
記住我

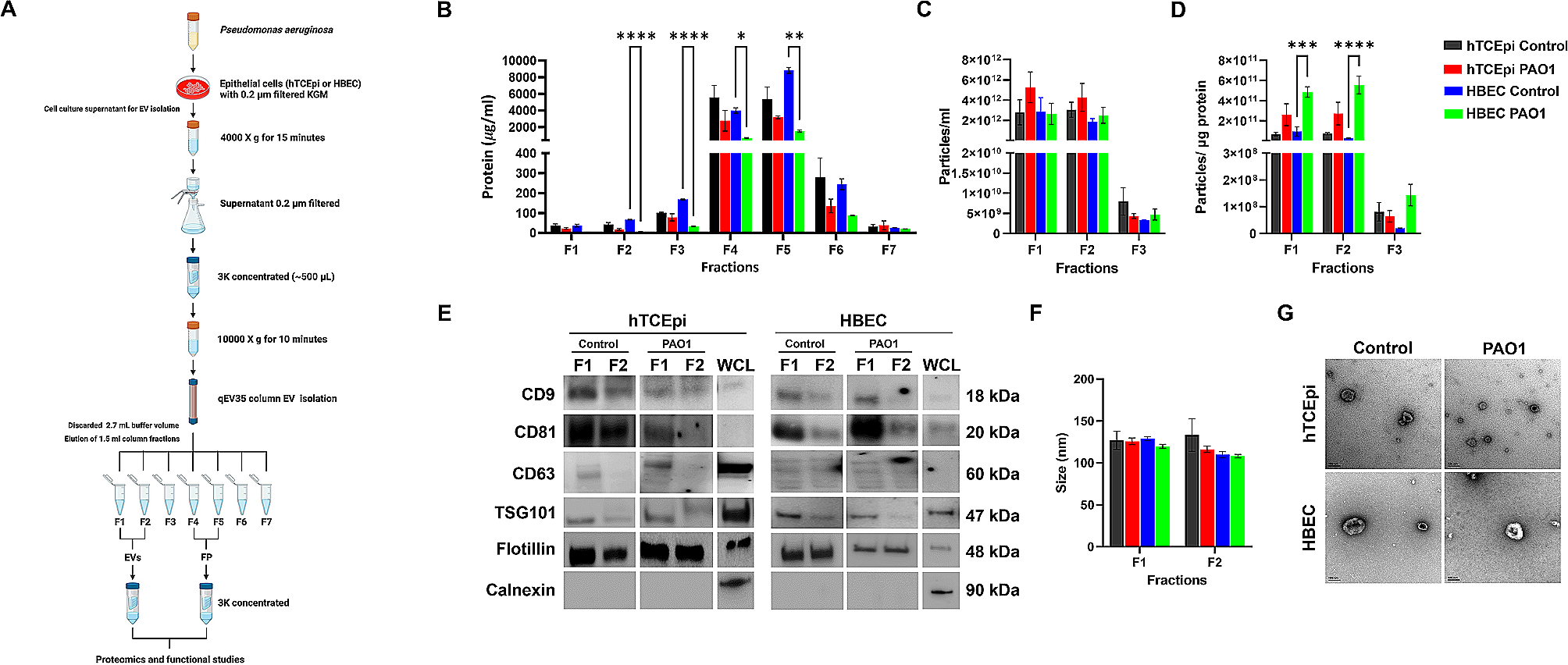



It has been assumed that CD6 regulates T cell activation following its binding to the APC-expressed ligand, CD166 [35], and its interaction in cis with the TCR complex at the IS [39] (Fig. 1a). To determine the key components of CD6 involved in the regulation of T cell signaling and to clarify their specific roles, we constructed plasmids to express: (i) the CD6 wild-type (WT) molecule; (ii) the naturally-occurring isoform CD6Δd3; (iii) a cytosolic-truncation mutant (CD6Δcyt); and (iv) a binding- and signaling-disabled mutant that lacks both d3 and the cytosolic tail (CD6Δd3Δcyt) (Fig. 1b). Je6-NF-κB::eGFP cells, a Jurkat cell line engineered to express an eGFP reporter for NF-κB transcription factor activation and thus to measure T cell activation [43], were transduced to express these CD6 variants at equivalent levels (Fig. S1a).
