
記住我
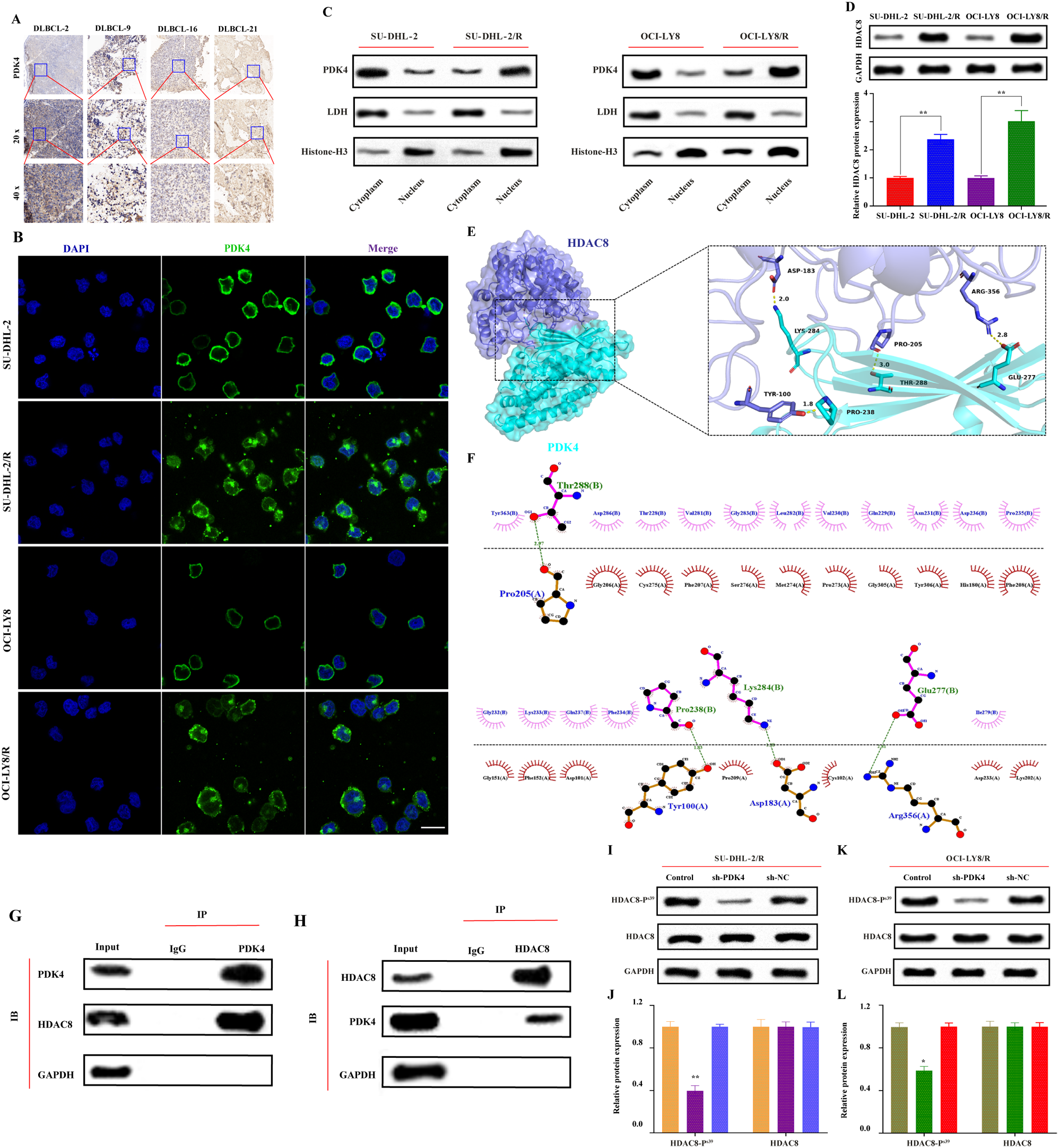


We aimed to identify clinically relevant genes that mediate palbociclib sensitivity by using an in vivo genome-scale CRISPR/Cas9 loss-of-function screen in a preclinical model of TNBC. We used an Rb-proficient human SUM159PT TNBC cell line [13]. We selected SUM159PT because it is (i) a well-established tumorigenic and metastatic model in vivo (ii) Rb + [14] and thus intrinsically sensitive to CDK4/6 inhibitor treatment, and (iii) representative of TNBC as it harbors PIK3CA and TP53 mutations, two of the most frequently observed mutations in TNBC [15, 16]. As illustrated in Fig. 1a, SUM159PT cancer cells were transduced with the lentiviral pooled genome-scale CRISPR/Cas9 knockout (KO) GeCKOv2 library. GeCKOv2 covers the whole genome with three single guide RNAs (sgRNAs) for each of the 19,050 target genes and 1000 non-targeting control sgRNAs [17, 18]. A low multiplicity of infection (MOI ~ 0.3) was chosen to ensure the integration of only one sgRNA per cell. Due to the sheer number of cells to be transduced, and the complexity of delivering perturbation reagents directly in the host organs of a large number of mice that would have been required to perform a direct in vivo screen, an indirect screen was chosen. Stable knockout cells were thus injected subcutaneously (s.c.) into severely immunodeficient NOD scid gamma (NSG) mice at approximately 400-fold library coverage for each animal in each of the three independent experiments. Tumors were allowed to grow for seven days, until palpable. Mice were then randomized and subjected to intraperitoneal injections of either vehicle or 30 mg/kg palbociclib once daily for five days/week for 23 days. Tumor volume was monitored over the entire 30-day duration of the experiment. Exposure of GeCKO-derived tumors to palbociclib effectively reduced tumor size, illustrating the potency of palbociclib when administered in the in vivo TNBC setting (Fig. 1b). The cell representation samples were sequenced on the day during which the cells were transplanted subcutaneously in mice, to examine the evenness of the library representation. The cell population at day 0 harbored a 99% library representation, indicative of an excellent library coverage (data not shown). Sequencing of tumors revealed a high degree of reproducibility, as demonstrated by the close grouping of principal component analysis (PCA) (Fig. 1c) in six same-condition in vivo biological replicates. PCA again highlighted the relative separation of sgRNA distribution between the untreated and palbociclib-treated samples (Fig. 1c). sgRNAs that were enriched or depleted after in vivo screening under palbociclib selection pressure were then identified. Enriched sgRNAs in palbociclib-treated tumors define genes conferring sensitivity to palbociclib, where loss-of-function mutations in these genes increase overall cell resistance to drug treatment and would thus present novel markers predictive of the palbociclib response. While we did not obtain any significantly depleted sgRNAs, a total of 205 candidate sgRNAs were positively enriched in the palbociclib-treated tumors (Fig. 1d). The sgRNA enrichment profile was generated by filtering sgRNAs with false discovery rate (FDR) < 0.05. Any sgRNAs with fewer than 10 control reads were dropped from the analysis to ensure screen quality and reduce the potential for false positive hits. Gene ontology pathway enrichment analysis performed on the 205 gene list revealed no significantly enriched gene sets or pathways.
