
記住我

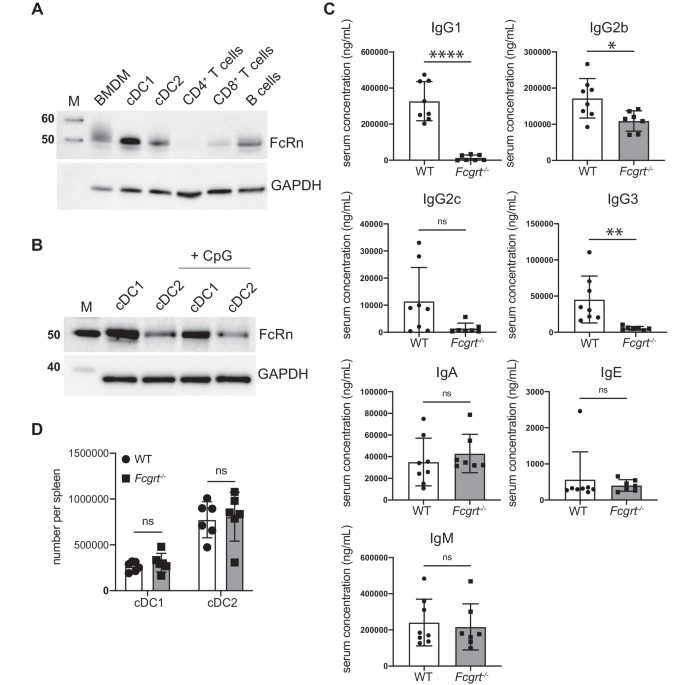
To characterize T cell responses induced by V160 vaccination, we collected blood samples for isolation of RNA, PBMCs, and serum samples from subjects enrolled at one site participating in the V160 phase 2b clinical trial (NCT03486834). One hundred units of V160 or an equal volume of saline solution containing the Merck aluminum phosphate adjuvant (MAPA) was administered as a 0.5 ml intramuscular injection. HCMV-seronegative subjects received intramuscular injections of two doses of V160 plus one dose of placebo (2-dose group), three doses of V160 (3-dose group), or three doses of placebo (placebo group) at day 1, month 2, and month 6 as indicated (Fig. 1a). We detected HCMV IE-1 and pp65 specific T cells in peripheral blood mononuclear cells (PBMCs) collected from V160 and placebo subjects at month 9, and included PBMCs from age- and sex-matched healthy HCMV seropositive (HCMV+) and seronegative (HCMV−) blood donors as controls. Following stimulation by 15-mer peptide pools of IE-1, pp65, or DMSO control (peptide solvent) together with co-stimulators, the PBMCs were analyzed by intracellular cytokines staining (ICS) flow cytometry combined with cell surface markers staining for T cell differentiation status. Sequential gating strategies were used for the analysis of responding CD4 and CD8 T cells that express four effector molecules (IFN-γ, IL-2, TNF-α, and CD107a) (Supplementary Fig. 1). The level of DMSO background responses was shown in Supplementary Fig. 2. Antigen-specific CD4 T and CD8 T cells that expressed four effector molecules were determined as DMSO background subtracted data (Fig. 1b, c). The positive cutoffs were calculated from the median plus two-fold standard error of the mean of the DMSO background responses in 27 analyzed individuals. A positive response is defined as higher than corresponding positive cutoffs. A positive responder is defined as an individual with at least one positive response. Based on the positive cutoffs, there was one marginal positive response among four HCMV− individuals, but no positive response was observed in the placebo group (0/4). About 71.4% (5/7) subjects in the 2-dose group, 80% (4/5) subjects in the 3-dose group, and 57.1% (4/7) donors in the HCMV+ group were positive T-cell responders (supplementary data file 1). Overall, higher percentages of antigen-specific CD8 T cells are detected than antigen-specific CD4 T cells for both the V160 subjects and HCMV+ donors, and the percentages of pp65-specific CD4 T cells are higher than that of IE-1-specific CD4 T cells, which are below positive cutoffs in all individuals (Fig. 1b, c). Due to a small number of subjects and big variations in T-cell responses, our data lack the power for reliable comparisons between 2-dose and 3-dose groups. Notably, two subjects (S26 and S28) with the most potent CD8 T-cell responses (up to 6.2% IFN-γ+ CD8 T cells) have both received 3 doses V160 (Supplementary Fig. 3). In addition, all V160-vaccinated subjects showed strong serum antibody responses to multiple HCMV antigens (including gB, pentamer and whole virion), as well as high serum neutralizing titers against HCMV infection in ARPE-19 cells (Supplementary Fig. 4), including three V160 subjects who had negative T-cell response to both IE-1 and pp65 stimulations according to our positive cutoffs. These results demonstrate that V160 vaccination of seronegative subjects efficiently induces HCMV-specific antibody responses among all subjects while the sizes of T-cell responses are heterogeneous among the subjects.