Each Je6 cell line was subjected to interaction with Raji cells that naturally express CD166 (Raji-CD166+) or with Raji cells defective for CD166 expression (Raji-CD166neg) [42], in the absence or presence of the superantigen (sAg) staphylococcal enterotoxin E (SEE). Je6 and Raji cells in these different combinations were allowed to interact for 24 h, and the activation of NF-κB on the Je6 cells was assessed by measuring eGFP levels. Activity profiles for each Je6:Raji pair are shown as raw data in Fig. 1c, and converted to activation indexes (eGFP values of cells interacting in the presence of SEE / eGFP values of cells interacting in the absence of SEE) in Fig. 1d. The most conspicuous observation is that the cytosolic tail of CD6 considerably restrains the strength of T cell activation, thus providing support to our previous studies [9]. In particular, cells that express CD6 mutants lacking the cytosolic tail (CD6Δcyt and CD6Δd3Δcyt) were significantly more activated than those expressing tail-containing CD6WT (∼ 4-fold) or CD6Δd3 (∼ 2 to 2.5-fold) (Fig. 1d).
Fig. 1
CD6 extra- and intra-cytoplasmic domains contribute differently to T cell activation. a Schematic representation of CD6 extracellular interactions during antigen-driven T cell activation (Created with BioRender.com). Binding of CD6 to APC-expressed CD166 involves the interaction between the membrane proximal domain (d3) of CD6 with the membrane distal IgSF domain of CD166. This interaction is abolished when CD6 is expressed as the CD6Δd3 isoform. T cell activation results in the activation of several transcription factors, including NF-κB and NFAT. b Schematic representation of CD6 constructs expressed in Je6-NF-κB::eGFP and Je6-NFAT::eGFP reporter cells (Created with BioRender.com): CD6 wild-type (WT), the naturally-occurring isoform lacking the ligand binding domain (CD6Δd3), a cytoplasmic-truncation mutant (CD6Δcyt), and a binding- and signaling-inert mutant (CD6Δd3Δcyt). Raji cells were used as antigen presenting cells, either naturally expressing CD166 (Raji-CD166+) or engineered to be defective of CD166 (Raji-CD166neg). c Je6-NF-κB::eGFP cells expressing CD6WT and mutants were allowed to interact for 24 h at 37 °C with Raji-CD166neg or Raji-CD166+ cells, previously incubated, or not, with the sAg staphylococcal enterotoxin E (SEE). T cell activation was assessed by flow cytometry analysis of NF-κB::eGFP up-regulation. d Normalized fold induction (activation index) of geometric mean of fluorescence intensity (gMFI) values in (c) of cells interacting in the presence of SEE over the gMFI values of the interactions without SEE. e, f gMFI values (e) and activation indexes (f) of Je6-CD166neg-NF-κB::eGFP cells expressing CD6WT and mutants interacting for 24 h at 37 °C with Raji-CD166neg or Raji-CD166+ cells, with or without SEE. T cell activation was assessed by flow cytometry analysis of NF-κB::eGFP up-regulation. g, h gMFI values (g) and activation indexes (h) of Je6-NFAT::eGFP cells expressing CD6WT and mutants interacting for 24 h at 37 °C with Raji-CD166neg or Raji-CD166+ cells, with or without SEE. T cell activation was assessed by flow cytometry analysis of NFAT::eGFP up-regulation. Each experiment was performed at least four times, with technical duplicates, and data were analyzed with two-way ANOVA, followed by Turkey’s multiple comparison test or one-way ANOVA followed by Dunnett’s multiple comparisons test. Results of gMFI for each Je6-Raji pair condition (c, e, g), and statistical analysis relative to the respective CD6WT (d, f, h). **P < 0.01, ****P < 0.001
On the other hand, the presence of the CD166-binding domain, d3, did not seem to contribute to T cell positive signaling. Comparing Je6 cells expressing CD6WT or CD6Δd3, it appears that the presence of d3 contributed, if anything, to a somewhat more restrained response (Fig. 1d). Interestingly, this effect was ligand-independent, i.e., Je6-CD6WT cells were slightly less activated than Je6-CD6Δd3 cells even when both cells interacted with Raji-CD166neg cells (∼ 1.5-fold, Fig. 1d, left, red squares vs. blue diamonds, P = 0.029, Mann-Whitney test for direct comparisons), a condition in which none of the two CD6 isoforms is able to contact CD166.