Fig. 1
In vivo genome-wide CRISPR knockout screen in TNBC. a Schematic representation of the approach used for gene discovery and validation. b Average tumor volume in NSG mice measured over 30 days. Intraperitoneal (i.p.) injections of either vehicle or palbociclib started on day 7 post-cell implantation, and lasted 23 days. Mean of three independent infection replicate experiments (n = 6, 2 mice per biological replicate). Data are represented as mean ± standard deviation (SD). Significance was calculated using two-sided, unpaired t-test, p-value * < 0.05, ** < 0.01, *** < 0.001. c Principal component analysis (PCA) of the sgRNAs from the library sequenced in vehicle-treated tumors (n = 6), and palbociclib-treated tumor samples (n = 6) at day 30 after normalization. d 205 sgRNAs were enriched with log2-fold change (LFC) > 0 at false discovery rate (FDR) < 0.05 in palbociclib-treated tumors during the screen. Genes representing significant hits are highlighted in red. e Palbociclib sensitivity data was used to rank 38 breast cancer cell lines of varying subtypes, generating two profiles of cell lines, ‘sensitive’ and ‘resistant’. GSEA was used to determine whether 205 sgRNA gene set was significantly enriched in either group of cell lines. Enrichment plot provides the distribution of the enrichment score (green line) of the 205-gene set in the ranked cell lines (sensitive to resistant, left to right). The final, positive normalized enrichment score (NES) at 1.288 indicates significant enrichment of the 205-gene set at FDR < 0.25 in palbociclib ‘sensitive’ cell lines (FDR = 0.0568, p-value = 0.0568). f Using GSEA, expression levels of the 47 genes (core enrichment subset) are presented here. Cell lines are annotated with clinical information
To shortlist candidate genes that could best predict palbociclib sensitivity in TNBC, we next cross-referenced our CRISPR screen gene dataset with microarray data from a panel of 38 breast cancer cell lines with varying sensitivities to palbociclib [19]. Cell lines were ranked from most to least sensitive based on palbociclib IC50 values determined in Finn and colleagues' study [7] and correspondingly divided into two groups: ‘more sensitive’ and ‘less sensitive’ to palbociclib. Using Gene Set Enrichment Analysis (GSEA), we sought to determine if the gene set obtained by our screen was enriched in the ‘more sensitive’ cell lines sorted by sensitivity to palbociclib (IC50) [20]. As expected, our 205-gene set was significantly upregulated at FDR < 0.25 in cell lines which are sensitive to palbociclib (FDR = 0.0568) (Fig. 1e). The ‘more sensitive’ cell lines expressed higher levels of genes in our gene set, underscoring the power of our screen to identify genes predictive of palbociclib efficacy across a broad landscape of breast cancer subtypes (Suppl. Figure 1a). Of this gene set, 47 genes formed the ‘core enrichment subset’ as defined by GSEA; genes which contributed most to the positive normalized enrichment score (NES) generated for the entire gene set [20, 21].
