
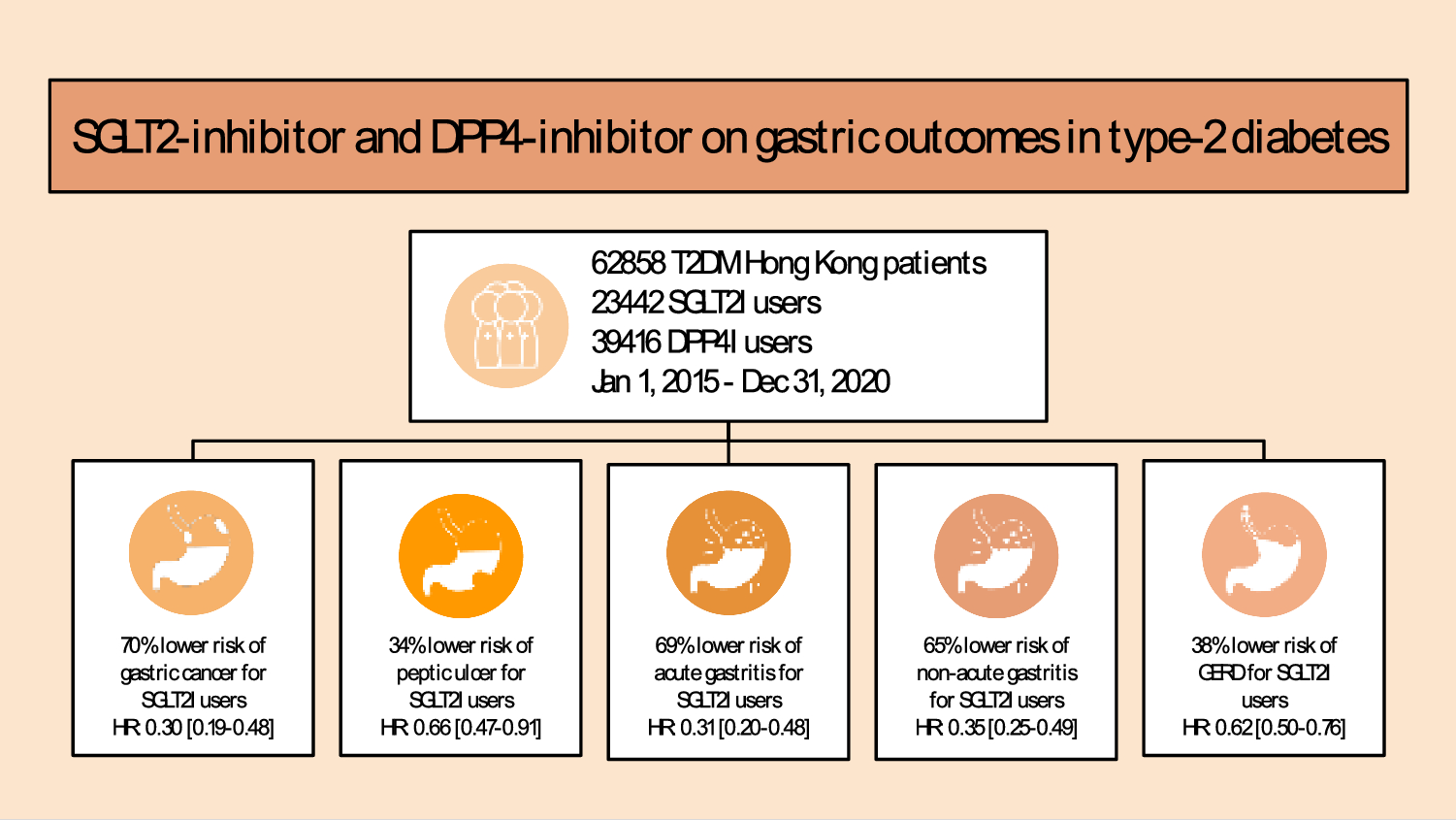
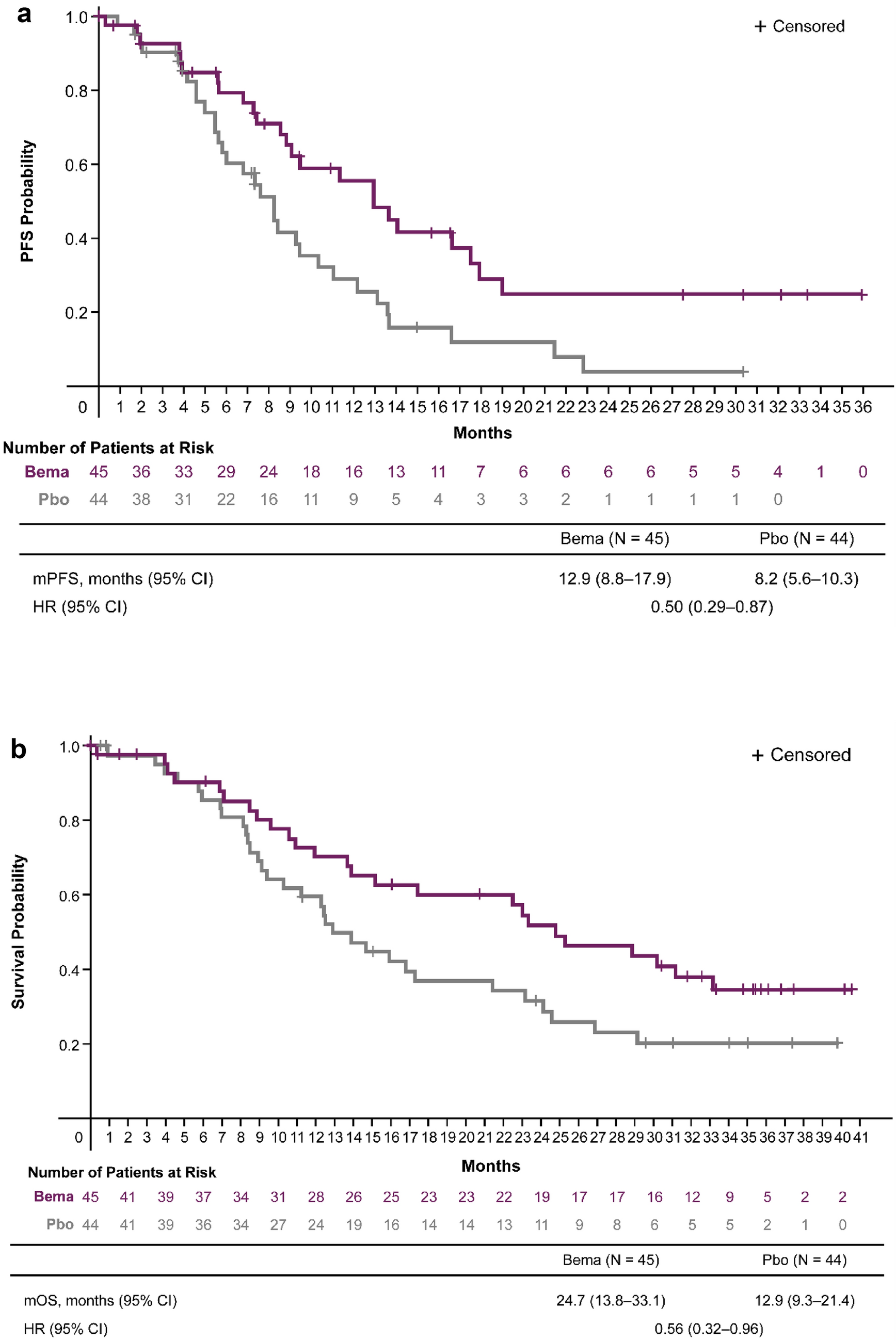
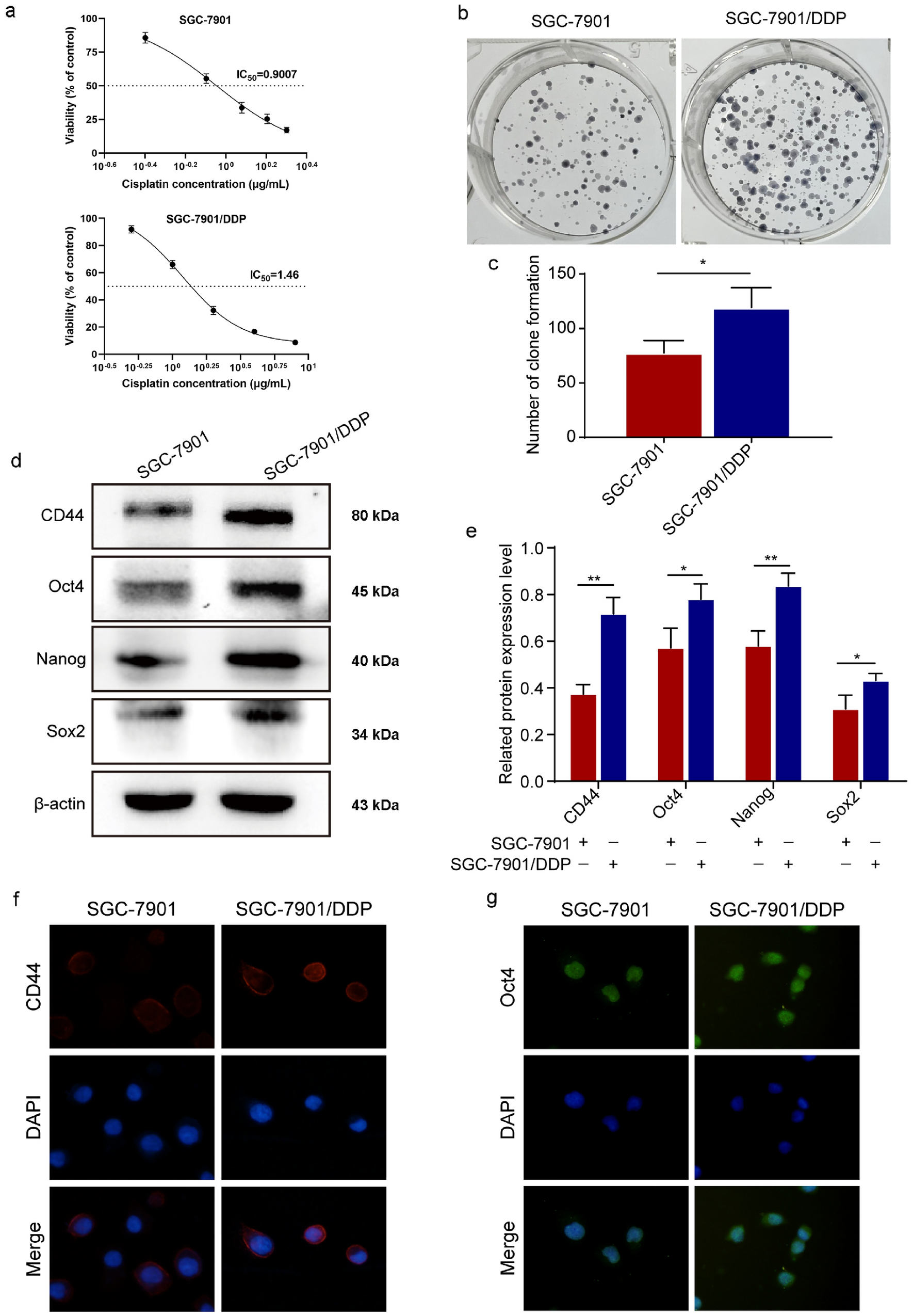
記住我



To evaluate the efficacy of ICI in the context of peritoneal dissemination of GC, we established a female C57BL/6 J GC peritoneal dissemination mouse model [13, 14]. The grouping and treatment timeline are depicted (Fig. 1A, supplementary Fig. 1A). Following the implantation of YTN16 cells on day 0, IP administration of ICI or PBS commenced on days 5, 8, 12, 21, and 28. Concurrently, T-cell depletion was achieved via IP injection of anti-CD4 or anti-CD8 mAb on days 4, 7, and 11 for confirming the therapeutic effect originated from T cell. On day 35, samples were collected for analysis. Beginning in the fourth week following YTN16 cell transplantation, mice displayed a declining trend in weight, excluding those in the dual ICI treatment group (supplementary Fig. 1B). One mouse out of 5 mice treated with anti-PD1 monotherapy was cured, but none out of 5 mice treated with anti-CTLA4 monotherapy was cured. In the anti-CTLA4 group, one mouse, which died before the autopsy, was excluded from the analysis (Supplementary Fig. 1C). Remarkably, among the five mice subjected to dual ICI treatment, three exhibited significant improvements in macroscopic disseminated nodules compared to the monotherapy group (Fig. 1B, Supplementary Fig. 1D). The treatment efficiency was abrogated upon introducing CD4 + or CD8 + T cell depletion in conjunction with dual ICI treatment. In