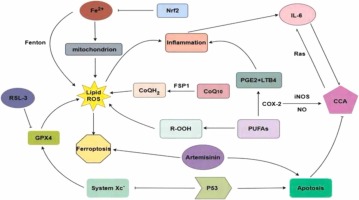
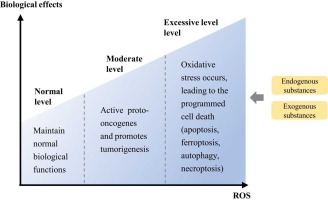
Ferroptosis is a form of planned cell death that distinguishes itself from other mechanisms of cell death, such as necrosis, apoptosis, pyroptosis, and autophagy (Mou et al., 2019a). Necrosis can be triggered by many innate immune signaling pathways (Bedoui et al., 2020), which is an uncontrolled and non-programmed form of cell death (Moujalled et al., 2021). Additionally, non-physiological stressors, such as physical, mechanical, and chemical stress, can also contribute to necrosis (Seo et al., 2021). When the cells underwent necrosis, their plasma membrane experienced rupture, and the organelles became enlarged with a modest level of chromatin condensation (Mou et al., 2019b). Pyroptosis involves the rupture of the plasma membrane, which is triggered by Caspase 1 and Caspase 11. This kind of cell death is characterized by the formation of vesicles on the plasma membrane (Bedoui et al., 2020, Moujalled et al., 2021). Pyroptosis, a common form of cell death, is primarily caused by the aggregation of inflammasomes. This process leads to the cleavage of GSDMD, resulting in the release of IL-1β and IL-18. The gasdermin superfamily plays a crucial role in the occurrence of pyroptosis. Pyroptosis not only hinders the growth of tumor cells, but also creates a favorable environment for their growth, leading to cell swelling, DNA damage, and nuclear shrinkage, therefore facilitating tumor development (Yu et al., 2021a). Apoptosis can be initiated either by the mitochondrial mechanism or the exogenous pathway of death receptors (Moujalled et al., 2021). During apoptosis, cells experience plasma membrane blebbing, a reduction in cell and nucleus volume, and DNA fragmentation and chromatin condensation (Bedoui et al., 2020). The presence of double-membrane autolysosomes in cells is a significant morphological characteristic of autophagy (Mou et al., 2019b). Autophagy is a cellular process that involves the transport of intracellular components and defective organelles to lysosomes for breakdown and recycling. There are three forms of autophagy: macrophage, microautophagy, and chaperone-mediated autophagy (Yang and Klionsky, 2020). Autophagy has a dual role in tumor development. On one hand, it hinders tumor growth by removing aberrant proteins and damaged organelles. On the other hand, it facilitates tumor growth by circulating metabolites (Yang and Klionsky, 2020). In terms of morphology, apoptosis involves the oligomerization of caspases and the fragmentation of DNA. In necrosis, there is a decrease in ATP levels and the activation of RIP1, RIP3, and MLKL. Pyroptosis is characterized by the release of caspase-1 and pro-inflammatory cytokines in significant quantities. Autophagy, on the other hand, leads to an increase in lysosomal activity (Mou et al., 2019b). The morphological characteristics observed in cancer cells, such as the maintenance of nuclear structural integrity and the lack of nuclear condensation or chromatin borders, are useful in differentiating ferroptosis from apoptosis, necrosis, and autophagy after erastin treatment (Xie et al., 2016). Unsaturated fatty acids present on the cell membrane exhibit increased expression following the action of ferrous or esterase. This leads to lipid peroxidation and subsequently triggers ferroptosis (Stockwell, 2022). During the process of ferroptosis, various cellular alterations take place. Initially, the volume of the cell increases, while the mitochondria experience a reduction in size. As a result, the density of both the mitochondrial membrane and electrons increases. In severe instances, the mitochondrial membrane may experience rupture, leading to a reduction or complete absence of the cristae, which are the internal components of the mitochondria (Lei et al., 2022, Gao et al., 2019). Regarding biochemical characteristics, system Xc- experiences inhibition, resulting in a decrease in the number of GPX4. This leads to a reduction in the uptake of cysteine and cystine, as well as a significant consumption of glutathione (GSH) (Zhang et al., 2022a) Consequently, there is an accumulation of intracellular iron ions and reactive oxygen species (ROS), leading to lipid peroxidation (LPO) (Hou et al., 2016, Gao et al., 2016). Additionally, the mitogen-activated kinase system is activated, causing the release of arachidonic acid. Another indication of ferroptosis is the up-regulation of ferroptosis inhibitor protein 1 (FSP1) level (Yu et al., 2021a, Drosten and Barbacid, 2020).
Glutathione (GSH) is an essential chemical synthesized from glutamate, cysteine, and glycine. GSH demonstrates antioxidant and detoxifying properties, which are crucial for preserving cellular well-being. One important element that triggers ferroptosis is the reduction of GSH levels (Shi et al., 2021). Prior studies have shown that the cysteine/GGSH/GPX4 axis has a crucial function in inhibiting ferroptosis. Moreover, System Xc- in neurons demonstrates both advantageous and harmful impacts on GSH production. In addition, the gene that encodes the tumor suppressor protein p53 reduces the expression of the light chain subunit SLC7A11 in the System Xc- transporter, making cells more vulnerable to ferroptosis (Stockwell et al., 2020). Conversely, Nrf2 stimulates the synthesis of GSH (Ursini and Maiorino, 2020). GSH is produced by reducing oxidized glutathione (GSSG) through NADPH (Qiu et al., 2020). Comprehending these pathways is essential for formulating efficient techniques to regulate GSH levels and avert ferroptosis.
GPX4, a Selenoprotein, functions by delivering selenium to cells or animals and so suppressing ferroptosis. The catalyst GPX4 reduces harmful lipid peroxides (L-OOH) into non-toxic alcohols (L-OH), and the interaction with GSH produces GSSG (Zhang et al., 2022b). When GSH is depleted, GPX4 loses its ability to inhibit the buildup of lipid peroxides. As a result, the accumulation of lipid peroxides leads to the occurrence of ferroptosis. When the expression of GPX4 is reduced, the GSH-GPX4 route and the NADPH-FSP1-CoQ10 pathway cooperate rather than oppose each other to enhance ferroptosis (Stockwell et al., 2020).
FSP1 is a lipophilic radical generating oxidoreduction enzyme that seizes antioxidants (RTA) to restrict the dissemination of lipid peroxides and utilizes coenzyme Q10 to impede ferroptosis. FSP1, a protein implicated in the regulation of ferroptosis, has a critical function in suppressing phospholipid peroxidation. It accomplishes this by decreasing the concentrations of CoQ10, a chemical implicated in the process (Doll et al., 2019). Research has demonstrated that GPX4 is located in both the cytoplasm and mitochondria, FSP1 is found in the plasma membrane, and DHODH is present in mitochondria. These proteins are all part of cytoprotective systems that are localized in certain subcellular regions and are resistant to ferroptosis (Zhao et al., 2022). DHODH is an enzyme found on the outer surface of the inner membrane of mitochondria. It slows the process of mitochondrial lipid peroxidation and ferroptosis in a CoQ-dependent manner (Mao et al., 2021).
This review will specifically examine the AMPK, p53, NF2-YAP, mTOR, System Xc-, Wnt, Hippo, Nrf2, and cGAS-STING pathways in relation to ferroptosis. It aims to provide insights and discussions on many aspects of ferroptosis, drawing from these nine commonly seen ferroptosis-related signaling pathways (Wu et al., 2019a).




留言 (0)