
記住我






Telomeres are essential for genome integrity. PARP1 is recruited to telomeres to repair internal telomere DNA breaks and base lesions15,16 and can promote telomere fusions via the alternative end-joining mechanism17. Furthermore, telomeric DNA terminates with a single-stranded 3′ overhang and recessed 5′ end, a potent trigger of PARP1 activity, that is shielded by the telomere-binding and protection complex, Shelterin18. In assessing patterns of nuclear ADP-ribosylation by immunofluorescence, we observed a pronounced accumulation of PAR foci in TARG1-deficient cells that colocalized with telomeres, marked by the telomere-binding protein, TRF1 (Extended Data Fig. 1a). Considering the strong evidence for PARP1 dependent activities at telomeres, as well as the putative link of TARG1 in reversing DNA-ADPr, this prompted us to investigate whether the modification occurs on telomeric DNA in human cells.
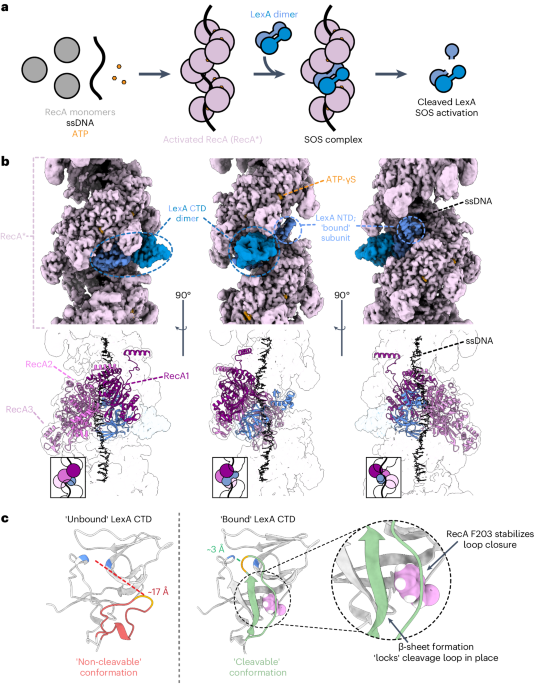
The chemical properties of the ADP-ribose linkage in DNA have precluded its detection and genomic assignment by conventional immunoprecipitation-PCR or next-generation sequencing methods14. We used a region-specific extraction (RSE) methodology that relies on the specific hybridization of a biotin-conjugated (AATCCC) oligonucleotide to the TTAGGG-rich telomere overhang followed by streptavidin pulldown from purified genomic DNA (Fig. 1a)19. The enrichment of telomeric DNA repeat sequences by this methodology and preservation of ADP-ribose due to the absence of TARG1 hydrolytic activity could enable the detection of DNA-linked ADP-ribose in dot blot using specific anti-ADP-ribose antibodies. The enrichment of telomeric DNA from control and TARG1-deficient U2OS cells was verified in Southern blot with radiolabeled telomere-specific probes (Fig. 1a). In contrast, Alu repeat sequences were not enriched in the telomere RSE samples (Fig. 1a). Western blotting using specific antibodies confirmed the equal capture of double-stranded (dsDNA) while revealing greater levels of telomere ssDNA from TARG1-deficient cells (Fig. 1a). Notably, ADP-ribose signals were only detected in telomeric DNA captured from TARG1-knockout (KO) cells. The telomere DNA-ADPr signals were DNaseI sensitive (Fig. 1a) and resistant to RNaseA. DNA-ADPr was also detected in samples isolated by telomere RSE from TARG1-deficient IMR90 human fibroblasts that were immortalized with HPV E6/E7 oncoproteins (IMR90E6/E7), as well as in TARG1-deficient HeLa cells (Fig. 1a). U2OS and HeLa cells activate alternative lengthening of telomeres or telomerase-mediated telomere extension mechanisms, respectively. IMR90E6/E7 cells lack a telomere extension mechanism. Therefore, DNA-ADPr may be a general feature of telomeres.