Fig. 1: V160 induced IE-1 and pp65 T-cell responses as compared to natural HCMV infection.
a Overview of study design, sample collection, experiments, and analyses. Month 9 PBMCs of the 2-dose group (n = 7), 3-dose group (n = 5), and placebo group (n = 4) together with PBMCs of HCMV+ donors (n = 7) and HCMV− donors (n = 4) were analyzed for T-cell responses to IE-1 and pp65 by intracellular cytokine staining (ICS) flow cytometry. b and c Percentages of (b) CD4 T cells and (c) CD8 T cells that expressed four kinds of effector molecules (CD107a, IFN-γ, IL-2, and TNF-α) after DMSO background subtraction (supplementary data file 1). Data are plotted in a box and whiskers style showing the median (center line), the first quartile, and the third quartile together with all data points. Black dots indicate individual responses. Positive cutoffs of specific T-cells were calculated from the median plus a two-fold standard error of the mean of background responses in all individuals (Supplementary Fig. 2) and were shown next to the corresponding legend.
To compare the quality of T cell responses in month 9 PBMCs of V160-vaccinated subjects and HCMV+ donors, we further determined the percentages of T cells that co-expressed each combination of four effector molecules (Fig. 2). Since the IE-1 responding CD4 T cells were below positive cutoffs in all analyzed individuals, we did not include it in the analysis. The overall results of 2-dose and 3-dose V160 subjects were very similar to those of HCMV+ donors. Specifically, pp65 responding CD4 T cells had a dominant CD107a−IFN-γ+IL-2+TNF-α+ subset, while IE-1 and pp65 responding CD8 T cells had two dominant subsets, the CD107a+IFN-γ+IL-2−TNF-α+ and CD107a−IFN-γ+IL-2−TNF-α+ subsets (Fig. 2a, b). Average proportions of responding T cells that were positive for one, two, three, or four effector molecules in each group are shown as pie charts, with the average total frequency of responding T cells indicated in the center (Fig. 2c, d, Supplementary Fig 5). Significant proportions of responding CD4 T cells and CD8 T cells co-expressed four effector molecules, and ~80% of total responding cells co-expressed ≥2 effector molecules in all three groups. Taken together, the V160-induced IE-1- and pp65-specific polyfunctional T-cell responses are similar to those induced by natural HCMV infection.
Fig. 2: Polyfunctional status of IE-1 and pp65 specific T cells induced by V160 and HCMV infection.
Combinatorial analysis of IE-1 and pp65 responding T cells in Fig. 1 was performed. a and b The percentages of (a) CD4 and (b) CD8 T cells that expressed fifteen combinations of four effector molecules (CD107a, IFN-γ, IL-2, and TNF-α) after background subtraction. Data were shown in a box and whiskers style showing the median (center line), the first quartile, and the third quartile together with all data points. Black dots indicate individual responses. c and d Average proportions of antigen-specific (c) CD4 or (d) CD8 T cells in each group that were positive for 1–4 effectors. The average total responsive T cells in each group were shown in the center of the pie chart. Individual data of Fig. 2c, d are shown in Supplementary Fig. 5.
