
記住我





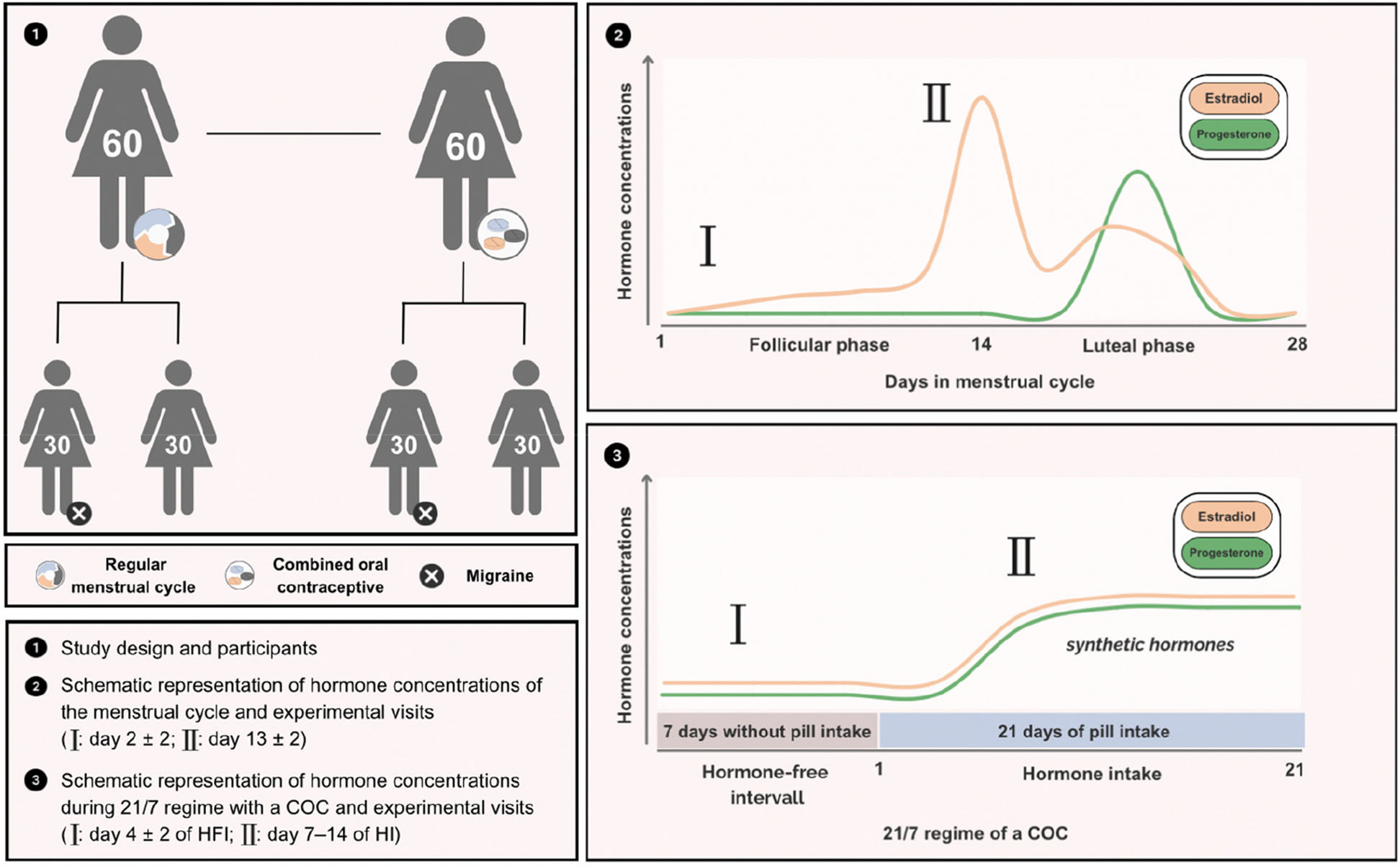
Our randomized, placebo-controlled study discovered that MaxiPost, a BKCa channel opener, can induce migraine-like headache in persons with PPTH who reported no pre-mTBI history of migraine. Importantly, the participants reported that their spontaneous migraine-like headache closely mirrored the one induced by MaxiPost. This observation bolsters our assertion that BKCa channel opening can generate cephalic pain, particularly migraine-like headache, among persons with PPTH. Thus, our findings pave the way for the development of BKCa channel blockers as a novel approach to address unmet treatment needs in managing PPTH.