We hypothesized that this subset would therefore have the strongest association with palbociclib effectiveness and could serve as a predictive gene signature for palbociclib sensitivity and overall clinical outcomes in patients. We associated the 38 cell lines used in the GSEA with corresponding clinical information. As expected, this cell line ranking coincided with clustering of cell lines based on Rb proficiency, hormone receptor (HR)/HER2 status, and molecular subtype classification, such that known CDK4/6 sensitivity phenotype criteria were fulfilled (Fig. 1f) [22,23,24]. Indeed, Rb-deficient cell lines clustered together in the ‘less sensitive’ subgroup, as did most cell lines representing the basal subtype of breast cancer. Conversely, HR + and HER2 + cell lines, and cell lines of luminal or HER2 molecular subtype, largely clustered in the ‘more sensitive’ subgroup (Fig. 1f). These findings contributed to our confidence in the screening and the prioritization methods used as they allowed us to situate our results in the context of what is already known. Nonetheless, these results also help strengthen our rationale for the study, showing that palbociclib sensitivity is not simply dictated by ER status or Rb mutation status during patient stratification. We next sought to evaluate whether the 47-gene core enrichment subset could serve as a predictive gene signature for palbociclib sensitivity and overall clinical outcomes in publicly available data sets. We evaluated these genes’ expression patterns in a cohort of patients with breast invasive carcinoma (METABRIC) using cBioPortal [25,26,27]. We observed a trend towards a decrease in gene expression in the HR-/HER2- (TNBC) subgroup, as compared to the other groups classified by their expression of HR and HER2, although this was not significant (Suppl. Figure 1b). A seemingly lower expression of the 47-gene signature was also observed in the more aggressive basal and claudin-low groups of patient samples, and tended to correlate with higher tumor grade, although this was not significant (Suppl. Figure 1c, d). Taken together, the significant upregulation of the 205-gene set obtained from our in vivo CRISPR/Cas9 screening in the 20 ‘more sensitive’ cell lines underscores the power of the screen to reliably and robustly identify markers of drug effectiveness. These findings strengthen the predictive power of the gene signature defined using our prioritization method, showing that overall lower expression of genes here correlates with poorer clinical outcomes in general, while also promoting palbociclib resistance.
Having evaluated the clinical relevance of the 47-gene signature using patient data, we next assessed these genes' ability to modulate the palbociclib response in vivo, using TNBC xenograft models. For this, the eight top-ranking genes of the 47-gene core enrichment subset (SLC40A1, TGFB3, SNRPN, ITGB6, BAMBI, TMEM176A, PDGFB and TMEM150A) were selected for validation. Briefly, each gene was individually knocked-out in SUM159PT using CRISPR/Cas9 before being orthotopically transplanted in the mammary fat pad of NSG mice, as previously described [10, 17]. Gene modification efficiency was assessed using a SURVEYOR assay from a bulk population of cells, confirming the indel mutations for each KO (Fig. 2a). Once tumors became palpable, daily intraperitoneal injections of the vehicle or 30 mg/kg palbociclib were each administered to five mice within each group, where each group consisted of 10–12 mice per gene knockout. As expected, tumor growth in non-targeting (NT) control mice groups was significantly inhibited by palbociclib by study endpoint (Fig. 2b, c). We found that individual knockout of our target genes effectively made cells more resistant to palbociclib over time (Fig. 2b). By study endpoint, all eight of the eight individual KOs (SLC40A1g1, TGFB3g1, ITGB6g3, BAMBIg2, TMEM176Ag3, PDGFBg1 and TMEM150Ag2) significantly inhibited the palbociclib anti-tumor effect in vivo, defining these genes as key regulators of TNBC response to palbociclib (Fig. 2c).