the dual ICI + anti-CD8 group, two mice, which died before the autopsy, were excluded from the analysis. In these mice, a notable reversal in therapeutic outcomes was observed, marked by an exacerbation in peritoneal tumor dissemination and even pronounced adhesions and jaundice (Fig. 1B, Supplementary Fig. 1E). Immune infiltrating cells were quantified based on bulk RNA-seq data using mMCP-counter analysis (Fig. 1C). Because peritoneal dissemination had been entirely eradicated in three mice from the dual ICI treatment group by the fifth week, distinct infiltration patterns on the heatmap suggested a state of “post-war” quiescent immune response (Fig. 1C, mice #2, #3, and #4). However, even among the uncured cases, mice from the dual ICI treatment group (Fig. 1C, mice #1, #5) displayed significantly higher levels of immune-infiltrating CD8 + T cells compared to the mice in the other groups. In addition, the volcano plot illustrated the differentially expressed genes (DEGs) between the dual ICI and untreated group, with upregulated DEGs marked with red solid dots. The notable upregulation of gene expression in Cd8a, Cd8b1, H2-Eb2, H2-Q2, and Ly6c2 indicated enhanced immune activation and potential involvement of MHC-related pathways, as well as an association with inflammation-related processes in dual ICI-treated mice (Fig. 1D).
Fig. 1