Raji cells do not express CD318, the alternative ligand for CD6, but a possible confounding factor in the previous analysis is that CD166, besides being expressed on APCs, is up-regulated on activated T lymphocytes and Jurkat cells [30]. During Je6:Je6 contacts in these multicellular assays, CD166 binding to CD6 might impact on the activation signals, possibly by removing CD6 away from the Je6:Raji interface. To bypass this concern and address the direct effect of binding of CD6 expressed on Je6 only to CD166 expressed on Raji, we generated Je6-NF-κB::eGFP cells devoid of CD166, by CRISPR/Cas9 engineering (Fig. S1b). The resulting Je6-CD166neg-NF-κB::eGFP cells were subsequently transfected to express each of the CD6 forms (Fig. S1c). Each CD6WT- or mutant-expressing Je6-CD166neg cell line was then incubated for 24 h with Raji-CD166neg or Raji-CD166+ cells, in the absence or presence of SEE, and the activation of NF-κB assessed by eGFP levels. Activity profiles for each Je6-CD166neg:Raji pair are shown as raw data in Fig. 1e and activation index values in Fig. 1f. No different behavior of NF-κB activation was observed between Je6-CD166neg and Je6-CD166+ cells (Fig. 1d, f), showing that CD166 expressed on Jurkat cells did not interfere with the signaling mechanisms directly regulated by CD6.
The cytosolic tail of CD6 contains multiple signaling motifs and it is therefore expected to impact on different pathways. We explored the effect of CD6 mutants on the NFAT pathway by transducing the vectors encoding CD6WT and CD6 mutants into Je6 cells engineered to express an eGFP reporter for the activation of NFAT [43] (Fig. S1d). Like before, Je6-NFAT::eGFP cells expressing the various CD6 forms were conjugated with Raji-CD166neg or Raji-CD166+ cells, without or with SEE, and the activation of NFAT was assessed by eGFP levels. Activity profiles for each Je6:Raji pair are shown as raw data in Fig. 1g and as activation indexes in Fig. 1h. Although the activation indexes of the NFAT reporter were higher than those of the NF-κB reporter in the same conditions (50 to 150-fold in NFAT vs. > 10 to 50-fold in NF-κB), the tendencies were similar between the two reporter cell sets. However, the specific impact of CD6 seems less relevant on the NFAT pathway: cells that express CD6 tailless mutants are still more activated than those expressing tail-containing CD6, but the differences were less pronounced (ratio between tailless and tail-containing CD6 forms is ∼ 1.5 to 2-fold) and did not reach statistical significance (P = 0.07) (Fig. 1h).
Overall, using a cellular system where model Jurkat T cells are allowed to directly interact with APCs without the interference of activating or blocking mAbs or other reagents, we can conclude that upon activation triggered by SEE presentation, the cytosolic tail of CD6 is a major contributor for the inhibitory effects that help to control and tune different signaling pathways, and that binding of CD6 to CD166 expressed on either APCs or on other Jurkat cells during multicellular clustering has no positive impact on signaling strength.
The cytosolic domain of CD6 contains a diverse array of signaling inhibitory elementsThe cytosolic tail of CD6 contains nine tyrosine residues that upon phosphorylation potentially mediate interactions with SH2 domain- or phosphotyrosine binding (PTB) domain-containing enzymes or adaptors that participate in signal transduction. We constructed CD6 mutants with individual Y to F substitutions, and analyzed their signaling properties. Many of the individual substitutions had no detectable impact on signal transduction, as measured by the activation indexes of NF-κB reporter cells (Fig. 2a, Fig. S2a). However, substitutions of three tyrosine residues, Y503, Y556 and Y662, resulted in slight increases in activation, i.e., loss of inhibitory signaling, indicating that these specific tyrosines likely play a role in the inhibitory effects of CD6. The simultaneous substitution of all 9 tyrosine residues (CD6-YtoF) resulted, however, in a CD6 form that only slightly differed in signaling properties from the WT molecule. This suggests that stimulatory effectors may conceivably bind to some of the phosphorylated tyrosine residues; disruption of their binding could result in the abrogation of any potential co-stimulatory signaling, rendering on aggregate of all CD6 tyrosine substitutions a close to neutral effect. By contrast, cells expressing the CD6Δcyt mutant, which also lacks all 9 tyrosine residues, were much less repressed than the CD6-YtoF-expressing cell line. This shows that sequences other than phosphotyrosine motifs must definitively contribute to repressing activation.