Fig. 1: TARG1 mediated ADP-ribosylation of telomere DNA.
a, RSE methodology (left), western blot, telomeric western dot blot of ADPr, ssDNA, dsDNA and Southern blot of telomeric DNA and Alu repeats of CTRL and TARG1-KO IMR90E6/E7, HeLa and U2OS cells (middle) and quantification of ADPr and ssDNA normalized to the CTRL line (right). b, Western blot and telomeric western dot blot of ADPr and Southern blot of telomeric DNA (left) of U2OS CTRL, TARG1-KO and TARG1-KO cells with expression of either GFP-tagged full-length (WT) TARG1 or TARG1-K84A treated with dimethylsulfoxide (DMSO), PARPi, PARGi or PARPi and PARGi and quantification of ADPr of the samples normalized to TARG1-KO + DMSO (right). c, Western blot and telomeric western dot blot of ADPr and mono-ADPr and Southern blot of telomeric DNA (left) of U2OS CTRL, PARP1-KO and PARP1-KO cells with inducible expression of YFP-tagged full-length (WT) PARP1, PARP1 E998Q or PARP1-EQHA2 after control or TARG1 knockdown and quantification of pan-ADPr and mono-ADPr (right). Quantifications are normalized to U2OS CTRL siCTRL (data not shown). d, Telomeric western dot blot and Southern blot of telomeric DNA (top) and ADPr quantification (bottom) of U2OS CTRL and TARG1-KO treated with H2O2, transiently transfected with TRF1-FokI (D450A or WT), Cas9 D10A (scr or tel) or FAP-TRF1 cells treated with dye or dye and light. Each quantification is normalized to control conditions. e, Telomeric western dot blot of ADPr and Southern blot of telomeric DNA and western blot (top) and ADPr quantification (bottom) of U2OS CTRL and TARG1-KO in asynchronous (Async), serum-starved (starve), G1/S or S phases. Quantification is normalized to TARG1-KO (async). f, Telomeric DNA-ADPr dot blot (top) and ADPr quantification (bottom) of U2OS CTRL and TARG1-KO in either asynchronous (async) or S-phase treated with H2O2 (2 mM, 15 min), hydroxyurea (HU) (2 mM, 1 h) or ATR inhibitor (ATRi) (5 nM, 1 h). Quantification is normalized to TARG1-KO (async). Mean and s.e.m. are shown from three independent experiments in a–f, and groups were compared with a one-way ANOVA followed by Tukey’s multiple-comparisons test for pairwise comparisons.
Reconstituting U2OS TARG1-KO cells with green fluorescent protein (GFP)-tagged WT-TARG1 removed the ADPr signals from telomere DNA. In contrast, the DNA-ADPr signals were not altered on expression of the TARG1 ADP-ribose hydrolysis defective mutant (TARG1-K84A)12,20 (Fig. 1b). By in vitro hydrolase assay, we found that purified full-length TARG1 protein completely removed ADP-ribose from telomere DNA isolated from TARG1-KO cells, while the TARG1-K84A mutant protein did not. This contrasted with the partial in vitro hydrolysis of ADP-ribose either human PARG or bacterial DarG (Extended Data Fig. 1b). This was despite cellular observations in which PARG inhibition (PARGi) stimulated telomere DNA-ADPr in control U2OS cells and enhanced the DNA-ADPr signals detected in TARG1-KO cells, as well as TARG1-WT/K84A complemented cells (Fig. 1b). This can be explained by biochemical studies demonstrating that PARG specifically removes protein-linked PAR chains but that the terminal mono-ADP-ribose (MAR) moiety is removed by specialized MAR hydrolases21. Since TARG1 is capable of cleaving MAR, the additive effect of PARGi on the levels of telomeric DNA-ADPr detected in TARG1-KO cells likely reflects the removal of PAR from DNA by PARG to limit the excessive accumulation of toxic PAR chains21.