Dual ICI treatment outperformed monotherapy. CD4 + and CD8 + T cells were both indispensable for the anti-tumor effect. A Treatment timeline in female C57BL/6 J GC peritoneal dissemination model. B Representative macroscopic images of peritoneal dissemination tumor of mice on day 35. Three out of five mice demonstrated improved efficacy with dual ICI treatment compared to the monotherapy group. However, the clearance of CD4 + or CD8 + T cells, in conjunction with dual ICI treatment, resulted in exacerbated peritoneal dissemination of the tumor. Adhesions of the peritoneum and liver could be seen in the CD8 + T cells depletion group (black arrow). C Quantification of immune-infiltrating cells based on bulk RNA-seq data using mMCP-counter analysis. The dual ICI group (mice #1 and #5) exhibited significantly higher levels of immune-infiltrating CD8 + T cells compared to other groups. By the fifth week, the peritoneal dissemination in mouse #2, #3, and #4 had been completely cured. Consequently, the observed distinct infiltration pattern in the heatmap suggested a quiescent immune response, indicating a cessation of immune activation. D The volcano plot illustrated the DEGs between the dual ICI and untreated groups. The x-axis represented the log2 fold change, indicating the magnitude of gene expression change, while the y-axis represented the statistical significance. Red solid dots highlighted the upregulated DEGs in the dual ICI group, exceeding the significance thresholds (p < 0.05 and fold change > 2). E Representative images of IHC staining of CD8 + T cells on day 35. F Enhanced infiltration of CD8 + T cells was observed in