A recent report suggested that the CD6 sequence S482DSDY486 (SDSDY) could inhibit TCR-proximal signaling, most likely by promoting CD6 intracellular binding to PTB domain-containing proteins [44]. Moreover, the combination of serine (S482A and S484A) and tyrosine (Y486F, Y629F and Y662F) substitutions added to the relief of CD6-mediated inhibition [44]. We thus expressed CD6 mutants of the canonical SDSDY sequence containing the appropriate Ser and Tyr substitutions, namely SDSDF (= Y486F), ADADY and ADADF on Je6-NF-κB::eGFP cells, and combinations of these with double substitutions of the C-terminal tyrosines Y629 and Y662 (ADADY Y629F Y662F; ADADF Y629F Y662F), and measured the activation of these cells upon interaction with Raji-CD166+ cells in the absence or presence of SEE. The activation indexes of Je6-CD6-ADADY- and Je6-CD6-ADADF cells were only slightly increased, compared with Je6-CD6WT cells. However, Je6 cells expressing either ADADY Y629F Y662F or ADADF Y629F Y662F displayed an increase in the activation index of > 2-fold compared with CD6WT-expressing cells (Fig. 2b, Fig. S2b), suggesting a stronger effect of these serine residues towards the inhibitory effect of CD6.
Altogether, the data show that the cytosolic domain of CD6 contains tyrosine phosphorylatable sequences that individually influence signal transduction only minimally, and that certain combinations of Ser and/or Tyr residues are more impactful in inhibiting signaling. The integrity of the entire cytosolic tail, possibly containing additional undetermined motifs, appears, nonetheless, to be absolutely crucial for the full inhibitory function of CD6.
Fig. 2
The cytosolic domain of CD6 contains a diverse array of signaling Inhibitory elements. a Je6-NF-κB::eGFP cells expressing different CD6 mutants, each having a Y-to-F substitution, or a mutant with all nine CD6 tyrosine residues substituted by phenylalanines (YtoF), were co-cultivated with Raji-CD166+ cells, previously incubated without or with SEE. Activation indexes are represented in the graph. b Je6-NF-κB::eGFP cells expressing CD6 specific serine and tyrosine substitutions were co-cultured for 24 h with Raji-CD166+ cells, previously incubated without or with SEE. Activation indexes are represented in the graph. Each experiment was performed at least five times, with technical duplicates. Statistical differences shown for CD6WT and analyzed with two-way ANOVA, followed by Turkey’s multiple comparison test. **P < 0.01, ****P < 0.001
The role of the 3rd SRCR domain of CD6 on cell adhesiveness, immunological synapse organization and signaling inhibitionTwo unexpected conclusions relating to the function of the third SRCR domain (d3) of CD6 were drawn from the analysis of Fig. 1. The first was that the physical binding to CD166, one of the assumed key events for CD6 function and which is mediated by d3, did not have a major impact on T cell activation, either inhibiting or enhancing signaling. Since Je6-CD6Δd3 cells do not express the ligand-binding domain, it was already anticipated that these cells would show the same activation index either interacting with Raji-CD166neg or with Raji-CD166+ cells (Fig. 1d, blue diamonds). But the same behavior was unexpectedly observed for cells expressing CD6WT, i.e., Je6-CD6WT cells exhibited the same activation index irrespective of interacting with Raji-CD166neg or with Raji-CD166+ cells (Fig. 1d, red squares). A similar result was also seen in the NFAT system (Fig. 1h).
However, we noticed that in the absence of SEE (i.e., non-activated Je6 cells), the eGFP levels of only Je6-CD6WT cells interacting with Raji-CD166+ cells were higher from baseline (∼ 2.5-fold), when compared to all other CD6-mutant-expressing cells (Fig. 1c, plots in Raji-CD166+/no SEE). This effect, also seen in the NFAT system (Fig. 1g, baseline 1.5-fold higher in Je6-CD6WT than in all other cells), suggested that binding of CD6 to CD166 might actually have an impact not during activation, but prior to antigen presentation.
We thus addressed the relevance of the CD6-CD166 pair on cell adhesiveness during antigen-dependent and -independent priming of T cells. E6.1 Jurkat cells expressing CD6WT were placed in contact with Raji-CD166neg or Raji-CD166+ cells in the absence or presence of SEE, and the percentage of cell conjugates (doublets) was measured by imaging flow cytometry (representative images shown in Fig. 3a). Whereas after presentation of SEE (activated cells), the presence of CD166 did not result in additional number of doublets, in the case where no SEE was present, the percentage of E6.1-CD6WT cells that conjugated with Raji-CD166+ cells was significantly higher than those conjugating with Raji-CD166neg cells (Fig. 3b, left). When using E6.1 cells expressing CD6Δd3, this effect was not reproduced (Fig. 3b, right). These results confirm that the contribution of CD6-CD166 binding to cell adhesion is mostly relevant still in the absence of TCR engagement.
