
留言 (0)