the dual ICI treatment group, particularly with CD4 + T cell depletion, resulting in a significant increase compared to other groups. G In the dual ICI + anti-CD4 treatment group, IHC staining revealed that although there was significant CD8 + T cell infiltration, only a small proportion exhibited GZMB + cytotoxic phenotype, while the majority displayed an EOMES + exhausted phenotype. Scale bar: 50 μm; inset: magnified view of the boxed area. *p < 0.05, one-way ANOVA with Tukey's multiple comparisons test
The presence of CD8 + T cell infiltration was verified via immunohistochemical (IHC) staining (Fig. 1E, F). Remarkably, in the uncured mice from the dual ICI treatment group, there was a substantial increase in CD8 + T cell infiltration compared to the monotherapy and untreated groups. Furthermore, an intriguing observation arose when combining dual ICI treatment with CD4 + T cell depletion. Specifically, though substantial CD8 + T cell infiltration was evident, the majority exhibited an EOMES + exhausted phenotype, while only a minority displayed the GZMB + cytotoxic phenotype (Fig. 1G). In these unremitting tumors, a substantial infiltration of CD68 + macrophages, Ly6G + neutrophils, and Foxp3 + regulatory T cells (Tregs) were observed, indicating the formation of an immunosuppressive TME. It is worth noting that, despite a significant reduction in Foxp3 + Tregs infiltration following anti-CD4 and anti-CD8 treatment, the tumor did not show any improvement (supplementary Fig. 2).