Fig. 3
The third SRCR domain of CD6 promotes T cell adhesion to APCs prior to antigen presentation. a E6.1 Jurkat CD6WT cells, previously stained with a cell tracker dye (CMFDA), were conjugated with Raji-CD166neg or with Raji-CD166+ cells, pre-loaded or not with SEE. Representative images of cell:cell fixed conjugates in the absence or presence of CD6-CD166 engagement, assessed by imaging flow cytometry. b Graphs show the percentage of conjugates formed between E6.1-CD6WT (left) or E6.1-CD6Δd3 cells (right) with Raji-CD166neg or Raji-CD166+ cells pre-loaded or not with SEE. Results are from five independent experiments with the representative mean ± SD, analyzed with unpaired Student’s t test with Welch’s correction. **P < 0.01
The second unexpected observation from Fig. 1 was, as mentioned earlier, that although both natural isoforms, CD6WT and CD6Δd3, contain the same intracellular signaling motifs, Je6-CD6WT cells were slightly more repressed than Je6-CD6Δd3 cells, even when both interacted with Raji-CD166neg cells. We therefore investigated whether these isoforms had different distributions at the IS that could explain their slightly different activation profiles. E6.1 Jurkat cells expressing CD6WT or CD6Δd3 were put in contact with SEE-loaded Raji-CD166neg or Raji-CD166+ cells, and CD6 molecules were visualized by immunofluorescence. Inside the first 5 min of cell contacts, CD6WT and CD6Δd3, as well as CD3, were evenly distributed over the whole cell surface in all conditions [Fig. 4a, left panels (Start)]. After 30 min of cell contacts, CD6WT molecules in Jurkat cells interacting with Raji-CD166+ cells were transported to the central supramolecular activation cluster (cSMAC) of the synapse, as anticipated (Fig. 4a, 2nd row, 30 min; Fig. 4b, red-bordered column). Unexpectedly, CD6WT also targeted to the interface between Jurkat and Raji-CD166neg cells (Fig. 4a, 1st row, 30 min; Fig. 4b, red column). This indicates that the synaptic localization of CD6 occurs independently of binding to CD166, in complete contrast with the current paradigm [38, 40]. This conclusion was further strengthened by the observation that CD6Δd3 was also directed to the synapse when the respective Jurkat cells conjugated with either Raji-CD166neg or Raji-CD166+ cells (Fig. 4a, 3rd and 4th rows, 30 min; Fig. 4b, blue and blue-bordered columns).
Fig. 4
Translocation of CD6 to the immunological synapse is independent of ligand-binding. a Representative confocal images of CD6WT and CD6Δd3 localization at synapses formed between E6.1 and Raji-CD166neg or Raji-CD166+ cells. Cells were fixed within the first 5 min of initiation of contacts (left images) and after 30 min of cell interactions (right images). CD3 is shown in green and defines the cSMAC, CD6 (detected by MEM98, anti-CD6d1) is visualized in red. Merge images allow to define CD3 and CD6 co-localization. Scale bar, 5 μm. b Percentage of conjugates in which CD6WT or CD6Δd3 accumulate at the IS formed with SEE-loaded Raji. Results are from at least 40 conjugates per condition, obtained from at least three independent experiments represented as mean ± SD. Quantification was done by three different examiners, blind to the experimental conditions. c Quantitative analysis of CD3 and CD6 pixel co-localization upon conjugate formation between SEE-loaded Raji and E6.1 cells expressing CD6WT or CD6Δd3. Pearson’s correlation coefficients from the synaptic region were generated using ImageJ software and JACoP plugin. Values obtained from conjugates from three independent experiments are represented as mean ± SD and analyzed with one-way ANOVA followed by Turkey’s multiple comparison test. Representative 3D projections are shown in Expanded View (Movie S1). ***P < 0.005 and ****P < 0.001
However, despite that both CD6WT and CD6Δd3 were identically transported to the IS, thus independently of the ligand, quantitative analysis of their precise co-localization with CD3 revealed that the interaction with the TCR signaling machinery is not identical (Fig. 4c and Movie S1). In particular, CD6Δd3 co-localization with CD3 was significantly lower than that of CD6WT. On the other hand, CD6WT showed similar levels of interaction or proximity with CD3 independently of the presence of CD166 on the APC. These results suggest that the described cis-interaction between CD6 with the TCR complex [39] may be established through the d3 domain, and independently of CD6 binding to CD166 on an opposing cell. Considering the inhibitory signaling nature of CD6, by interacting with the TCR signaling apparatus (directly or indirectly), CD6WT, more than CD6Δd3, may contribute to restrain T cell activation.
