
記住我
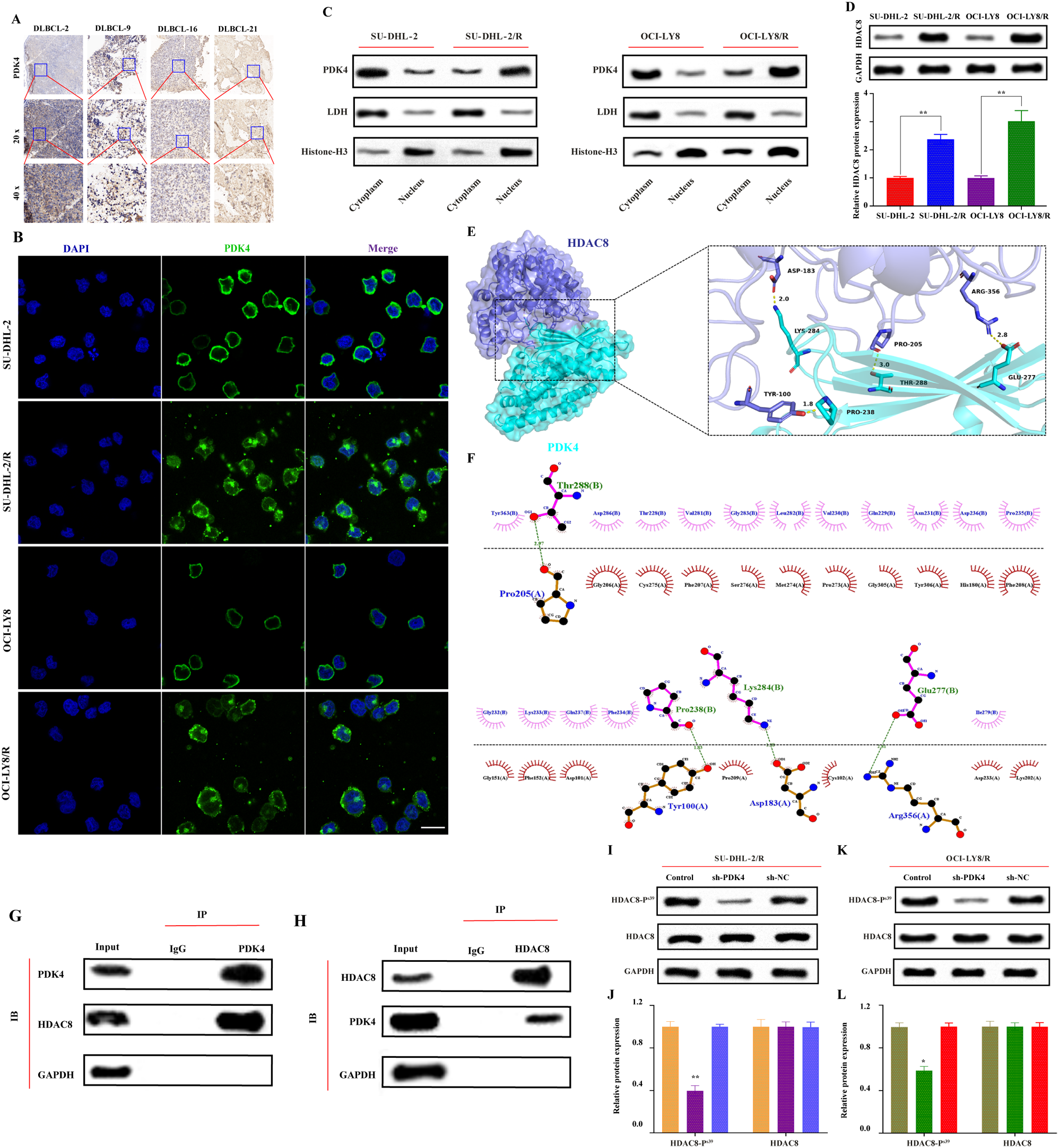


To clarify the role of AP-1 TFs in PCa progression, we investigated the level of the master factor JUN in tissue microarrays (TMA) of low and high progressive human prostate tumors by immunohistochemistry (IHC). We performed semi-quantitative analysis and categorized each tumor based on JUN levels from 0 (absent), 1 (low-grade), 2 (medium-grade) to 3 (high grade) (Fig. 1a). Patients were divided into present (n = 29 + 6 censored subjects) (JUNpresent) and absent (n = 32 + 8 censored subjects) (JUNabsent) cohorts and correlated with biochemical recurrence (BCR) data (Supplementary Fig. 1a). PCa progression is marked by histological changes of the tumor architecture and is categorized by Gleason scoring [34]. We observed a gradual decrease of JUN protein abundance from healthy tissue to primary tumors (low Gleason; Gleason score 5–6), reaching the lowest JUN expression state in advanced tumor stages (high Gleason; Gleason score 7–9) (Fig. 1a). The correlation between JUN protein and patient BCR status revealed a significantly (p = 1.8e-02) diminished BCR-free survival in patients with low JUN, whereas high JUN levels were associated with increased survival probability (Supplementary Fig. 1a). We next mined a publicly available transcriptome dataset ([35]; n = 140) and stratified PCa patients into high-risk and low-risk groups as defined by the prognostic index and characterized by a significant difference in relapse-free survival (RFS) using the SurvExpress webtool [36] (p = 4e-04) (Supplementary Fig. 1b). We investigated JUN mRNA expression in the high- and low-risk groups and found significantly (p = 1.3e-30) higher JUN among low-risk patients compared to the high-risk group (Supplementary Fig. 1c). To explore JUN levels in advanced stages of PCa, we used the Taylor dataset [35], comprising primary tumors of different progression stages and Gleason scores (n = 131) as well as healthy prostate tissue (n = 29). Compared to healthy tissue, we observed higher levels of JUN in early disease stages with Gleason scores 5–6 and significantly decreased expression of JUN in high grade tumors (p = 3e-03; Gleason scores 7–9) (Fig. 1b). Concordantly, JUN was highly expressed in primary tumors (n = 131; n = 65) but significantly lower expressed in PCa metastases (n = 19; n = 25) as observed in two independent datasets (Fig. 1c-d; p = 1.3e-02; [35]; p = 5.3e-09; [37]). We next investigated levels of JUN, JUNB and FOS and observed a comparable regulation (Supplementary Fig. 1d-e). Metastatic CRPC and neuroendocrine PCa (NEPC) present aggressive tumor subtypes that emerge under androgen deprivation therapy and are associated with poor prognosis. We compared levels of JUN and its related TFs FOS and JUNB in primary (n = 715) and metastatic (n = 320) PCa [38], including CRPC and NEPC (Fig. 1e-f, Supplementary Fig. 1f). The tumor-subtype and stage-dependent expression of JUN