Fig. 2
In vivo validation of top candidate genes. a Gene modification detection of individual CRISPR-mediated knockouts of top candidate genes. b Cells transduced with non-targeting (NT) control or top candidate gene (SLC40A1, TGFB3, SNRPN, ITGB6, BAMBI, TMEM176A or PDGFB, TMEM150A) KO constructs were transplanted orthotopically into the mammary fat pads of NSG mice. Tumors were palpable before mice from each NT (n = 10–22) or targeting group (n = 10–12) were randomized into treatment groups (vehicle, n = 5–11; palbociclib (30 mg/kg), n = 5–11). Mean ± SD tumor volume is shown. Significance was calculated using two-sided, unpaired t-test, p-value ns. = nonsignificant, * < 0.05. c Tumor volumes of individual mice in each group, NT or targeting a candidate gene, either treated with vehicle or palbociclib at experiment endpoint (n = 5). Midlines indicate median tumor volume. Significance was calculated using two-sided, unpaired t-test, p-value * < 0.05
Having found that the depletion of our top targets generated resistance to palbociclib, we further explored the clinical translatability of our genes to predict the sensitivity of mammary tumors to CDK4/6 inhibitors. Accordingly, we used patient data from the NeoPalAna clinical trial, a single-arm phase II clinical trial evaluating the neoadjuvant use of palbociclib, with an anastrozole backbone, in clinical stage 2 or 3 ER + primary breast cancer [28]. Upon starting the trial, eligible patients received the aromatase inhibitor anastrozole (1 mg daily) for 28 days (Cycle 0). Palbociclib (125 mg daily on days 1–21, Cycle 1) was then added to the treatment regimen on day 1 of cycle 1 (C1D1). Tumor biopsies were collected on C1D1 and 14 days after the start of palbociclib treatment (C1D15). Although all patients were ER + , the only clinical subtype of breast cancer assumed to be responsive to palbociclib, the response to treatment varied in these patients. This illustrates the inadequacy of relying solely on the predictive power of ER positivity. We therefore posited that varying the expression levels of other genes, such as genes from our shortlist, might better predict these varying responses to palbociclib. Gene expression data from total RNA were generated using an Agilent microarray platform during the trial. Here, we compared data from palbociclib-sensitive patients with data from patients deemed palbociclib-resistant at C1D15 because of an inability to achieve complete cell cycle arrest (Ki67 > 2.7%). At C1D1, analysis of gene expression levels revealed lower levels of SLC40A1 and TGFB3 in resistant versus sensitive patients (Suppl Fig. 2a). This trend of lower SLC40A1 and TGFB3 expression in resistant versus sensitive patients was also observed at C1D15. Some of the remaining genes showed similar trends at both time points, but the overall statistical analysis was difficult to perform given that there were too few patients for whom we had gene expression data in the ‘palbociclib-resistant’ group. These data should therefore be interpreted with caution. Nonetheless, we propose that the trends observed in the expression of the top two genes, SLC40A1 and TGFB3, hint at the potential clinical relevance of our CRISPR screening results in Rb-proficient TNBC in patients with varying Rb statuses in ER + patients.
Analysis of publicly available clinical data on KM Plotter revealed that many of these genes were also correlated with relapse-free survival (RFS) across all breast cancer subtypes [29]. Lower gene expression of SLC40A1, TGFB3, SNRPN, TMEM176A and TMEM150A was significantly correlated (p < 0.05) with lower RFS (Suppl. Figure 2b). This may suggest that lower expression of these genes not only affects the response to palbociclib treatment but is also indicative of a worse overall prognosis for breast cancer patients.
Altogether, these results highlight the robustness of both the prioritization and the screening design used in our study. Furthermore, our in vivo findings may attest to the translatability of these results towards clinical applications, as we found that patients who were resistant to palbociclib did have lower median expression of SLC40A1 and TGFB3 in the NeoPalAna trial.
The high ranking obtained by TGFB3 in the prioritization scheme, the strong negation of the palbociclib effect by TGFB3 knockout in vivo, along with the inverse relationship observed between TGFB3 expression and palbociclib resistance in patients led us to further explore the potential value of TGFβ3 as a sensitizer to the palbociclib response. We hypothesized that the effect of palbociclib would be potentiated in TGFB3-overexpressing tumors, resulting in a greater growth reduction than in control tumors. Therefore, we applied a gain-of-function approach through activation of the TGFB3 endogenous gene promoter using the CRISPR/dCas9 Synergistic Activation Mediator (SAM) system, as previously described [10, 30]. As shown in Fig. 3a, we strongly induced TGFB3 gene expression in SUM159PT cells using three different sgRNAs targeting the TGFB3 gene promoter, without affecting TGFB1 or TGFB2 expression. TGFB3g2 SAM-infected SUM159PT cells were transplanted into the mammary fat pads of NSG mice.