Dual ICI treatment exerted anti-tumor immune effects by enhancing the infiltration of tumor neoantigen-specific CD8 + T cellsTo observe treatment affected tumor microenvironment at an earlier time point, before tumor disappearing for the dual ICI therapy, we euthanized the mice at the third week (Fig. 2A). The study involved IP implantation of YTN16 cells on day 0, followed by IP treatments with ICI or PBS on days 5, 8, and 12. After 21 days, mice were euthanized, and peritoneal tumors were collected for RNA-seq, fluorescence activated cell sorting (FACS), and IHC analysis. In addition, peritoneal lavage fluid was examined for FACS analysis of peritoneal exudate cells.
Fig. 2
Dual ICI treatment exerted anti-tumor immune effects by enhancing the infiltration of tumor neoantigen-specific CD8 + T cells. A Treatment timeline in female C57BL/6 J GC peritoneal dissemination model. B The quantification of immune cell infiltration during the anti-tumor immune response was conducted at both the early stage (3wk) and the late stage (5wk, uncured mice) using mMCP-counter analysis. Dual ICI treatment resulted in increased CD8 + T cell infiltration compared to the untreated group at both time points. Notably, at the early time point (3wk), the dual ICI group exhibited relatively fewer neutrophil infiltrations compared to the untreated group. However, in the late stage of dual ICI treatment, increased neutrophil infiltration was observed in uncured tumors. C FACS analysis was performed on peritoneal tumor and peritoneal exudate cells at the early time point (3wk). Significant increase observed in tumor neoantigen-specific CD8 + T cells in the dual ICI treatment group. Tumor neoantigen-specific CD8 + T cells were defined as composed of mCdt1-H-2 Kb-dimer + CD8 + T cells, mZfp106-H-2Db-dimer + CD8 + T cells, and mScarb2-H-2Db-dimer + CD8 + T cells. *p < 0.05, Kruskal–Wallis test with Dunn’s multiple comparison test. D Representative images of peritoneal dissemination of mice on day 21. E IHC staining of CD8 and GZMB showed enhanced infiltration of GZMB + cytotoxic T cells into the tumor following dual ICI treatment at the early time point (3wk). Scale bar: 50 μm; Inset: a magnified view within the red dashed line, presented in a black frame on the right
The immune cell composition at the third week was depicted in a pie chart format using CIBERSORT. The dual ICI treatment group exhibited an increased relative proportion of CD8 + T cells, whereas in the untreated group, neutrophils accounted for a higher proportion (supplementary Fig. 3). Similarly, at this earlier time point (3wk), we also conducted mMCP-counter analysis and compared the results with those from the later time point (5wk). Dual ICI treatment resulted in increased CD8 + T cell infiltration compared to the untreated group at both time points. Notably, at the earlier time point (3wk), the dual ICI group exhibited relatively fewer neutrophil infiltrations compared to the untreated group. However, in the late stage of dual ICI treatment (5wk), increased neutrophil infiltration was observed in uncured tumors (Fig. 2B). This shift in neutrophil infiltration within tumors was also confirmed by IHC for ly6G, a neutrophil marker (supplementary Fig. 4A).
Previously, we have identified 3 neo-antigens for YTN16 cells, including mCdt1, mZfp106, and mScarb2 peptides [15]. Flow cytometry analysis revealed increased CD8 + T cell in tumor-bearing peritoneum and peritoneal lavage fluid with dual ICI treatment, along with a significant rise in neo-antigen-specific CD8 + T cells at the earlier time point (Fig. 2C). Notably, there were no significant differences observed in the levels of NK cells, B cells, and CD4 + T cells among groups (Supplementary Fig. 4B).