was highly significant when comparing healthy and primary (p = 2.8e-05), primary and metastatic CRPC (p = 2.6e-43) and primary and metastatic NEPC (p = 5.3e-04) (Supplementary Fig. 1g), suggesting JUN as a potential marker of progressive subtypes of PCa. In addition, our survey revealed higher levels of JUN in primary PCa associated with low Gleason scores than healthy prostates (Supplementary Fig. 1g-h), suggesting a gradual change of JUN levels in PCa development and progression. Our data implicate that JUN and other AP-1 factors except MAF and MAFB may act as suppressors rather than drivers of PCa which was reflected by hazard ratios (HR) calculated from RFS (Supplementary Fig. 1i).
Fig. 1
JUN levels are correlated with prostate cancer progression stages. a Left panel: Representative immunohistochemistry (IHC) images of tissue microarrays (TMAs) investigating human prostate tumors (n = 60) with high or low Gleason scores stained for JUN protein. Scale bars indicate 150 µm (top row) and 30 µm (bottom row), images are presented in 16.8 × (top row) and 80.0 × magnification (bottom row). The area used for the higher magnification is indicated by the rectangle. Right panel: Violin plot showing JUN expression divided in absent (0), low-grade (1), medium-grade (2) and high-grade (3) in healthy (no Gleason score), low Gleason (Gleason score 5–6) and high Gleason (Gleason score 7–9) TMA samples. b JUN mRNA levels in high (Gleason score ≥ 7) and low (Gleason score < 7) grade human prostate tumors. Data were retrieved from [35]. Significance was determined by an unpaired, two-sided t-test or one-sided Anova. c High and low JUN levels significantly (p = 1.3e-02) discriminate primary prostate tumors (n = 131) (red) and metastases (n = 19) (blue). Data were retrieved from [35]. d High and low JUN levels significantly (p = 5.3e-09) discriminate primary prostate tumors (n = 65) (red) and metastases (n = 25) (blue). Data were retrieved from [37]. Significances in c-d were determined by an unpaired, two-sided t-test. e Principal component analysis (PCA) of prostate tumors of different developmental stages comprising normal prostate tissue, primary tumors and primary (p) and metastatic (m) CRPC and NEPC tumors. Datasets from [38]. f Overlay of JUN expression with PCA clustering from e). JUN levels are color coded from high expression (yellow) to low expression (blue). g Kaplan–Meier survival analysis of TCGA-PRAD [33] tumors (n = 333) assessing levels of JUN and PTEN. Hazard ratios (HR) were determined by Cox-regression analysis: HR(JUNhigh vs. JUNlow) = 0.461, p = 3.8e-02 and HR(PTENhigh vs. PTENlow) = 0.307, p = 1.5e-03. Statistical testing was done with a logrank test. h Co-analysis between PTEN expression (RNA-Seq by Expectation–Maximization (RSEM) and PTEN protein level reverse-phase protein array (RPPA)). i Co-analysis between PTEN protein level (RRPA) and JUN expression (RSEM)