We next determined which PARP(s) mediate telomere DNA-ADPr. Olaparib (PARPi) completely abolished the signals in TARG1-KO and TARG1-K84A expressing U2OS cells implicating PARP1 and PARP2 (Fig. 1b). While PARP2 depletion partially reduced telomere DNA-ADPr, PARP1 knockdown abolished virtually all the ADPr signal from telomere DNA samples (Extended Data Fig. 1c). Depletion of HPF1 (histone PARylation factor 1), a cofactor that directs PARP1-2 dependent serine-ADPr of histone H3 and chromatin during the DNA damage response (DDR)22 and ARH3 (ADP-ribosyl-hydrolase 3) a serine-ADPr hydrolase that counteracts PARP1/2-HPF1 (ref. 23) did not affect telomere DNA-ADPr, effectively ruling out their possible contribution (Extended Data Fig. 1c). As with Olaparib treatment and PARP1 knockdown, telomere DNA-ADPr was also not detected in TARG1-PARP1 deficient U2OS cells (Fig. 1c). Introducing WT-PARP1 restored telomere DNA-ADPr. By contrast, the catalytic-dead PARP1-EQHA2 (E988Q, H862A) mutant, did not24. However, expressing PARP1-EQ, a PARP1 mutant that catalyzes MAR but is incapable of PAR chain extension25, restored the DNA-ADPr signal intensity nearly to wild-type (WT) levels as indicated by using recently described MAR-specific antibodies26 (Fig. 1c and Extended Data Fig. 1d). From these combined in vitro and cellular experiments, we conclude that PARP1 is the major catalyst of telomeric DNA-ADPr and that TARG1 is the primary hydrolase responsible for removing ADP-ribose from telomere DNA.
DNA break and S-phase accumulation of telomeric DNA-ADPrWe next asked under which physiological conditions telomere DNA-ADPr is stimulated. First, we exposed control and TARG1-KO U2OS cells to the global genotoxic damaging agent hydrogen peroxide and used TRF1-FokI, Cas9 D10A and FAP-TRF1 to introduce targeted double-strand (ds) DNA breaks, single-strand (ss) DNA breaks and singlet oxygen (1O2) production specifically within telomeres (Extended Data Fig. 1e)27,28. Except for FAP-TRF1, which generates 8-oxo guanine base lesions at telomeres, global and localized telomere DNA damage induced high levels of protein-ADPr and DDR signaling (Extended Data Fig. 1f), but also enhanced telomere DNA-ADPr in TARG1-KO cells (Fig. 1d). This implied that ss- and dsDNA breaks can acutely stimulate telomeric DNA-ADPr.
Previous studies suggested endogenous (that is, DNA damage independent) protein-ADPr oscillates across the cell cycle, peaking in S-phase21,29,30. To examine whether the same patterns are associated with telomere DNA-ADPr, U2OS cells were synchronized in G0 and G1-S by serum starvation or double-thymidine block, respectively, and released G1/S arrested cells into mid-S-phase (Fig. 1e). Efficient cell cycle synchronization was verified by monitoring Cyclin E expression (Fig. 1e) and flow cytometry (Extended Data Fig. 1g). Using the RSE assay, we found that telomeric DNA-ADPr accumulates in S-phase (Fig. 1e). Furthermore, we found that stalling S-phase progression by hydroxyurea (HU)-mediated nucleotide deprivation and intra-S checkpoint activation (Extended Data Fig. 1h) abolished telomeric DNA-ADPr (Fig. 1f). In contrast, acute (1 h) ATR inhibition (ATRi) that provokes replication origin firing31 increased telomeric DNA-ADPr (Fig. 1f). These results provided strong evidence that telomeric DNA-ADPr is associated with DNA replication in S-phase.