Representative images showed similar peritoneal tumor sizes among mice in the untreated and dual ICI group (Fig. 2D). IHC analysis revealed an increase in CD8 + T cell infiltration in the tumor area following dual ICI treatment, compared to the untreated group. Remarkably, most of these CD8 + T cells exhibited a GZMB + cytotoxic T cell phenotype, while the limited number of CD8 + T cells present in the untreated group did not show GZMB expression (Fig. 2E).
The above analysis indicated that dual ICI treatment exerted potent anti-tumor effects by enhancing the infiltration of tumor neoantigen-specific CD8 + T cells. Notably, neutrophil infiltration displayed varying patterns, with reduced presence at the early stage and increased infiltration in uncured tumors during late-stage treatment. It is conceivable that during the treatment process of dual ICI, the tumor response gradually shows different trends. Some mice may manifest a sustained response to ICI, resulting in a gradual tumor clearance, whereas others, despite the activation of an immune response, may develop resistance to ICI for unspecified reasons, subsequently leading to further tumor progression.
Excessive activation of JAK-STAT pathway in dual ICI-resistant tumorsTo investigate the underlying causes of resistance in dual ICI treatment, we conducted an examination of immune checkpoints, including both inhibitory and co-stimulatory molecules, along with the assessment of relevant cytokine and cytotoxic molecule expression. Notably, following dual ICI treatment, several immunosuppressive checkpoint genes showed significant upregulation, such as Pdcd1, Ctla4, Lag3, Tight, Tim, Lgals9, Btla, and Tnfrsf14. This upregulation was observed in some mice during the early stage and persisted in uncured mice during the late stage following dual ICI treatment. In contrast, immune-positive stimulatory signals such as CD28, were expressed during the early stage (3wk) but markedly declined in the late stage (Fig. 3A). In response to dual ICI treatment, the expression levels of interleukin (IL)-1, 4, 6, and 13 increased indicating an elevated inflammatory response and potential tumor-promoting involvement. Notably, IL-12, crucial for T cell activation and essential for anti-tumor immune responses, increased after dual ICI treatment but declined in late-stage (5wk) ICI-resistant mice. Granzyme B (Gzmb), Perforin-1 (Prf1), Tumor Necrosis Factor (Tnf), and Interferon Gamma (Ifng) also exhibited noticeable upregulation following dual ICI treatment. However, a declining trend in Ifng expression was observed in the late-stage (5wk) ICI-resistant mice, concomitant with an increase in Transforming Growth Factor Beta-1 (Tgfb1) associated with tumor epithelial–mesenchymal transition (EMT) (Fig. 3B). In addition, we generated a heatmap to assess changes in chemokine expression following dual ICI treatment, revealing significant upregulation of multiple chemokines, including Ccl2,17,20,28, and Cxcl 1,2,3,5,11,16. These chemokines, crucial for regulating immune cell chemotaxis and inflammatory processes, highlight the potential impact of dual ICI treatment on modulating the intricate network of chemotactic signaling. Such changes may facilitate the recruitment and activation of immune cell populations, ultimately influencing the immune landscape within the TME (Supplementary Fig. 5).
Fig. 3
Enhanced activation of the JAK-STAT pathway observed in dual ICI-uncured mice compared to untreated mice. A, B Heatmap illustrating the expression of immune checkpoint molecules and cytokines, cytotoxic molecules at both early (3wk) and late (5wk) time points in untreated and dual ICI-uncured mice. C GSEA of MSigDB hallmark gene sets in dual ICI-uncured mice compared to untreated Mice. The top 10 enriched gene sets in dual ICI-uncured mice were displayed. D Enrichment analysis of KEGG pathways was performed using STRING. Pathways enriched in dual ICI-uncured mice were displayed in a bubble chart
We employed gene set enrichment analysis (GSEA) to analyze the top ten enriched Molecular Signatures Database (MSigDB) hallmark gene sets with corresponding enrichment scores in dual ICI-uncured group compared to the untreated group (Fig. 3C). Following the dual ICI treatment, pathways related to immune response, such as INTERFERON_GAMMA_RESPONSE and TNFA_SIGNALING, showed discernible upregulation. Particularly noteworthy was the activation of the IL6_JAK_STAT3_SIGNALING pathway, a key driver of pro-inflammatory signaling. This pathway plays a pivotal role in diverse cellular processes such as cell proliferation, differentiation, and immune regulation. Furthermore, the INFLAMMATORY_RESPONSE pathway also showed substantial activation in the dual ICI-uncured group, further highlighting the complex interplay of inflammatory mechanisms (Supplementary Fig. 6A).