Mutations in the tumor suppressor PTEN are considered as main drivers of oncogenic transformation and malignancy in PCa [4]. As PTEN loss is highly correlated with increasing Gleason score and associated with activation of several downstream processes, primarily via hyperactivation of PI3K/AKT and inactivation of AR signaling [4], we next investigated synergistic effects of additional JUN alterations. We applied the KMplot tool to assess RFS of PCa patients ([33]; n = 333) that were stratified into four risk groups JUNhigh/PTENhigh, JUNhigh/PTENlow, JUNlow/PTENhigh and JUNlow/PTENlow. We observed that patients featuring low levels of JUN and PTEN showed the lowest survival probability whereas patients with high JUN and PTEN expression presented with the most favorable prognosis. In contrast to singular JUN depletion, downregulation of PTEN alone resulted in intermediate survival probabilities confirming its role as main oncogenic driver in PCa (Fig. 1g). Finally, we surveyed reverse-phase protein array (RPPA) data of the TCGA-PRAD cohort [33] and observed that PTEN protein correlated well with PTEN mRNA levels whereas we identified an inverse relationship between PTEN and JUN levels (Fig. 1h-i). Although loss of JUN alone is not sufficient to cause significant changes in survival probability, our data suggest that the absence of PTEN promotes JUN to a survival-determining factor in PCa patients.
Genetic depletion identifies a tumor-suppressive role of JUN in prostate cancer developmentAs patients presenting with low expression of JUN and PTEN showed severely reduced survival rates, we next sought to elucidate the mechanistic role of JUN in the development of PTEN-deficient PCa and employed a Pten floxed murine model of PCa (Fig. 2a) [26, 39]. The homozygous deletion of murine Pten via the Probasin (Pb) Cre recombinase [27] mirrored 20% of all primary human PCa cases with homozygous loss of PTEN (Fig. 2a, PbCre/Pten). The PE of homozygous mutants developed hyperplasia that progressed into prostate adenocarcinoma between 12 and 29-weeks of age [39]. We inter-crossed a floxed Jun mouse strain where the sole exon is flanked by loxP sites [25] (Fig. 2a, Jun) to generate 4 individual genotypes. This enabled comparison of prostate tissue of wildtype (wt) mice to either Jun (JunPEΔ/Δ), Pten (PtenPEΔ/Δ) or Jun/Pten (JunPEΔ/Δ; PtenPEΔ/Δ) double knockout mice (Fig. 2a, colored F1 mice). We examined protein extracts of whole prostates and observed a significant increase in levels of phosphorylated (S73) and total JUN in PtenPEΔ/Δ, whereas notable JUN expression was absent in wt prostates (Fig. 2b). We also confirmed efficient Cre-mediated deletion of Jun alone (JunPEΔ/Δ) and in combination with Pten (JunPEΔ/Δ; PtenPEΔ/Δ) (Fig. 2b). As a verification of functional Pten deletion, we detected robust activation of the PI3K/AKT pathway in PtenPEΔ/Δ and JunPEΔ/Δ; PtenPEΔ/Δ mice as assessed by analysis of phosphorylated AKT (pAKTS473) levels (Fig. 2b).