DNA-ADPr during telomeric lagging strand maturationTo further define the nature of telomeric DNA-ADPr in S-phase, we applied cesium chloride (CsCl) density gradient centrifugation of IdU (5´-Iodo-2-deoxyuridine)-pulsed U2OS cells treated with PARGi to differentially separate nascent leading and lagging telomeric DNA strands (Fig. 2a). We chose to use acute PARG inhibition as it promotes DNA-ADPr without adversely interfering with cell cycle dynamics (Fig. 1c)32. Fractionated DNAs corresponding to leading, lagging and unreplicated telomeres were slot blotted and probed with telomere probes in Southern blot and western blot using ADPr antibodies. This uncovered that telomere DNA-ADPr occurs selectively on the lagging telomere DNA strand (Fig. 2a).
Fig. 2: DNA-ADPr is associated with lagging strand telomere replication.
a, CsCl density gradient centrifugation of IdU-pulsed U2OS TARG1-KO cells methodology (top) to separate leading and lagging telomeric DNA strands. Southern blot of telomeric DNA and telomeric western ADPr slot blot (bottom left) with quantification of the corresponding strands (bottom right). gDNA, genomic DNA. a.u., arbitrary units. b, Schematic of effects of POT1 knockdown (siPOT1) or FEN1 inhibition (FEN1i) (top). Telomeric western dot blot of ADPr, ssDNA, dsDNA and Southern blot of telomeric DNA of U2OS CTRL and TARG1-KO with either FEN1i or siPOT1 treated with adarotene (Ada) or emetine (Eme). c, Quantification of ADPr (top) or ssDNA (bottom). Quantification of ADPr and ssDNA dot blots (right) normalized to TARG1-KO + DMSO and CTRL + DMSO, respectively. Data represent the mean ± s.e.m., n = 3 biological replicates and groups were compared with a one-way ANOVA followed by Tukey’s multiple-comparisons test for pairwise comparisons. d, DNA from CTRL and TARG1-KO HeLa, IMR90E6/E7 and U2OS cells treated with exoI, ran on a gel and probed for telomeric DNA in native and denaturing Southern blots. Telomeric western dot blot for ADPr (bottom). Quantification of the native/denatured telomeric DNA ratio (left) normalized to corresponding HeLa, IMR90E6/E7 or U2OS CTRL line and quantification of ADPr dot blot (right) normalized to TARG1-KO without exoI treatment for each corresponding cell line. Data represent the mean ± s.e.m., n = 2 biological replicates and groups were compared with a one-way ANOVA followed by Tukey’s multiple-comparisons test for pairwise comparisons. e, SMAT-representative fibers (top) and quantification of telomere fiber length (left), EdU tract length at telomeres (middle) and normalized EdU tract length (right). Data represent the mean ± s.e.m. of three biological replicates (>400 fibers scored (two-tailed Mann–Whitney test)) (left and middle). Data on the right are shown in box and whisker format with minimum and maximum values and median line. f, Telomere length analysis by PFGE of CTRL and TARG1-KO HeLa, U2OS and IMR90E6/E7 cells with indicated population doublings (PDs) following transfection with Cas9 and single-guide RNAs. Red dots indicate mean telomere lengths.
PARP1-mediated ADP-ribosylation has been linked with controlling the maturation of nascent DNA strands and the rate of DNA replication33,34. In imaging experiments, it was shown that FEN1 (Flap Endonuclease 1) inhibition and depletion, which prevents cleavage of the 5′ DNA-RNA flap from nascent Okazaki fragments formed during lagging strand synthesis, increased S-phase ADPr and exacerbated PARP inhibitor cytotoxicity35. These studies provided strong evidence for unligated Okazaki fragments as the primary sources of S-phase ADPr. In agreement with this, we found that FEN1 inhibition (FEN1i) enhanced telomeric DNA-ADPr in TARG1-KO U2OS cells (Fig. 2b,c). Knockdown of FEN1 and DNA ligase I (LigI), which ligates Okazaki fragments, also increased elevated telomeric DNA-ADPr signals (Extended Data Fig. 2a–c). These increases in telomere DNA-ADPr correlated with ss telomere DNA abundance in TARG1-KO cells (Fig. 2b,c and Extended Data Fig. 2b). The FEN1i-induced DNA-ADPr signals were completely ablated after incubating TARG1-KO cells with adarotene or emetine, potent inhibitors of the POLA1 catalytic subunit of DNA Polα36 and lagging strand synthesis37, respectively (Fig. 2b,c). These results indicate that DNA-ADPr is coupled to DNA synthesis on the lagging telomere strand, potentially accumulating at ssDNA discontinuities between unligated Okazaki fragments.