Enrichment analysis of Kyoto Encyclopedia of Genes and Genomes (KEGG) provided an encompassing perspective on the molecular processes influenced by the dual ICI treatment (Fig. 3D). The activation of the antigen processing and presentation pathway, T cell receptor signaling pathway and Th1 and Th2 cell differentiation pathways indicated the potential enhancement of antigen recognition by immune cells, heightened T cell activation and differentiation, which were critical for an effective immune response against tumors. Notably, KEGG analysis also revealed the activation of the JAK-STAT pathway.
The Gene Ontology (GO) enrichment analysis yielded a wide spectrum of enriched terms across Biological Process (BP), Cellular Component (CC), and Molecular Function (MF) categories in dual ICI-uncured mice compared to untreated mice (Supplementary Fig. 6B). BP terms included ‘Positive regulation of T cell activation’ and ‘Myeloid leukocyte activation’ suggested associations with immune modulation and regulation. The enriched CC terms, such as ‘Kinetochore’ and ‘Immunological synapse’, offered insights into potential cellular compartments and structures influenced by the therapeutic interventions. Within the MF category, terms like ‘Cytokine receptor activity’ and ‘Phosphoprotein binding’ implied involvement in signaling and interaction mechanisms. These findings collectively provided clues about how interventions may impact various immunological and molecular processes, contributing to our understanding of the complex interplay between immune responses and TME dynamics in the context of GC peritoneal dissemination.
JAK-STAT pathway was considered a potential therapeutic targetWe compared the gene profiles of the anti-PD1, anti-CTLA4 monotherapy group, the dual ICI-uncured group, and the untreated group with those of the dual ICI-cured mice. We screened for DEGs from each group and identified potential therapeutic targets by overlapping them with FDA-approved 216 cancer relative targets (obtained from Human Protein Atlas). This analysis was visually represented in a Venn diagram, illustrating a total of 58 potential therapeutic targets crucial to understanding the tumor’s progression (Fig. 4A). A protein–protein interaction (PPI) network diagram was constructed using these 58 target proteins. The network was filtered to retain only high-confidence interactions (0.7), while hiding disconnected nodes. In addition, Markov Cluster Algorithm (MCL) clustering was utilized to visualize the protein interaction clusters (Fig. 4B). Local network clustering analysis using the String platform revealed enrichments in key biological terms, such as ‘apoptosis,’ ‘NF-kappa B signaling,’ ‘TNF receptor superfamily,’ ‘JAK-STAT pathway,’ and ‘vascular endothelial growth factor signaling pathway.’ These findings shed light on potential therapeutic avenues (Supplementary Table 2). Consequently, whether it relates to the tumor’s initiation and progression or its resistance to ICI treatment, the activity of the JAK-STAT pathway assumes a profoundly pivotal role. This exploration suggested a promising therapeutic potential of JAK inhibitor.
Fig. 4
The JAK-STAT pathway was considered a potential therapeutic target. A A Venn diagram that is generated by overlapping the DEGs (from each group compared to the dual ICI-cured group) with the FDA-approved cancer-related targets. B A PPI network diagram was constructed using the overlapped 58 target proteins. The network was filtered to retain high-confidence interactions (0.7) and visualize protein interaction clusters using MCL clustering while hiding disconnected nodes
Ruxolitinib (JAK1/2 inhibitor) synergistically improved the immunosuppressive TME in mice with dual ICI resistanceOn the foundation of dual ICI application, we administered oral treatment with a JAK inhibitor Ruxolitinib in an attempt to ameliorate resistance to ICI treatment by mitigating excessive inflammatory responses following ICI-induced immune activation. Female C57BL/6 J mice, aged 5 weeks, were divided into four groups: untreated, JAKi, dual ICI, and dual ICI + JAKi.