Fig. 2
Jun-deficiency fosters the progression of Pten-loss induced tumors. a Top: Schematic representation of mouse models used in the study. Homozygous loss of Pten or Jun was achieved by a Probasin promoter-controlled Cre recombinase (PbCre)-mediated ablation of floxed exons 4 and 5 (Pten) or exon 1 (Jun). Bottom: established and investigated genetic models. Wildtype (PbCre+/+; wt) and mice with single knockout of Pten (PbCretg/+; PtenPEΔ/Δ) and Jun (PbCretg/+; JunPEΔ/Δ) were compared with double knockout (PbCretg/+; JunPEΔ/Δ;PtenPEΔ/Δ). PE = prostate epithelium; tg = transgene; Δ = knockout. b Western blot analysis of phosphorylated (pJUNS73 and pAKTS473) and total JUN and AKT. β-TUBULIN served as loading control. Protein lysates of entire organs (n = 3 biological replicates) from 19-week-old wt, PtenPEΔ/Δ, JunPEΔ/Δ and JunPEΔ/Δ;PtenPEΔ/Δ were investigated. c Top row: H&E stainings of 19-week-old wt, PtenPEΔ/Δ, JunPEΔ/Δ and JunPEΔ/Δ;PtenPEΔ/Δ prostates. Scale bars indicate 60 µm (top row) and 2 µm (second row), images are presented in 40.0 × (top row) and 600.0 × magnification (second row). Black rectangles represent the area used for the zoom image below. Bottom row: IHC with an antibody against JUN in 19-week-old prostates of all four experimental groups. Scale bars indicate 30 µm; images are presented in 100.0 × magnification. d Macroscopic images of 19-week-old dissected prostates of wt, PtenPEΔ/Δ, JunPEΔ/Δ and JunPEΔ/Δ;PtenPEΔ/Δ mice. e Box plot showing the weights of prostates in grams between wt, PtenPEΔ/Δ, JunPEΔ/Δ and JunPEΔ/Δ;PtenPEΔ/Δ 19-week-old animals (n = 20). Significance was determined with an unpaired, two-sided t-test. f Kaplan–Meier survival analysis of wt, PtenPEΔ/Δ, JunPEΔ/Δ and JunPEΔ/Δ;PtenPEΔ/Δ animals. Biological replicates are indicated and the cumulative survival (%) is shown. Statistical significance was calculated with a logrank test. g Organs (heart, lung, liver, spleen, kidney, lymph nodes and brain) of 39-week-old wt, PtenPEΔ/Δ, JunPEΔ/Δ and JunPEΔ/Δ;PtenPEΔ/Δ mice were stained with H&E and analysed for metastatic lesion formation. The number of metastases detected in each tissue are shown
To investigate the morphological architecture of prostates upon Jun deletion in the PCa mouse model, we analyzed histological sections by hematoxilin and eosin (H&E) staining (Fig. 2c, top panel). Both wt and JunPEΔ/Δ animals showed physiological growth patterns and morphology, characteristic for the respective prostate lobes. In PtenPEΔ/Δ and JunPEΔ/Δ; PtenPEΔ/Δ prostates, we observed hyperplastic epithelium growing in cribriform patterns into the lumen. Both groups showed anisocytosis, anisokaryosis and alterations in nucleus-to-cytoplasmic ratios, but largely without invasion of the stroma.
Next, we analyzed JUN levels in prostates of all genotypes. Supporting our immunoblot results, IHC revealed increased levels of total JUN predominantly in the PE of PtenPEΔ/Δ mice and absence in epithelial cells of JunPEΔ/Δ and JunPEΔ/Δ; PtenPEΔ/Δ (Fig. 2c, bottom panel). We assessed the effects of Jun deficiency on tumor burden and survival by morphological and survival analyses. Macroscopically, prostates from PtenPEΔ/Δ and JunPEΔ/Δ; PtenPEΔ/Δ mice were notably enlarged as compared to wt or JunPEΔ/Δ prostates (Fig. 2d). This finding was corroborated by prostate weight analysis (Fig. 2e). The additional deletion of Jun on the Pten-deficient background resulted in even higher prostate weights, hinting at JUN’s potential function as a tumor-suppressor in murine PCa development. We performed a Kaplan–Meier survival analysis where overall survival or the occurrence of the discontinuation criteria according to the guidelines of the 3Rs principles were defined as the endpoint of the experiments (Fig. 2f) [40]. We observed comparable survival probabilities of wt and JunPEΔ/Δ mice (p = 5.4e-01) but a significantly decreased survival of PtenPEΔ/Δ (mean survival 85.3 weeks, p = 6e-03) as compared to wt mice. Remarkably, the survival of PtenPEΔ/Δ mice was significantly (p < 1e-04) reduced by the additional deletion of Jun. JunPEΔ/Δ; PtenPEΔ/Δ mice exhibited a mean survival of 67.2 weeks. Despite the significantly reduced survival rates in JunPEΔ/Δ; PtenPEΔ/Δ mice, we did not detect metastatic lesions in the analysed genotypes (Fig. 2g). We therefore conclude that Jun-deficiency alone is not sufficient to induce prostate tumorigenesis, but causes a significant increase in tumor burden and a significant reduction in overall survival in combination with Pten knockout. The results of our murine PCa model reinforce our observations from human PCa samples, suggesting that JUN acts as a tumor-suppressor in PCa.