Whereas FEN1 and DNA LigI regulate global lagging strand synthesis, POT1 is a Shelterin complex subunit that binds the single-stranded TTAGGG-rich 3′ overhang and coordinates telomere replication. As with interfering with FEN1-mediated Okazaki fragment maturation, depleting POT1 caused elevated DNA-ADPr that again was sensitive to adarotene and emetine (Fig. 2b,c and Extended Data Fig. 2a). This reinforced the premise that DNA-ADPr accumulation is associated with lagging strand synthesis. However, POT1 also binds the ssDNA 3′ overhang and recruits the CST (CTC1-STN1-TEN1) complex that mediates fill-in DNA synthesis by Polα-Primase38,39. POT1 disruption causes increased telomeric ssDNA 3′ overhangs. This potentially accounted for the increased ssDNA detected at telomeres following POT1 depletion in TARG1-KO cells (Fig. 2b,c). We conducted in-gel Southern blot using radiolabeled telomere probes, this time under native conditions to detect the single-stranded 3′ telomere overhang in HeLa, IMR90E6/E7 and U2OS control and TARG1-KO cells (Fig. 2d). The sensitivity of the signals to bacterial 3′ to 5′ exonuclease (exoI) digestion confirmed that they represent the 3′ overhangs (Fig. 2d). We then denatured and reprobed the same gel to calculate the amount of ssDNA overhang relative to the total telomeric DNA content. This revealed a roughly twofold increase in the ratio of ssDNA corresponding to the 3′ overhang relative to duplex telomeric DNA in the TARG1-deficient cell lines examined (Fig. 2d). Furthermore, expressing WT-TARG1 restored the normal 1/1 ratio of overhang to duplex telomere DNA in U2OS (Extended Data Fig. 2d). The effect of TARG1 on the abundance of ssDNA telomeric DNA corresponding to the overhang raised the question of whether the overhang itself could be ADP ribosylated. We found that parallel exoI treatment of telomere RSE samples from these TARG1-KO cell lines significantly reduced DNA-ADPr (Fig. 2d) indicating that the 3′ ssDNA overhang accounts for a substantial (50–70%) portion of the total ADPr signals detected.
Using conditional POT1-KO 293E cells that can be complemented with either WT or mutant POT1 (Extended Data Fig. 2e,f), we again observed increased telomere DNA-ADPr and ssDNA after depletion of TARG1 (Extended Data Fig. 2g,h). This increase was suppressed following complementation with WT POT1. Similarly, expression of POT1-R83E, a newly described POT1 mutant that cannot protect the 5′ recessed end at the ds–ss-DNA junction from recognition by the DNA damage machinery40 strongly suppressed the accumulation of telomere DNA-ADPr in TARG1-deficient POT1-KO cells. However, the expression of POT1-F62A, a POT1 ssDNA binding mutant whose expression hyperextends the overhang41 (Extended Data Fig. 2g,h) did not suppress DNA-ADPr as efficiently (Extended Data Fig. 2e,f). Collectively, these experiments clarified further that exposed ssDNA is the preferred substrate for PARP1-mediated DNA-ADPr, whose timely degradation is mediated by TARG1.