Dual ICI treatment was initiated on days 5, 8, and 12 following YTN16 implantation. Starting from day 8, Ruxolitinib was administered orally at a daily dose of 30 mg/kg for 1 week. The mice were euthanized on day 21 (Fig. 5A). In the untreated and JAKi monotherapy groups, no improvement was observed, and in the dual ICI group, 3 out of 5 mice exhibited resistance. However, to our surprise, the group that received the combination therapy of dual ICI and JAKi achieved complete tumor clearance in 4 out of 5 mice, with only one mouse showing residual tumor in a small area (Fig. 5B red circle). After subsequent IHC staining, an abundance of GZMB + cytotoxic T cells was discovered within the tumor of this mouse, resembling that observed in the dual ICI group. Furthermore, minimal presence of M2 macrophages, neutrophils, and Tregs was noted. In addition, by day 21, following cessation of JAKi administration on day 15, the expression of p-STAT3 was similar across all groups (Fig. 5C).
Fig. 5
Ruxolitinib in combination with dual ICI treatment further enhanced the anti-tumor effect. A Treatment timeline in female C57BL/6 J GC peritoneal dissemination model. B Representative macroscopic images of peritoneal dissemination of mice on day 21. Red circle indicated the residual tumor site in the only mouse from the dual ICI + JAKi group. C Representative images of IHC staining of p-STAT3, cytotoxic T cells, M2 macrophages, neutrophils, and Tregs on day 21. The dual ICI + JAKi group displays the only uncured mouse with a local residual tumor in the peritoneum. The IHC staining of this mouse was characterized by an abundance of GZMB + cytotoxic T cell infiltration, accompanied by minimal presence of M2 macrophages, neutrophils and Tregs. Scale bar: 50 μm
To collect tumor tissue for analysis before complete clearance, we modified the treatment schedule. We administered dual ICI treatment on day 5, followed by a second dose on day 8. JAKi was administered from day 8 to 11, and the mice were euthanized 3 h after the last dose of JAKi on day 11 (Fig. 6A). Upon dissection, it was observed that the extent of peritoneal dissemination was consistent across all groups (Fig. 6B). Building on this observation, we conducted Western blot analysis to assess the impact of the JAK inhibitor on p-STAT3 protein expression. Following JAKi administration, we observed a significant decrease in the intensity of the p-STAT3 protein bands in comparison to the untreated group (Fig. 6C), suggesting effective suppression of JAK-STAT3 pathway activation. In addition, the IHC results for p-STAT3 suggested its suppression by JAKi (Fig. 7). The IHC results showed that in the treatment regimens involving dual ICI, there was a significant infiltration of GZMB + cytotoxic T cells, while in the regimens involving JAKi, the expression of CD206 + M2-type macrophages was relatively lower. Ly6G + neutrophils in groups that received either JAKi, the dual ICI treatment regimen, or a combination of both did not show a significant increase, unlike the untreated group. While dual ICI treatment showed a modest increase in Foxp3 + Tregs, although it did not reach statistical significance. The therapeutic effectiveness of dual ICI + JAKi was accompanied by both GZMB + cytotoxic T cell infiltration and the lower Ly6G + neutrophil infiltration.
Fig. 6
Ruxolitinib inhibited the activation of the JAK-STAT3 pathway in both JAKi and dual ICI + JAKi group. A Treatment timeline in female C57BL/6 J GC peritoneal dissemination model. B Representative macroscopic images of peritoneal dissemination of mice on day 11. The extent of peritoneal dissemination was consistent across all groups. C Western blot analysis and quantification of p-STAT3 protein bands following JAKi administration. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, one-way ANOVA with Tukey’s multiple comparisons test
Fig. 7
Dual ICI in combination with JAKi reshaped the immunosuppressive microenvironment. Representative images of IHC staining of p-STAT3, cytotoxic T cells, M2 macrophages, neutrophils, and Tregs on day 11. Scale bar: 100 μm; *p < 0.05, **p < 0.01, ***p < 0.001, one-way ANOVA with Tukey’s multiple comparisons test. ns not significant, Kruskal–Wallis test with Dunn’s multiple comparison
留言 (0)