To determine whether aberrant cellular proliferation contributes to enhanced tumor growth in JunPEΔ/Δ; PtenPEΔ/Δ-deficient prostates, we assessed the number of Ki67+ epithelial cells by IHC. Although we noticed higher Ki67 levels in JunPEΔ/Δ; PtenPEΔ/Δ tumors by trend, the difference was not significant (p = 1.3e-01) when compared to PtenPEΔ/Δ prostates (Supplementary Fig. 2a). To investigate the effects of JUN ablation in vitro, we utilized the CRISPR/Cas9 technology in the human PCa cell lines DU145 (PTEN wildtype) and PC3 (PTEN mutated). We designed three individual guide RNAs for the JUN locus (Supplementary Fig. 2b) and used lentiviral transduction of empty vector (EV) and guide RNA (G1, G12, G14) plasmids. We identified varying efficiencies of JUN knockout in bulk cultures of DU145 and PC3 cell lines (Supplementary Fig. 2c, e) and no significant differences in cellular proliferation (Supplementary Fig. 2d, f). We confirmed the results of unchanged proliferation in single clones of both cell lines which were selected according to complete loss of JUN protein (Supplementary Fig. 2g-j). The in vivo and in vitro results indicate that proliferation may not be the primary biological process influenced by JUN during PCa progression.
Transcriptome profiling reveals JUN-mediated alterations in senescence-associated secretion and immune responseTo elucidate the tumor cell-specific molecular programs regulated by JUN in vivo, we performed transcriptome profiling of PE cells across all four experimental murine groups (Fig. 2a). To obtain a homogenous epithelial fraction, we enriched prostate lysates for the Epithelial cell adhesion molecule (EpCAM) showing a uniform expression in PE cells (Fig. 3a, Supplementary Fig. 3a) via magnetic cell separation [32] (Fig. 3b, Supplementary Fig. 3b). The correlation analysis revealed high congruence between JunPEΔ/Δ; PtenPEΔ/Δ and PtenPEΔ/Δ tumor and wt and JunPEΔ/Δ samples (Fig. 3c).
Fig. 3
Transcriptome profiling of genetic models reveals a JUN-dependent regulation of innate immunity. a Representative immunofluorescence (IF) image of a wt murine prostate for the epithelial marker EpCAM (green). DAPI (blue) is shown as a nuclear stain. Top image: 40.0 × magnification, scale bar represents 60 µm; Bottom image: 147.5 × magnification, scale bar represents 20 µm. b Overview of sample preparation for transcriptome profiling of wt, PtenPEΔ/Δ, JunPEΔ/Δ and JunPEΔ/Δ;PtenPEΔ/Δ prostate samples of 19-week-old animals. An antibody against the epithelial marker EpCAM was used to separate single cell suspensions of minced and digested prostates into EpCAM positive (pos) and negative (neg) fractions by magnetic cell sorting. EpCAMpos cells were used for RNA-seq expression profiling. c Heat map showing correlation analysis of tumor samples described in b) regarding global similarity of samples. The Pearson correlation coefficient (R) is shown (color coded). d Gene onthology (GO)-enrichment analysis of differentially expressed genes (DEGs) showing the top differentially regulated pathways between PtenPEΔ/Δ and JunPEΔ/Δ;PtenPEΔ/Δ. Significance as shown by FDR is color coded, enriched (positive normalized enrichment score (NES)) or depleted (negative NES) processes are indicated. Asterisk represents non-significant pathways (ns). e Heat map showing SenMayo genes most significantly (p ≤ 1e-02) regulated among PtenPEΔ/Δ and JunPEΔ/Δ;PtenPEΔ/Δ prostates. f GSEA enrichment analysis using the Guccini_core_SASP gene set in PtenPEΔ/Δ versus JunPEΔ/Δ;PtenPEΔ/Δ and PtenPEΔ/Δ versus wt animals. g Heat map representation of wt, PtenPEΔ/Δ, JunPEΔ/Δ and JunPEΔ/Δ;PtenPEΔ/Δ samples showing DEGs. “Innate immunity”, FDR = 7.64e-05; “Immune system”, FDR = 2.77e-04 and “Extracellular space”, FDR = 6.60e-03 related processes most discriminated the groups. Genotypes and expression levels are color coded. h GO-enrichment analysis of DEGs showing the regulation of innate immune cells such as neutrophil granulocytes. Significance as shown by p-value is color coded, enriched (positive NES) or depleted (negative NES) processes are indicated. Shown are the signaling pathways enriched in PtenPEΔ/Δ tumors compared to wt (left side) and JunPEΔ/Δ;PtenPEΔ/Δ tumors compared to PtenPEΔ/Δ (right side)