Fig. 3
TGFβ3 potentiates palbociclib anti-tumor effect in vivo. a mRNA expression levels of TGFB1, TGFB2 and TGFB3 in SUM159PT following TGFB3-specific overexpression using CRISPR activation (CRISPR/dCas9 SAM) (n = 3). Data are represented as mean ± standard deviation (SD). Significance was calculated using two-sided, unpaired t-test, p-value * < 0.05. b Mice from control (lentiSAMv2) or TGFB3-overexpressing (TGFB3g2 SAM) groups (n = 13) were each randomized into treatment groups (vehicle, n = 6; palbociclib, n = 7). I.p. injections of the vehicle treatment or a low dose of palbociclib (10 mg/kg) were administered until study endpoint. Data are represented as mean ± SD. c Reduction in tumor growth presented for each group treated with palbociclib, lentiSAMv2 or TGFB3g2 SAM, as compared to the same groups treated with the vehicle. Data are represented as mean, at each timepoint. d left Tumor volumes of individual mice in each group at study endpoint. right Tumor weights of individual mice in each group at study endpoint. Midlines at median. Significance was calculated using ordinary, one-way ANOVA with Tukey’s multiple comparisons test, p-value * < 0.05, ** < 0.01, *** < 0.001. e Average mRNA expression levels of TGFB3 in tumors derived from the vehicle-treated control mice (n = 6) and the TGFB3-overexpressing mice (n = 6). Data are represented as mean ± SD. Significance was calculated using two-sided, unpaired t-test, p-value * < 0.05, ** < 0.01, *** < 0.001. f Protein levels of TGFB3 (60 kDa) in tumors derived from the vehicle-treated control mice (n = 6) and the TGFB3-overexpressing mice (n = 6). g Spontaneous metastasis to the lungs was assessed. Lung nodules were counted and compared in lungs derived from the vehicle-treated control mice (n = 7) and the TGFB3-overexpressing mice (n = 6). Data represent metastatic nodule count per pair of lungs per mouse. Midlines at median. Significance was calculated using nonparametric Mann–Whitney U-test, p-value * < 0.05, ** < 0.01, *** < 0.001. h The effect of TGFB3 CRISPR-mediated knockout on lung colonization was assessed. Data represent metastatic nodule count per pair of lungs per mouse. Midlines at median. i Schematic representation of the use of recTGFβ3 in combination with palbociclib. MDA-MB-231 TNBC cells were transplanted into the mammary fat pads of NSG mice. Tumors were palpable before mice were randomized into treatment groups: vehicle, n = 9; recTGFβ3, n = 8; palbociclib, n = 8, combo (recTGFβ3 + palbociclib), n = 9. j Average tumor volume was measured over time. Data are represented as mean ± SD. k Tumor volumes of individual mice in each group at study endpoint. Midlines at median. Significance was calculated using ordinary, one-way ANOVA with Tukey’s multiple comparisons test, p-value * < 0.05. l Quantification of Ki67-positive cells stained by immunohistochemistry in tumor tissues from all four groups. Data are represented as mean ± SD (n = 3–4). Significance was calculated using two-sided, unpaired t-test, p-value * < 0.05. m Representative images of Ki67 staining in two tumors per group
Tumors were grown until palpable and treated daily with a relatively low dose of palbociclib (10 mg/kg, i.p.) or vehicle up to 33 days post-implantation. Here, low-dose palbociclib was used to allow for the observation of a potential synergy between treatment and high TGFβ3 levels. As shown in Fig. 3b, low-dose (10 mg/kg) palbociclib treatment significantly reduced tumor growth in the lentiSAMv2 control tumors. A similar level of effect was observed when TGFB3 expression was induced in untreated cells (TGFB3g2 SAM vehicle). However, of the mice treated with palbociclib, those with TGFB3-overexpressing tumors had significantly lower average tumor growth rates than the control mice (Fig. 3b). Statistical significance of the difference in tumor volume was measured at all timepoints and is provided in Suppl. Figure 3a. This is reflected in the mean palbociclib-mediated tumor growth inhibition in each group of mice at every timepoint investigated, where the palbociclib effect on tumor growth inhibition is significantly greater in TGFB3-overexpressing tumors as compared to control mice during the entire experiment (Fig. 3c). This is indicative of a potentiation of the palbociclib effect by TGFβ3. At the study endpoint, palbociclib treatment combined with increased TGFB3 expression greatly reduced tumor volume compared to that in control mice (lentiSAMv2) treated with palbociclib (Fig. 3d, left panel). Tumors were weighed upon resection, and the results shown in Fig. 3d (right panel) indicate that the anti-tumor effects of palbociclib were also greatly enhanced when TGFB3 was overexpressed. To verify that the enhanced anti-tumor effect observed in the TGFB3 SAM tumors was attributable to a sustained increase in TGFB3 levels, TGFB3 levels were assessed in excised tumors. TGFB3 SAM tumors exhibited higher levels of TGFB3 at both the mRNA level and the protein level than the control tumors (Fig. 3e, f). Taken together, these results suggest that an increase in TGFB3 expression activates a synthetic lethal interaction upon CDK4/6 inhibition, allowing for greater growth inhibition.