Impaired telomere replication due to TARG1 deficiencyThe persistence of unhydrolyzed DNA-ADPr due to the absence of TARG1 could represent an obstacle that compromises telomere replication. By conducting SMAT (single-molecule analysis of telomeres), a DNA fiber assay adapted for telomeres, we first observed that the mean telomere fiber length was considerably shorter in TARG1-KO U2OS cells (Fig. 2e). In measuring the ratio of telomere fiber length to the length of fibers labeled with EdU (5-ethynyl-2-deoxyuridine), we determined that net telomere DNA synthesis is considerably reduced in TARG1-KO cells indicative of impaired telomere replication in those cells (Fig. 2e). Following up on the apparent significantly shorter telomere DNA fibers in TARG1-KO U2OS cells, we conducted classical telomere length analysis by pulsed-field gel electrophoresis (PFGE) and Southern blot of DNA from control and TARG1-KO HeLa, U2OS and IMR90E6/E7 cells that were cultured over successive weeks. Under native conditions, we observed progressive, robust increases in the signal intensity in DNA from the TARG1-KO cell lines (Fig. 2f). This confirmed previous results showing increased ssDNA in the RSE samples from TARG1-KO cells. Following denaturation of the same gel, robust telomere shortening was evident in TARG1-KO HeLa, U2OS and IMR90E6/E7 cells (Fig. 2f). The extent of telomere shortening varied from moderate in TARG1-KO IMR90E6/E7 and HeLa to extensive in TARG1-KO U2OS cells. These variations may be due to cell line intrinsic telomere shortening rates following incomplete replication of the lagging strand and potential interference in overhang management that impairs fill-in DNA synthesis by the CST complex39. We did not detect differential 53BP1 accumulation at telomeres in asynchronous control or TARG1-KO U2OS cells. There was, however, a twofold increase in 53BP1 positive telomeres in S-phase synchronized TARG1-KO cells. The modesty of this effect, however, indicates that unhydrolyzed DNA-ADPr does not elicit a robust DDR (Extended Data Fig. 2i). This may be linked to the preferential accumulation of DNA-ADPr on ssDNA (Extended Data Fig. 2g,h), which is a less potent activator of ATR DNA damage-induced signaling than exposed ends42. However, more micronuclei, particularly those containing telomeric DNA fragments and Replication Protein A (RPA), which binds to ssDNA, were observed in TARG1-KO U2OS cells (Extended Data Fig. 2j). The accumulation of these by-products that often form during mitosis demonstrates the enhanced telomere and chromosomal instability due to TARG1 deficiency. These data indicate that removing DNA-linked ADPr from ssDNA, including at the 3′ overhang, is required for efficient telomere replication and telomere length management, thereby contributing to genome integrity.
Direct targeting of DNA-ADPr to telomeresConsidering the pervasive contribution of PARP1 in DNA damage repair and replication, we could not be certain that these defects and alterations in telomere replication and length were truly due to PARP1-directed DNA-ADPr. Yet, discriminating PARP1’s protein and DNA modifying activities is a major obstacle to advancing our understanding of the physiological impact of deregulated DNA-ADPr. No separation of function PARP1 mutant has been identified and will prove challenging. However, we devised an experimental system that enables the direct ADPr of telomeric DNA, without stimulating PARP1 or its protein-ADPr activity.
Thermus aquaticus DarT (also known as DarT2) is a PARP1-like enzyme that catalyzes mono-ADPr of thymidine bases in ssDNA9,12,43. DarT shows no activity on dsDNA and is incapable of protein or RNA-ADPr13. We targeted GFP-tagged DarT to telomeres by fusing it with the telomere-sequence binding protein, TRF1 (Fig. 3a). We also generated a mutant DarT-TRF1 fusion lacking its ADP-ribosyl-transferase activity, DarT-TRF1-E160A. By immunofluorescence, both WT and mutant DarT-TRF1 localized to roughly 80% of detectable telomeres marked by TRF2 (Extended Data Fig. 3a). Mono-ADPr foci were at telomeres only in TARG1-KO U2OS cells expressing WT-DarT-TRF1 (Extended Data Fig. 3b). Both DarT-TRF1 localization and mono-ADPr foci were unaltered by PARPi (Extended Data Fig. 3b). We also used FLAG-tagged T. aquaticus DarG, which hydrolyzes DarT-generated thymidine-linked DNA-ADPr, as well as a catalytically inactive mutant, DarG-K80A13. WT-DarG completely suppressed MAR foci formation by DarT-TRF1 in TARG1-KO cells. In contrast, telomeric MAR foci were readily detected following expression of catalytically inactive-DarG (Fig. 3b). We observed reduced expression of DarG when mutant DarT was co-expressed in cells (Fig. 3c). This is consistent with structural studies revealing DarT–DarG interactions that can be disrupted by mutation of the catalytic domain of DarT43. Western blot analysis confirmed that in contrast to H2O2 treatment, characteristic protein-ADPr smears were not observed following the expression of DarT-TRF1 in control or TARG1-KO U2OS cells, indicating that it does not trigger widespread protein or histone ADPr (Fig. 3c).