We next performed a comparative analysis of JunPEΔ/Δ; PtenPEΔ/Δ and PtenPEΔ/Δ prostate samples to discern JUN-dependent programs potentially contributing to PCa formation. Our survey revealed 1706 (p.adjust < 5e-02) differentially expressed genes (DEGs) with top 102 genes being up- (log2fold change ≥ 1) and top 91 genes downregulated (log2fold change ≤ -1; Supplementary Table 1). DAVID analysis of top genes showed increased “innate immunity” and “immune system processes” but decreased secretory-, extracellular matrix- and immune-related processes. Notably, Jun ranked among the top 10 downregulated genes confirming the successful knockout in epithelial cells (Supplementary Table 1). Gene set enrichment analysis (GSEA) revealed immune system-related processes, IL-6/STAT3 signaling and senescence-associated gene signatures among the most enriched processes in PtenPEΔ/Δ prostates which were significantly depleted in JunPEΔ/Δ; PtenPEΔ/Δ (Fig. 3d). Our previous work suggested that activation of IL-6/STAT3 signaling and of the downstream acting p19ARF–MDM2–p53 axis contributed to senescence in PtenPEΔ/Δ prostates [12]. We therefore investigated the enrichment level of different senescence signatures including “oncogene-induced senescence” (OIS), “SASP” signatures and the novel “SenMayo” gene signature, consisting of 125 previously identified senescence/SASP-associated factors. SenMayo genes are transcriptionally regulated by senescence and allow identification of senescent cells across tissues [41]. SenMayo genes were significantly (qval = 2.40e-02) enriched in PtenPEΔ/Δ prostates and depleted (qval = 2.64e-02) in JunPEΔ/Δ; PtenPEΔ/Δ tumors (Fig. 3d). Among the depleted SenMayo genes in Jun-deficient PtenPEΔ/Δ prostates, we identified chemokines such as Ccl3, Ccl4 and Ccl8, along with pro-inflammatory cytokines such as Il1b and Tnfa (Fig. 3e). As these secreted cytokines and chemokines represent well described SASP factors, we next investigated a SASP core gene signature previously described in a Pten-deficient prostate model [42]. Using GSEA, we indeed detected enrichment of the SASP core signature in Pten-deficient prostates which was reverted in JunPEΔ/Δ; PtenPEΔ/Δ animals (Fig. 3f). To investigate further aspects of JUN-dependent regulation of senescence in Pten-deficient murine prostates, we stained formalin-fixed paraffin embedded (FFPE) material with the senescence markers p16INK4A, p21CIP1/WAF1 and Galactosidase beta 1 (GLB1) (Supplementary Fig. 3c). We did not observe differences in the amount of p16INK4A positive cells between PtenPEΔ/Δ and JunPEΔ/Δ; PtenPEΔ/Δ tumors, but found significant changes in staining patterns. While we detected prominent nuclear staining in PtenPEΔ/Δ samples, JunPEΔ/Δ; PtenPEΔ/Δ revealed predominantly cytoplasmic localization, hinting at a potential inactivation of p16INK4A via nuclear export [43]. In wt and JunPEΔ/Δ prostates, we observed a weak lobe-dependent expression pattern of p21CIP1/WAF1. Conversely, in PtenPEΔ/Δ and JunPEΔ/Δ; PtenPEΔ/Δ samples, p21CIP1/WAF1 was expressed in each individual epithelial cell, with no discernible difference between the two groups. GLB1 staining displayed its characteristic granular expression pattern prompting us to quantify percentage of positive area however we found no significant difference between PtenPEΔ/Δ and JunPEΔ/Δ; PtenPEΔ/Δ groups. Apart from changes in the p16 staining pattern, we found no significant deregulation of the classic senescence-associated cell cycle markers, implicating that JUN affects the SASP but not senescence-associated cell cycle arrest.
As our results suggest JUN-dependent activation of the IL-6/STAT3 axis and our previous study connected loss of activated STAT3 in Pten-deficient PCa to increased tumor burden via disruption of senescence [12], we sought to analyze STAT3 tyrosine 705 (Y705) phosphorylation (pSTAT3Y705) in the Jun-deficient background. We indeed detected reduced levels of pSTAT3Y705 in both stroma (p = 5.0e-04) and epithelial cells (p < 1.0e-04) of JunPEΔ/Δ; PtenPEΔ/Δ compared to PtenPEΔ/Δ tumors (Supplementary Fig. 3d upper panel, Supplementary Fig. 3e) while total STAT3 levels remained constant (Supplementary Fig. 3d, lower panel). Our findings provide evidence that loss of JUN accompanied by reduced activation of STAT3 bypasses SASP and subsequently amplifies the tumor load in JunPEΔ/Δ; PtenPEΔ/Δ animals. We suggest an interplay of JUN and STAT3 mediating senescence-associated secretion of inflammatory factors in PCa in vivo, reinforcing JUN’s proposed function as a pioneering factor of senescence [23].