Having thus far only evaluated TGFβ3’s contribution to tumor suppression, we wanted to address the other, pro-metastatic arm of the TGFβ family’s dual role in cancer – a concern due to frequent extrapolation of data relating to TGFβ1’s role in promoting breast cancer to TGFβ3 [31]. The role of TGFβ in providing breast cancer cells with metastatic capabilities – such as inducing epithelial-to-mesenchymal transition and priming cells for extravasation, has been well established for TGFβ1 [32, 33]. However, the TGFβ3 ligand specifically has not been well studied. Thus, we evaluated the effect of TGFB3 overexpression on the spontaneous metastasis of orthotopically transplanted breast cancer cells to the lungs using the CRISPR/dCas9 SAM system described above. Lung nodules were counted after euthanizing the transplanted mice. Mice overexpressing TGFB3 showed significantly fewer nodules on average than non-targeting control mice (Fig. 3g). In a follow-up experiment, we assessed the effect of TGFB3 gene silencing on lung colonization. TGFB3 KO SUM159PT cells were injected into the tail veins of NSG mice, and lung nodules were counted 38 days after cell injection. We observed a trend towards an increased number of nodules in TGFB3 KO mice compared to non-targeting control mice (Fig. 3h). Taken together, these data suggest that inducing TGFB3 gene expression does not adversely affect lung metastasis in vivo, while leading to an increased sensitivity of tumors to palbociclib treatment in vivo. This highlights a possible therapeutic avenue for the administration of exogenous TGFβ3.
Therefore, we exploited the inherent ease of use of TGFβ3 as a potential treatment, being a naturally occurring ligand. Human recombinant TGFβ3 (recTGFβ3) has previously been developed into an intradermal injectable (avotermin) and has been safely used in phase II and III clinical trials for the prevention of scarring [34]. To validate our findings in another TNBC model and thereby broaden the scope of the implications of our findings, we assessed recTGFβ3/palbociclib anti-tumorigenic effects when administered alone or in combination in preformed MDA-MB-231-derived mammary tumors. MDA-MB-231 is a poorly differentiated, aggressive TNBC cell line derived from the pleural effusion of a 51-year-old Caucasian female [35]. These cells were transplanted into the mammary fat pads of NSG mice, which were then randomized into four groups. Either the vehicle, human recTGFβ3 alone (2 µg/kg), palbociclib alone (10 mg/kg), or a combination of recTGFβ3 (2 µg/kg) and palbociclib (10 mg/kg) was administered intraperitoneally to mice in each group (Fig. 3i). Treatment was initiated 33 days after transplantation, once the tumors were palpable and administered daily. The smallest average tumor volume was observed in the combination group (Fig. 3j). By the endpoint, mice from the groups treated with suboptimal doses of either recTGFβ3 alone or palbociclib alone showed comparable tumor volumes to mice in the control group, whereas the recTGFβ3 + palbociclib combination group had significantly smaller tumors than the control group (Fig. 3k, Suppl. Figure 3b). Moreover, analysis of the proliferation index (Ki67) by immunohistochemistry in these tumors revealed that the combination treatment significantly reduced the proportion of proliferating cells as compared to the vehicle (Fig. 3l, m). This is reflective of tumor volume at endpoint, as neither palbociclib alone nor recTGFβ3 alone significantly reduced cell proliferation in vivo, indicating a potential synergy between the two treatments when administered together. These findings highlight the clinical relevance of TGFβ3 as a synthetic lethal target in our screen for its role in potentiating the anti-tumor effects of palbociclib when administered as a recombinant protein. They indicate the ease with which TGFβ3 could be administered in the clinic in combination with palbociclib to achieve significant tumor growth inhibition using low doses of either treatment. This could potentially help avoid unwanted adverse effects of using high individual doses while allowing for on-target inhibition of tumor growth unachievable at low doses of palbociclib.
Having shown that both TGFB3 overexpression and the use of recTGFβ3 significantly promoted the palbociclib response in reducing tumor growth (Fig. 3), we sought to gain insight into the molecular mechanism by which these two drugs work together. To better understand the nature of the relationship between palbociclib and recTGFβ3, we assessed combinatorial synergy using drug matrix assays in multiple Rb + TNBC cell lines: SUM159PT, SUM229PE, and MDA-MB-231. To start to address this, dose–response analyses with TGFβ3 or palbociclib alone were performed in these TNBC cell lines. As shown in Suppl. Figure 4a, TGFβ3 stimulation of the cells only produced a modest effect that plateaued at approximately 20% growth inhibition. Palbociclib efficiently reduced cell viability within a given concentration range (Suppl. Figure 4b). Ultimately, dose ranges of palbociclib (12.5 nM to 400 nM) and recTGFβ3 (3.13 pM to 100 pM) were used alone or in combination and cell proliferation was assessed by crystal violet staining.
We used four reference synergy models to assess combinatorial effects in our study: Bliss, Highest Single Agent (HSA), Loewe, and Zero Interaction Potency (ZIP). Each of these models uses different formulas and assumptions to calculate drug combination synergy [36]. Interestingly, we found that for all cell lines tested, overall synergy was observed across the dose combinations tested, with scores greater than 10 indicating a strong likelihood of a synergistic relationship [36] (Fig. 4a). Notably, cotreatment attained a level of synergy that could be reproducibly obtained using all four models tested. The highest degrees of synergism tended to occur at the lower concentrations used for palbociclib, as denoted by the grey rectangles in each graph and the ‘Most synergistic area score’ (Fig. 4a, b). The percentages of treatment-induced proliferation inhibition for each pairwise comparison in the drug matrices presented help underscore the impact of the combination treatment in each cell line (Suppl. Figure 4d). This further highlights the clinical relevance of our findings, where submaximal doses of palbociclib could be administered, limiting the associated side effects and reducing the need for treatment cycle delays, along with TGFβ3, to achieve an even greater anti-proliferative effect than palbociclib alone. This is especially relevant in a context where cancer patients are subjected to many treatment-associated toxicities, both with palbociclib and radiotherapy or chemotherapy treatments [37].