Fig. 3: Direct targeting of telomeres via a DarT-TRF1 endotoxin fusion.
a, Schematic of GFP-tagged DarT-TRF1-induced telomeric DNA ADP-ribosylation and subsequent removal by DarG (red). b, Immunofluorescence of U2OS CTRL and TARG1-KO cells after expression of GFP-tagged WT-DarT-TRF1 (WT) or E160A DarT-TRF1 (mut) and FLAG-tagged WT-DarG or K80A DarG (left) and quantification of colocalization of mono-ADPr and GFP-positive telomeres (right). In all conditions, more than 140 cells were analyzed. Scale bars represent 10 μm. c, Western blot of U2OS CTRL and TARG1-KO cells after expression of GFP-tagged WT-DarT-TRF1 (WT) or E160A DarT-TRF1 (mut) and FLAG-tagged WT-DarG (WT) or K80A DarG (mut). U2OS CTRL and TARG1-KO were treated with 2 mM H2O2 and 2 mM H2O2 with PARPi for positive and negative controls, respectively. d, Telomeric western dot blot of ADPr, ssDNA, dsDNA and Southern blot of telomeric DNA. e, Quantification of ADPr (top) normalized to TARG1-KO and ssDNA (bottom) normalized to TARG KO and CTRL, respectively. after expression of vectors in c. Mean and s.e.m. are shown from three independent experiments in b and e, and groups were compared with a one-way ANOVA followed by Tukey’s multiple-comparisons test for pairwise comparisons.
By RSE and ADPr dot blot assay, we observed that WT-DarT-TRF1 stimulated telomere DNA-ADPr above the baseline DNA-ADPr catalyzed by PARP1 in TARG1-KO U2OS cells (Fig. 3d,e). Notably, the expression of WT-DarG or WT-TARG1 suppressed all telomere DNA-ADPr in TARG1-KO U2OS cells (Fig. 3d,e and Extended Data Fig. 3c). This result is particularly significant since DarG exhibits selectivity for thymidine-linked ADPr hydrolase activity on ssDNA substrates8,9,12. Therefore, base-linked ADPr on ssDNA is likely to be the major species of DNA-ADPr present at telomeres that TARG1 removes. Last, the stimulation of ADPr by DarT-TRF1 again coincided with increased telomere ssDNA (Fig. 3d,e). We interpreted this to imply that DarT-TRF1 mediated DNA-ADPr on the lagging strand could stall or uncouple replication of the telomere DNA strands, thereby increasing the availability of ssDNA that could also provide an additional substrate for DarT-TRF1 dependent DNA-ADPr. In agreement, we observed that FEN1i and POT1 depletion enhanced telomere ADPr by WT-DarT-TRF1, but this was suppressed by both adarotene and emetine (Extended Data Fig. 3d). These experiments demonstrate that the DarT-TRF1 system can catalyze DNA-ADPr of thymidine bases in telomeric ssDNA during telomere replication.
Unhydrolyzed DNA-ADPr compromises telomere integrityPhenotypically, we first sought to assess whether telomere-specific DarT-TRF1 induced DNA-ADPr influenced telomere replication by again performing the SMAT assay. Here, we detected moderately shorter tracts of EdU-labeled telomere DNA fibers following the expression of WT-DarT-TRF1 compared with the catalytically inactive DarT-TRF1-E160A mutant in TARG1-KO U2OS cells (Fig. 4a). Normalization of the EdU tract lengths to telomere fiber lengths revealed that WT-DarT-TRF1 expression elicited a modest net reduction in telomere DNA synthesis (Fig. 4a). Yet, we independently corroborated the negative effect of WT-DarT-TRF1 on telomere replication by monitoring EdU incorporation directly at telomeres in U2OS cells (Extended Data Fig. 4a).