BKCa Channel Involvement in Headache DisordersThe contribution of BKCa channels to the pathogenesis of headache disorders has garnered increasing attention in recent years [17]. Randomized, placebo-controlled studies have found that intravenous infusion of MaxiPost can induce migraine attacks in persons with migraine [6]. In contrast, healthy individuals only experience non-migraine-like headache following MaxiPost infusion [5]. Moreover, experimental studies have shown that intravenous infusion of MaxiPost results in decreased blood flow velocity within the middle cerebral artery [5, 6]. This observation holds true for both persons with migraine and healthy individuals [5, 6]. Thus, BKCa channel openers appear to dilate intracranial arteries, a physiologic effect that is consistent across all known triggers of migraine-like headache [12, 22, 23]. Although it might seem intuitive that MaxiPost can induce migraine-like headache in people with PPTH, given that it induces migraine attacks in people with migraine, this assumption is not necessarily universally applicable across molecular headache triggers. For instance, despite CGRP being a well-known migraine inducer [10, 24, 25] it does not trigger migraine attacks in people with familial hemiplegic migraine [26, 27]. Likewise, a KATP channel opener, levcromakalim, induced migraine attacks in all participants with migraine without aura, [28] but the incidence of migraine-like headache in people with PPTH and cluster attacks in those with cluster headache, is clearly lower [21, 29]. This differential response is also evident in our findings, where 52% of participants with PPTH experienced migraine-like headache post-MaxiPost infusion, as opposed to a 95% induction rate in people with migraine [6]. Given the identical dosage and infusion durations used across both studies, it is plausible that BKCa channel opening plays a less prominent role in the pathogenesis of PPTH, compared with migraine. Furthermore, the molecular and cellular mechanisms underlying PPTH could differ from those of migraine, which might account for the varied responses to the same molecular headache trigger. Taken together, there is evidently variability in sensitivities and expression of different headache phenotypes in response to molecular headache triggers. This underscores the intricate interplay of molecules in headache pathogenesis, which goes beyond specific headache phenotypes. It is also important to recognize the challenge of a priori predicting individual responses to molecular headache triggers across different headache disorders. Therefore, randomized, placebo-controlled studies are essential for an in-depth evaluation of these responses. Taken together, the involvement of BKCa channels seem to cover a plethora of physiological processes, including regulation of intracranial vasculature, neuronal excitability, and pain transmission, among others.
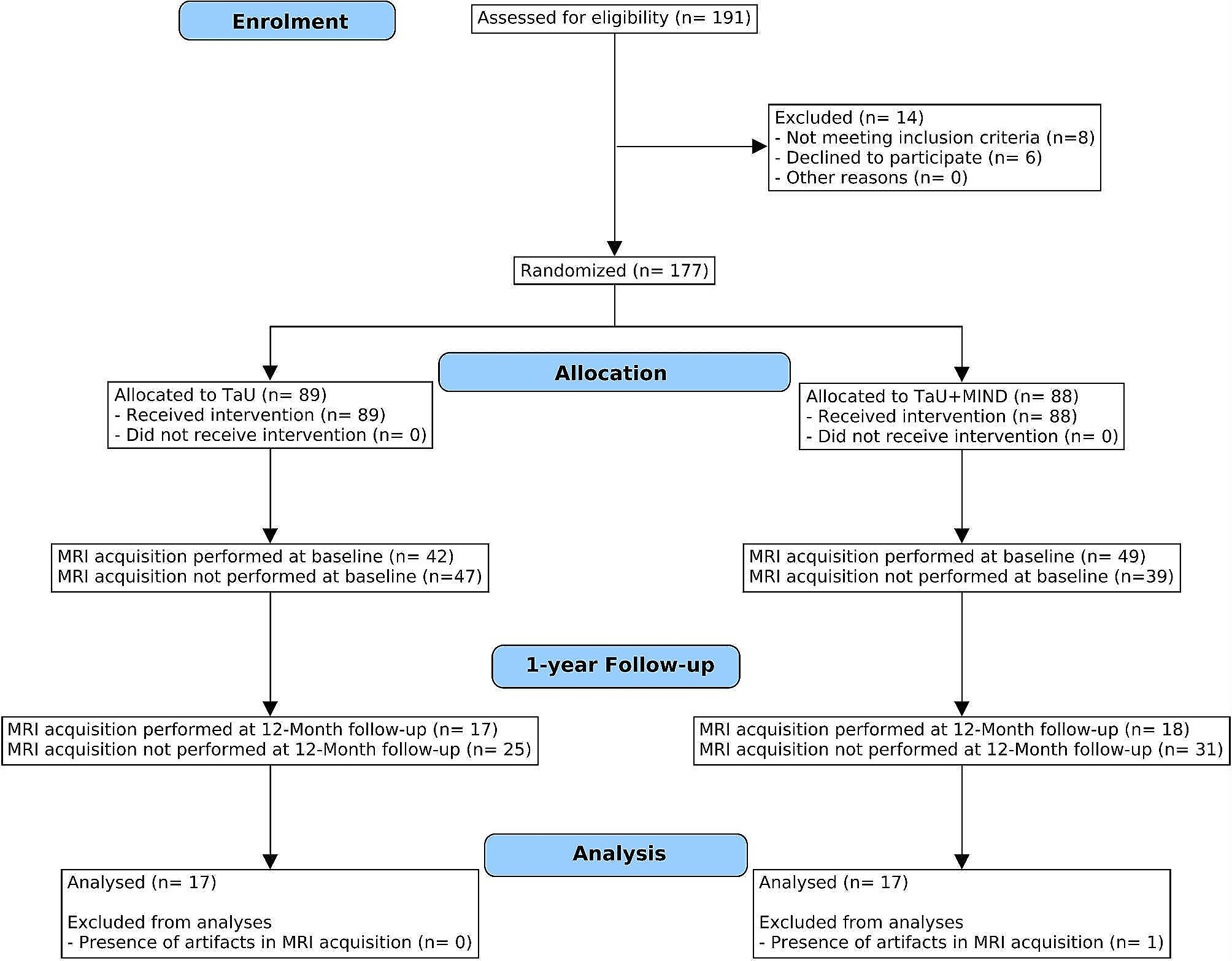
Proposed sites and mechanisms of actionThe site(s) and mechanism(s) through which the activation of BKCa channels culminate in migraine-like headache continue to be areas of active exploration. One plausible site of action is the meninges and its vasculature (Fig. 3). Prior research indicates that MaxiPost promotes dilation of intracranial arteries in both persons with migraine and healthy individuals [5, 6]. This vasodilatory effect is corroborated by preclinical data, demonstrating a causal link between the BKCa channel opening and