Fig. 4
Combination of recombinant TGFβ3 and palbociclib synergistically inhibits TNBC cell proliferation in vitro. a Synergy between palbociclib and recTGFβ3 dose combinations was calculated based on four reference models (Bliss, HSA, Loewe, ZIP) using SynergyFinder in four TNBC cell lines (MDA-MB-231, SUM159PT, SUM229PE, 159-R). Synergy maps highlight areas of synergistic (red) or antagonistic (green) interactions between given concentrations of either agent. Grey boxes indicate the area of maximum synergy observed. Mean of a minimum of three independent replicate experiments for each cell line (n ≥ 3). b ‘Overall synergy scores’ and ‘Most synergistic area scores’ presented for each drug matrix shown in a. Data are represented as score ± 95% confidence interval. c Dot plots show overall synergy scores (black) or most synergistic area scores (pink) for each cell line, with each dot representing the score obtained using the indicated reference model. Midlines represent median scores. Outer vertical lines correspond to minimum and maximum scores obtained. A zero ‘0’ score indicates no interaction between the two agents
We then investigated whether recTGFβ3 could be used to resensitize cells to palbociclib in a model where cells had become resistant to palbociclib due to chronic exposure to the drug. To this end, we first generated a palbociclib-resistant SUM159PT cell line (159-R) by treating SUM159PT with gradually increasing concentrations of palbociclib over four months. A dose–response curve evaluating palbociclib response in 159-R was used to confirm palbociclib resistance (Suppl. Figure 4c). We performed drug matrix assays using palbociclib concentrations ranging from 78 nM to 2.5 µM, while TGFβ3 concentrations ranged from 3.13 pM to 100 pM. Although higher concentrations of palbociclib were necessary in 159-R to generate a similar level of response to the low doses of palbociclib used in parental SUM159PT, we chose to keep the same range of recTGFβ3 concentrations to determine whether resistant cells could be resensitized to palbociclib at the same low concentrations. We found that not only could resistant cells be resensitized to palbociclib by cotreatment with recTGFβ3, but that TGFβ3 could synergize with the effects of palbociclib. Indeed, in 159-R, overall synergy was achieved for the drug concentration ranges tested using all four algorithms (Fig. 4a, b, Suppl. Figure 4d). As demonstrated in Fig. 4c, the robustness of this interaction is made evident by the high synergy scores obtained in all cell lines, regardless of previous exposure to palbociclib, and across all algorithms for the ‘Overall synergy scores’ (black) as well as ‘Most synergistic area scores’ (pink). The potential noninteractive zone (dotted line) was excluded from the range of scores obtained for every synergy score analysis (Fig. 4c). The synergy demonstrated in the treatment-naïve context helps to characterize the interplay observed in the in vivo study, demonstrating that the combination of recTGFβ3 + palbociclib treatment leads to the greatest tumor growth inhibition. Most importantly, this synergy is still achieved when cells are desensitized to palbociclib through chronic exposure to the drug.
To understand the molecular mechanisms underlying the synergism between palbociclib and TGFβ3 growth inhibitory effects in TNBC, we examined the effects of palbociclib on the expression levels of cell cycle regulators. Palbociclib treatment of SUM159PT cells over 24 h led to significant time-dependent increases in established resistance markers, such as CDK4, cyclin D1 and cyclin E1, along with concomitant decreases in Rb and phospho-Rb (Ser780) (Fig. 5a). The various times at which these changes in protein levels occurred may reflect the indirect nature of these changes in protein levels. Of note, observable and significant changes in phosphorylation of Rb occurred earlier in the time course, whereas a significant decrease in Rb levels was observed after 24 h only (Fig. 5a). We observed no consistent changes in CDK6 nor the CDK inhibitor CDKN1B (p27) over 24 h. For CDKN2A (p16) and CDKN1C (p57), we found there was no detectable signal. However, there were changes in protein levels of the other phases of the cell cycle, especially later in the time course (Suppl. Figure 5a). Accordingly, these decreases in CDK1, cyclin A1, cyclin B1, and PLK1 were in line with the decrease in proportion of cells which proceeded to S-phase and continued cycling through the cell cycle after addition of palbociclib (Suppl. Figure 5b). Indeed, following cell cycle analysis by flow cytometry, it is clear that treatment with palbociclib arrests cells in G1, but that the induction of G1 arrest is strongest and significant upon the addition of recTGFβ3, which also entails a significant decrease in the proportion of cells in S-phase (Suppl. Figure 5b).