JUN deficiency in the PCa mouse model leads to downregulated chemotaxis of innate immune cellsWe next compared JunPEΔ/Δ; PtenPEΔ/Δ and PtenPEΔ/Δ prostate samples to uncover additional JUN-dependent biological processes involved in PCa formation. A stringent selection identified ~100 significantly deregulated genes (padj ≤ 1.0e-03, FClog2 ≤ -1.2; n = 59/ FClog2 ≥ 1.2; n = 46; Supplementary Table 3) and uncovered innate immunity and other immune system-related processes as most distinguishing between JunPEΔ/Δ; PtenPEΔ/Δ and PtenPEΔ/Δ prostate tumors (Fig. 3g). Amongst the innate immunity and immune system cluster, gene onthology (GO)-enrichment analysis indeed confirmed immune system-related signatures that were activated in PtenPEΔ/Δ and significantly reduced by Jun-deficiency (Fig. 3h). Innate immunity-related processes are complex and encompass more than 2000 publicly available human and mouse annotated genes [44]. We defined a core immunity-related signature by GSEA applying 645 innate immunity-related genes and investigated the enrichment specifically in PtenPEΔ/Δ prostates. The analysis revealed 111 genes, of which 26 were significantly (p < 1.0e-03) differentially expressed between PtenPEΔ/Δ and JunPEΔ/Δ; PtenPEΔ/Δ prostates (Fig. 4a, top panels, Supplementary Fig. 4a, Supplementary Table 4). Using the “Hallmark Inflammatory response” signature, we uncovered a similar pattern as the majority of genes from both signatures were significantly (p < 1.0e-03) elevated in PtenPEΔ/Δ and depleted in JunPEΔ/Δ; PtenPEΔ/Δ prostates (Fig. 4a, bottom panels, Supplementary Fig. 4a). Hence, the homozygous loss of Pten was accompanied by inflammation and inflammatory response likely driven by increased levels of Il1b, Nlrp3 and chemokines such as Ccl5.
Fig. 4
JUN expression determines the level of immune cell infiltration of Pten-loss driven tumors. a Heat map showing JUN-dependent regulation of genes related to innate immunity (upper panel) and inflammatory response (lower panel) in wt, JunPEΔ/Δ, PtenPEΔ/Δ and JunPEΔ/Δ;PtenPEΔ/Δ prostates. JUN-dependent core factors such as Il1b, Nlrp3 and Ccl5 are highlighted. b Heat map presenting the JUN-dependent regulation of genes involved in migration and chemotaxis of neutrophil granulocytes in PtenPEΔ/Δ and JunPEΔ/Δ;PtenPEΔ/Δ prostates. Genotypes and expression levels in a-b are color coded. c Expression levels (log2, FPKM) of Ccl3, Ccl8 and Il1b are significantly (Ccl3, p = 2.4e-04; Ccl8, p = 9.7e-05 and Il1b, p = 5.0e-03) reduced in EpCAM+ cells of JunPEΔ/Δ;PtenPEΔ/Δ prostates. Significance was determined by an unpaired two-sided t-test. d Single-sample GSEA analysis using the M5 signature of Broad Institute’s molecular signature database (MsigDB) revealing enrichment of macrophage- and neutrophil-associated properties in PtenPEΔ/Δ compared to JunPEΔ/Δ;PtenPEΔ/Δ prostates. e Western blot analysis of NLRP3 and non-cleaved Pro-IL-1β in all four experimental groups in biological replicates. β-ACTIN served as loading control. f Multiplex immunoassay of homogenized prostate samples of 19-week-old wt, JunPEΔ/Δ, PtenPEΔ/Δ and JunPEΔ/Δ;PtenPEΔ/Δ animals for analysis of IL-1β levels in pico grams (pg)/ml of indicated biological replicates. Statistical testing was done with one-way Anova, significant p-values are indicated
Cells of the innate immune system, including neutrophil granulocytes, mast cells and macrophages serve as the primary defense against infections and consequently recruit T and B cells to infection sites [45]. Among the DEGs of PtenPEΔ/Δ versus JunPEΔ/Δ; PtenPEΔ/Δ prostates, we identified neutrophil movement-specific gene signatures that play a crucial role in the recruitment of immune cells (Fig. 4b) [46]. We observed that cytokines involved in chemotaxis of immune cells such as Ccl3, Ccl8 and Il1b were significantly deregulated between groups (Fig. 4c). To further dissect the potentially involved immune cell subsets, we conducted single sample GSEA using the M5 ontology gene sets signature from the molecular signature database (MsigDB). We identified enrichment of macrophage- and neutrophil-specific gene signatures characterized by cellular activities such as migration, activation/differentiation and enhanced expression indicating production of MIP1α/CCL3 and GM-CSF. Moreover, single sample GSEA revealed processes related to other immune cell subsets such as mast cells, myeloid cells and CD8+ T cells that were significantly enriched in PtenPEΔ/Δ compared to wt prostates and depleted in JunPEΔ/Δ; PtenPEΔ/Δ (Fig. 4d). This implicates JUN in the control of inflammatory states during PCa progression. We validated the JUN-dependent regulation of IL-1β, TNF-α and NLRP3, all involved in the regulation of inflammatory response processes by immunoblot and cytokine analyses (Fig. 4e-f and Supplementary Fig. 4b).