dilation [30]. A proposed mechanism involves the efflux of potassium ions from VSMCs, leading to vasodilation [17].17 The rationale is that increased extracellular levels of potassium, in part, due to BKCa channel opening offer electrochemical stimuli, while the concurrent vasodilation provides mechanical stimuli. Together, these stimuli might have the potential to activate perivascular nociceptors. The culmination of this cascade is hypothesized to result in migraine-like headache [18, 19]. This proposed sequence of events aligns well with emergent evidence indicating that administration of CGRP and PACAP, both potent vasodilators of meningeal arteries, [22, 31] can induce migraine-like headache in persons with PPTH [11, 13]. It is worth noting that this vasodilatory response is mediated, in part, through the activation of BKCa channels [17]. Taken together, future rodent experiments should ascertain whether BKCa channel opening within the meningeal vasculature can indeed activate perivascular nociceptors.
Fig. 3
Possible Mechanisms and Sites of Action of Migraine-Like Headache induced by BKCa Channel Opening in Persons with PPTH. The figure outlines a proposed mechanism and site of action though which opening of BKCa channels contributes to the development of migraine-like headache in persons with PPTH. In this suggested model, the signaling molecules CGRP and PACAP-38 bind to their respective G protein-coupled receptors present on the vascular smooth muscle cells of intracranial arteries. This initiates the activation of cAMP-dependent signaling pathways, which then results in opening of BKCa channels. The subsequent release of potassium ions and accompanying vasodilation leads to activation and sensitization of perivascular meningeal nociceptors, a process facilitated by both chemical and mechanical stimulation. Modified from Al-Khazali et al., 2023 [21]. CGRP = calcitonin gene-related peptide; PACAP-38 = pituitary adenylate cyclase-activating polypeptide-38; AC = adenylate cyclase; ATP = adenosine triphosphate; cAMP = cyclic adenosine monophosphate; BKCa channels = large conductance calcium-activated potassium channels
An alternate, albeit less probable, site of action might be the primary afferent fibers whose cell bodies are located in the trigeminal ganglion and upper cervical ganglia. These fibers are responsible for the nociceptive innervation of the meninges, including its vasculature [32]. Opening of the BKCa channels causes the membrane potential to become more negative, causing a hyperpolarization of the afferent fibers. The implication is then that administration of a BKCa channel opener, such as MaxiPost, will exert anti-nociceptive effects, which is incongruent with our findings. However, a possible explanation might be that BKCa channel opening on the primary afferent fibers can activate hyperpolarization-activated cyclic nucleotide-gated (HCN) channels. This, in turn, can result in depolarization and possible nociceptor activation [33,34,35]. Nonetheless, this hypothesis remains speculative and less convincing in light of preclinical evidence showing that iontophoretic application of a BKCa channel opener inhibits primary afferent fibers. In addition, preclinical data suggest that MaxiPost’s direct effect on nociceptive neurons within the CNS is unlikely, as bath application of a BKCa channel opener inhibited nociceptive responses in 2nd order trigeminal neurons, whereas blocking these channels facilitated nociceptive responses [36].