Fig. 4: Unhydrolyzed telomere DNA-ADPr impairs telomere integrity.
a, Representative SMAT fibers (left) of U2OS CTRL and TARG1-KO cells after expression of WT-DarT-TRF1 (WT) or E160A DarT-TRF1 (mut), quantification of EdU tract length at telomeres (middle) and normalized EdU tract length (right). Mean and s.e.m. (middle) and a box and whisker plot with minimum and maximum values and median line (right). Results shown are from three independent experiments and groups were compared with Mann–Whitney tests (>200 fibers scored per condition). b, Immunofluorescence images of U2OS CTRL and TARG1-KO cells (left) after expression of GFP-tagged WT-DarT-TRF1 (WT) or E160A DarT-TRF1 (mut) and quantification of percentage of RPA2 + GFP-positive telomeres (right) with more than 150 cells analyzed per condition. Scale bars represent 10 μm. c, CO-FISH representative images of U2OS CTRL and TARG1-KO chromosomes after dox-inducible expression of either GFP-tagged WT-DarT-TRF1 (WT) or E160A DarT-TRF1 (mut). d, CO-FISH quantification of percentage leading (red) and lagging (green) fragile telomeres (left) and percentage telomere sister chromatid exchanges (telomere SCEs) (right) of images in c (>5,000 telomeres scored per condition). e, Colony formation assay representative images of U2OS CTRL and TARG1-KO cells after transient expression of either GFP-tagged WT-DarT-TRF1 (WT) or E160A DarT-TRF1 (mut) and FLAG-tagged WT-DarG (WT) or K80A DarG (mut). f, Quantification of the number of colonies from e normalized to CTRL. Mean and s.e.m. are shown from three independent experiments in b, d and f, and groups were compared with a one-way ANOVA followed by Tukey’s multiple-comparisons test for pairwise comparisons.
The perturbations in telomere replication caused by DarT-TRF1-induced ADPr could lead to the exposure of ssDNA that would be recognized and bound by the RPA complex. In agreement, we observed more RPA2 foci colocalizing with telomeres by immunofluorescence in TARG1-KO cells expressing WT-DarT-TRF1 (Fig. 4b). This was despite acute WT-DarT-TRF1 expression stimulating RPA2 and H2AX phosphorylation, but not phosphorylation of CHK1-serine 317 by ATR kinase (Extended Data Fig. 4b) or significantly altering cell cycle phasing (Extended Data Fig. 4c). Furthermore, we expected that the unhydrolyzed ADPr clusters and defects in lagging strand synthesis could lead to the fragile telomere phenotype44. Telomere fragility is associated with replicative complications at telomeres and can be visualized as discontinuous telomere fluorescence in situ hybridization (FISH) signals on sister chromatids of metaphase chromosomes45. We conducted chromosome-orientation-FISH (CO-FISH) experiments to differentially label leading and lagging telomere strands on metaphase chromosomes and observed elevated frequency of telomere fragility that was more pronounced on lagging strand telomeres in TARG1-KO U2OS and IMR90E6/E7 cells (Extended Data Fig. 4d,e). Furthermore, we observed that this fragile telomere phenotype was significantly exacerbated following the expression of WT-DarT-TRF1 (Fig. 4c,d). In addition to telomere fragility, we found that the TARG1-deficient cell lines exhibited a clear increase in telomere sister chromatid exchanges (Extended Data Fig. 4d,e), which are generated through replicative stress-associated recombination to salvage stalled replication forks. As with telomere fragility, the frequency of telomere sister chromatid exchanges was enhanced following the expression of WT-DarT-TRF1, but not its mutant counte
留言 (0)