To further examine the apparent shifts in immune system-related transcriptomic signatures, we assessed granulocytic or lymphocytic cell infiltrations based on microscopic characteristics in H&E staining of all four genotypes (Supplementary Fig. 4c). We detected no or low-grade infiltration by inflammatory cells in wt and JunPEΔ/Δ specimens. In contrast, PtenPEΔ/Δ mouse prostates exhibited increased levels of high- and middle-grade infiltrations, which were significantly mitigated in JunPEΔ/Δ; PtenPEΔ/Δ prostates. Increased immune cell infiltration of PtenPEΔ/Δ prostates as identified by histo-pathological analysis therefore supported the results of transcriptome profiling. This highlights the importance of JUN in the regulation of inflammation by affecting the secretion of pro-inflammatory cytokines in Pten-deficient PCa.
Epithelial JUN deficiency modulates the migration of innate immune cells from the peripheryTo investigate the distribution and abundance of infiltrating immune cells, we performed IHC stainings. Neutrophils and inflammatory monocytes were stained using the antibody clone NIMP-R14, which targets the specific cell surface markers and differentiation antigens Ly-6G and Ly-6C (Fig. 5a). In PtenPEΔ/Δ prostates, we observed high numbers of neutrophils migrating from the blood vessels across the stroma into the epithelium, where they predominantly accumulated, and subsequently advanced into the lumen. In JunPEΔ/Δ; PtenPEΔ/Δ prostates, we detected significantly (p < 1.0e-04) less neutrophils in the stroma and epithelium, but the migration patterns remained consistent with PtenPEΔ/Δ tumors (Fig. 5b). In contrast, macrophages, stained by the marker F4/80 were primarily located in the stroma, with no significant differences between the groups (Fig. 5c-d). We observed significantly (p = 4.0e-04) less macrophages infiltrating the epithelium in prostates with additional deficiency of Jun. In conclusion, PtenPEΔ/Δ displayed a highly immune infiltrated phenotype, which was substantially reverted in prostates with additional deficiency of Jun. This observation suggests that JUN may be essential for tumor cell recognition by innate and consequently adaptive immune cells.
Fig. 5
Histological analysis of infiltrating immune cells reveals downregulated innate immune response in JunPEΔ/Δ;PtenPEΔ/Δ prostates. a Representative images of IHC stainings of NIMP-R14, a pan-marker of neutrophil granulocytes, indicating high neutrophil infiltration of PtenPEΔ/Δ prostates, reverted by the additional loss of Jun in JunPEΔ/Δ;PtenPEΔ/Δ prostates. Top row: 20.0 × magnification, scale bar represents 150 µm; Bottom row: 63.0 × magnification, scale bar represents 40 µm. b Quantification of NIMP-R14+ neutrophils in epithelium (left) and stroma (right). A significantly decreased (p < 1e-04) infiltration of neutrophils in tumors and adjacent stroma of JunPEΔ/Δ;PtenPEΔ/Δ prostates is evident. c Representative images of IHC stainings for the pan-marker of macrophages F4/80. A high infiltration of PtenPEΔ/Δ prostates and adjacent stroma by macrophages is evident and reverted by the additional loss of Jun in JunPEΔ/Δ;PtenPEΔ/Δ prostates. Top row: 40.0 × magnification, scale bar represents 60 µm; Bottom row: 100.0 × magnification, scale bar represents 30 µm. d Quantification of F4/80+ macrophages in epithelium (left) and stroma (right). A significantly decreased (p = 4e-04) infiltration of macrophages in tumors but not adjacent stroma (p = 8.3e-01) of JunPEΔ/Δ;PtenPEΔ/Δ prostates is evident. e Representative images of IHC stainings of B cell infiltration using the pan-marker CD79b. A high infiltration of stroma adjacent to PtenPEΔ/Δ prostates by CD79b+ B cells is evide
留言 (0)