Translocation of CD6 to the immunological synapse is dependent on its cytosolic tailAs, surprisingly, the extracellular domain is not mandatory for the synaptic localization of CD6, we questioned whether the translocation to the IS would depend on cytosolic motifs. E6.1 cells expressing CD6Δcyt were brought into contact with SEE-loaded Raji-CD166neg or Raji-CD166+ cells, and the localization of CD6Δcyt was visualized by immunofluorescence. Within the first 5 min, CD6Δcyt in most cells was dispersed across the entire cell surface and only concentrated at the cell interface in a very small fraction of contacts [Fig. 5a, b (Start)]. However, most CD6Δcyt molecules not only did not translocate to mature synapses after 30 min of conjugation with Raji-CD166+ cells (Fig. 5a, b), but also had no influence on the number of cell conjugates in the absence (left columns) or the presence (right columns) of SEE (Fig. 5c), even though CD6Δcyt contains the CD166-binding domain. These observations confirm that the synapse-targeting motifs are confined to the CD6 cytosolic tail, and that lack of the tail impairs both CD6 translocation to the IS and any important contribution to cell adhesion.
Translocation to the IS of CD6 mutants carrying Y to F substitutions was then measured in transfected E6.1 cells co-cultured with Raji-CD166+ cells, pre-incubated with SEE (Fig. 5d). The different mutants showed only slight variations in the synaptic-localization when compared with CD6WT, with the exception of the Y629F substitution that reduced IS-localization by approximately 40%. Because there was no correlation between the loss of synaptic localization and the contribution to signaling inhibition of the individual mutants (evaluate Fig. 2a vs. Figure 5d), it appears that these are separate events regulated by different CD6 cytosolic motifs. Interestingly, when all tyrosine residues were replaced by phenylalanines (CD6-YtoF), the decline in synapse-targeting was very pronounced. This suggests that extensive tyrosine phosphorylation of the CD6 cytosolic tail is essential for building the CD6 interactome that delivers this molecule to the synapse, although no tyrosine substitution alone is capable of deconstructing the complex. Importantly, the fact that CD6-YtoF did not target to the synapse (Fig. 5d) while still being a strong inhibitor (Fig. 2a), demonstrates that CD6 is inhibitory independently of localizing to the IS.
Fig. 5
CD6 translocation to the immunological synapse is dependent on the cytoplasmic tail. a Representative confocal images of conjugates formed between SEE-loaded Raji-CD166neg or Raji-CD166+ cells with E6.1 cells expressing CD6Δcyt. Cells were fixed within the first 5 min of initiation of contacts (left images) and after 30 min of cell interactions (right images). CD3 is shown in green and defines the cSMAC, CD6 is shown in red. Merge images reveal no specific co-localization between CD3 and CD6Δcyt. Scale bar: 5 μm. b Percentage of conjugates in which CD6Δcyt accumulates at the IS established with SEE-loaded Raji-CD166neg or Raji-CD166+ cells. Differences not significant, unpaired Student’s t test with Welch’s correction. c Frequency of conjugates (cell doublets) formed between E6.1-CD6Δd3 and Raji-CD166neg or Raji-CD166+ cells, pre-loaded or not with sAg, measured by imaging flow cytometry. Results are from five independent experiments, with the representative mean ± SD. Differences not significant, unpaired Student’s t test with Welch’s correction. d Percentage of conjugates in which CD6 accumulates at the IS established between SEE-loaded Raji and E6.1 cells expressing CD6 with tyrosine to phenylalanine substitutions. b, d Results are from at least 40 conjugates per condition, obtained from at least three independent experiments. Quantification was performed by three different examiners, blind to the experimental conditions, and data were analyzed using one-way ANOVA, followed by Dunnett’s multiple comparison test. *P < 0.05, ****P < 0.001
Overall, the cytosolic domain is responsible for the architecture of the interactome and inhibitory function of CD6, using non-coincident motifs. Tyrosine-based motifs are fundamental to assemble the CD6 signalosome and to deliver CD6 to the IS, whereas signaling inhibition is only marginally affected. The integrity of the cytosolic tail is therefore necessary for the integrated and concerted actions of CD6 in regulating T cell activation.
留言 (0)