Fig. 5
TGFβ3 synergizes with palbociclib in a p21-dependent way. a SUM159PT cells were treated with palbociclib (100 nM) for 2 h, 8 h, 16 h and 24 h and protein lysates were assessed for known CDK4/6i resistance markers (CDK4, cyclin D1, cyclin E1, Rb, phospho-Rb (S780)) by immunoblotting. Relative fold changes in protein levels, compared to untreated cells at each timepoint, were calculated (n = 3). Data are represented as mean ± SD. Significance was calculated using two-sided, unpaired t-test, p-value * < 0.05, ** < 0.01, *** < 0.001. b SUM159PT (159) and 159-R cells were assessed for known CDK4/6i resistance markers, as well as p21, by immunoblotting. c top SUM159PT and 159-R cells were treated with recTGFβ3 (100 pM) for 24 h and resulting changes in known CDK4/6i resistance markers and p21 were measured by immunoblotting. bottom MDA-MB-231 (231) and SUM229PE (229) cells were treated with recTGFβ3 (200 pM) for 24-48 h and resulting changes in p21 were measured by immunoblotting. d SUM159PT cells were transduced with plasmids encoding control (scramble, scr), Smad2-specific, or Smad3-specific short hairpin RNAs (shRNA). Protein levels of p21 and total Smad2/3 were measured by immunoblotting. e SUM159PT cells were transduced with plasmids encoding control (scr) and p21-specific shRNA. Protein levels of p21 were measured by immunoblotting. f SUM159PT scr shRNA-infected or p21 shRNA-infected cells were treated with varying combinations of palbociclib and recTGFβ3 concentrations. Synergy between dose combinations was calculated using SynergyFinder. upper Synergy maps highlight areas of synergistic (red) or antagonistic (green) interactions between given concentrations of either agent. Grey boxes indicate the area of maximum synergy observed between given recTGFβ3 and palbociclib dose combinations. lower ‘Overall synergy scores’ and ‘Most synergistic area scores’ presented for each drug matrix shown above. Data are represented as score ± 95% confidence interval (n = 3). Percentage variation in synergy score (score obtained in p21 shRNA cells/score obtained in scr shRNA cells) is also shown (red)
To next determine whether these changes in cell cycle marker expression would be transposed in the long-term palbociclib acquired resistance context, we compared their levels in naïve and resistant cells that had undergone chronic exposure to the drug, in SUM159PT and 159-R, respectively. As shown in Fig. 5b, strong increases in CDK4, cyclin D1, and cyclin E1, along with a stark decrease in Rb and p-Rb expression, were observed in the resistant cells, indicating that the effects of chronic palbociclib exposure mimicked the changes in marker levels observed in the short-term acquired context. We also found that palbociclib decreased the expression of the cell cycle inhibitor CDKN1A (p21). This defines p21 as a palbociclib target and is consistent with decreased palbociclib efficacy and short-term acquired resistance.
The TGFβ family of ligands acts as potent tumor suppressors notably by inducing CDK inhibitors (CDKIs) [38]. Thus, we examined whether TGFβ3 could modulate the expression of the CDK inhibitor p21 in both parental and palbociclib-resistant SUM159 cells. As shown in Fig. 5c, TGFβ3 strongly induced p21 expression in multiple TNBC cell lines, as demonstrated in SUM159PT, MDA-MB-231 and SUM229PE. Furthermore, it restored p21 levels in palbociclib-resistant cells, suggesting that TGFβ3-mediated p21 expression induction contributes to the synergism observed between palbociclib and recTGFβ3. This is also exhibited at the mRNA level, where treatment with recTGFβ3 significantly induces p21 levels in SUM159PT and, to an even greater extent, in 159-R (Suppl. Figure 5c). At the basal level, without recTGFβ3 treatment, there is a significant decrease in p21 in cells chronically exposed to palbociclib, 159-R, at the mRNA level (Suppl. Figure 5c). This is reflected at the protein level as well (Fig. 5c). Therefore, we further addressed the specific role and contribution of p21 in mediating these effects. First, we determined that the effect of p21 upregulation by TGFβ3 was Smad2/3-dependent. When Smad2 and Smad3 were knocked down individually in SUM159 cells, the TGFβ3-mediated increase in p21 level was diminished (Fig. 5d). Given that Smad2/3 induction of p21 occurs through the well-established canonical Smad signaling pathway shared by all TGFβ isoforms, we asked whether the synergy observed between TGFβ3 and palbociclib could also be observed between TGFβ1 and TGFβ2 with palbociclib. We tested whether these isoforms could confer similar synergistic effects on palbociclib efficacy in MDA-MB-231 and SUM159PT cell lines and found that all three TGFβ isoforms demonstrate a similar effect on palbociclib efficacy (Suppl. Figure 5d). This is in line
留言 (0)