Furthermore, the human provocation model has shown that CGRP infusion not only triggers migraine headache but also its accompanying symptoms [10, 24, 25]. Conversely, antibodies not only reduce headache and migraine days but also reduce the accompanying symptoms of migraine [37,38,39]. The mechanism is not yet known. Preclinical studies have shown that CGRP's downstream effect includes phosphorylation and opening of BKCa channels, which is why BKCa opening with Maxipost is also assumed to have a similar effect by activating these accompanying symptoms [40]. Al-Kharagoli et al. also found a significant difference in the incidence of accompanying symptoms (i.e, photophobia, phonophobia, and nausea) with Maxipost compared to placebo among migraine patients [6]. Therefore, it could be hypothesized that blocking BKCa channels would likely also inhibit the induction of these symptoms.
Therapeutic implications and future directionsOur results lend support to the pivotal role of BKCa channels in the pathogenesis of PPTH. If the opening of BKCa channels can trigger head pain, it is plausible that blocking these channels might alleviate the pain. The development of BKCa channel blockers might thus represent a novel and targeted approach to treating PPTH. However, while our findings are promising, they represent only the first step towards novel drug discovery. BKCa channels are known for their ubiquitous expression across various tissues, including those implicated in the genesis of head pain, such as the intracranial arteries and trigeminal ganglion [40, 41]. The development of effective and safe BKCa channel blockers will therefore require extensive research. In this context, it will be important to explore the exact sites and mechanisms of BKCa channel action within the neurobiological underpinning of PPTH.
An issue that merits special emphasis is when to initiate pharmacologic treatment with a BKCa channel blocker in persons with PPTH. Here, one might consider that CGRP’s downstream action in VSMCs involve BKCa channel opening, and inhibition of CGRP signaling can attenuate cutaneous allodynia in concussed rodents. This effects appears, however, to decrease over time, suggesting a time-sensitive window for optimal pharmacologic treatment post-concussion. This time-bound phenomenon might be intimately tied to diffuse noxious inhibitory controls (DNIC), a physiological regulator of descending pain modulation [42]. A disruption in DNIC function, as seen in concussed rodents, might reduce the brain’s ability to modulate ascending pain signals, thereby leading to the development of persistent and chronic headache. Here, it warrants mention that the loss of DNIC function can be prevented in concussed rodents after inhibition of CGRP signaling [43]. The question is then whether a BKCa channel blocker could replicate similar outcomes in these rodent models. Should this be the case, it suggests the importance of early treatment with a BKCa channel blocker after mTBI for the management of PPTH.
LimitationsThis study has some limitations that warrant mention. First, the in-hospital observation period extended only to one hour following the infusion due to logistical considerations. Thus, we cannot exclude the potential influence of various environmental factors, such as dietary intake or stress levels, on the data collected throughout the remaining 11 hours of the observation period. Second, the use of rescue medication presents another limitation, albeit a minimal one given its infrequent usage among our participants to alleviate headache symptoms. Third, the use of a placebo in our two-way crossover design directly addresses the concern regarding the potential overlap between spontaneous and MaxiPost-induced migraine-like headache in our participants. Using placebo, we can effectively differentiate between migraine-like headache that occurs spontaneously and those induced by MaxiPost. This design allows each participant to serve as their own control, thereby mitigating the influence of natural headache fluctuations. Hence, we can more accurately attribute differences in headache responses to the effects of MaxiPost, rather than natural variations in headache occurrence. Fourth, ongoing use of preventive headache medication might have mitigated the headache response after MaxiPost infusion. This highlights the need for further research to elucidate the potential impact of these variables on the initiation and evolution of migraine-like headache in PPTH. Fifth, in addition to Maxipost's primary role in opening BKCa channels, it also modulates GABAA receptors and neuronal Kv7 channels [44]. These interactions are expected to mitigate neuronal hyperexcitability, suggesting potential therapeutic applications in pain management. Yet, human experimental studies have clearly demonstrated that MaxiPost is a potent molecular headache trigger [5, 6]. It remains uncertain whether modulation of GABAA receptors and neuronal Kv7 channels contribute to or counteract MaxiPost’s ability to trigger headache. This adds a layer of complexity in interpreting our findings, and further research is required to clarify the roles of GABAA receptors and neuronal Kv7 channels in headache pathogenesis. Lastly, it cannot be excluded that some participants might be predisposed to migraine, which was then elicited in response to head trauma. Their migraine-like headache might, therefore, represent a genuine migraine attack. According to the ICHD-3 [20], people with migraine can experience an exacerbation of their typical migraine attacks after head trauma. It is, however, important to note that our participants reported no personal history of migraine and developed acute PTH within seven days of the head trauma. Further research is warranted to determine whether head trauma can indeed ‘activate’ a latent predisposition to migraine, a topic that has garnered attention in recent literature [45, 46].
留言 (0)