Single-cell transcriptomics of CD3+ T cells after in vitro stimulationTo better understand the long-term T cell response after V160 vaccination, we performed single-cell transcriptomic characterization and paired T cell receptor (TCR) α/β analysis using the 10× Genomics Chromium platform in subject S26, who received 3 doses of V160 and showed very strong T-cell response. We stimulated month 18 PMBCs of S26 (one year post the last vaccination) with peptide pools of IE-1 and pp65; we then sorted the live CD3+ T cells without antigen-specific enrichment and performed scRNA-seq analysis. Unsupervised clustering of the single-cell expression data segregated the T cells into three main CD8 T cell clusters (clusters 1, 7, and 11) and several CD4 clusters (Fig. 3a–c). Cluster 7 and 11 express many cytotoxic effectors such as granzyme B (GZMB), granzyme H (GZMH), natural killer cell granule protein 7 (NKG7), and perforin 1 (PRF1), while C–C motif chemokine receptor 7 (CCR7) expression is low in these clusters (Fig. 3b, c). Additionally, cytokines such as interferon-gamma (IFNG) and X-C motif chemokine ligand 1 (XCL1) are most highly expressed only in cluster 11. Regulatory molecules such as CBLB, an E3 ubiquitin-protein ligase, and the V-set immunoregulatory receptor (VSIR, also known as VISTR) are also most frequently expressed in cluster 11 (Fig. 3b). Conversely, CD4 T cell clusters are highly heterogeneous (Fig. 3b, c). Only two TCR clonotypes have a frequency > 0.5%, which is consistent with the absence of specific T-cell enrichment. These two clonotypes are both located in the cytotoxic CD8 cluster 7 (Fig. 3d). Further analysis is needed to locate V160-responsive T cells among different clusters.
Fig. 3: Transcriptional makeup of CD3+ T cells post V160 vaccination.
a Single-cell transcriptomes of enriched CD3+ T cells from IE-1 and pp65 stimulated month 18 PBMC of S26 were shown as a t-distributed stochastic neighbor embedding (TSNE) plot, colored based on RNA expression cluster assignment by the Louvain graph-based method. b Percentage of cells expressing selected genes within each expression cluster (size scale) and log fold change (FC) of expression within each cluster vs. other clusters (color scale). c Expression levels of selected genes are shown as color overlays on the TSNE plot from (a). d Highlighting of T cells of the two most prevalent clonotypes (with CDR3β sequences shown) on the TSNE plot.
Longitudinal TCR profiling identifies V160 responsive T cell clonotypesTo track T-cell clonal expansion post V160 vaccination, we performed longitudinal blood TCR CDR3β sequencing to samples from the same subject (S26) at 8-time points up to 24 months. We identified 1,936,450 distinct TCR clonotypes from a total of 2,318,537 detected TCR CDR3β sequences. A total of 33,074 unique clones were supported by ≥2 reads in ≥3 samples (time points). The two most prevalent clonotypes (clonotype 1 and 2) represented up to 15% and 1.5% of TCR sequences, and their CDR3β sequences (CASSFRVGELFF and CASSQASGTDTQYF) matched the same dominant clonotypes identified in the single cell paired TCR α/β analysis at month 18 (Fig. 3d). Surprisingly, the clonal fractions of these two clonotypes showed high-level baselines at day 1 and declined at months 2 and 6 after the first and second dose V160 vaccination, which indicates these dominant clonotypes were not induced by V160 (Fig. 4a). In contrast, the clonal fraction of clonotype 3 was undetected at baseline (day 1) and showed a consistent increase after each dose of V160 and peaked at month 12 (Fig. 4a), which suggests that this clonotype underwent V160-induced clonal expansion. To identify other clonotypes that respond similarly to V160 vaccination, we performed unsupervised K-means clustering (K = 100) of the longitudinal relative clonal fraction changes of 33,074 unique CDR3β clonotypes at eight-time points. The clusters containing TCR clones that are consistently expanded post each vaccination (at months 2, 6, and 7) were considered V160 responsive. Most trajectory clusters appeared unresponsive to V160 vaccination according to this criterion (Fig. 4b). Five trajectory clusters that expanded post all three vaccinations were identified as V160 responsive clusters (Fig. 4c, d). The relative clonal fractions of those five responsive clusters were low (<0.5) or undetected at baseline, suggesting that these clones could be de novo induced by V160 vaccination. Trajectory clusters 30, 41, and 35 showed a V160-induced clonal expansion, peaked and declined sharply at month 7 after the last vaccine dose, and we defined these clusters as exhibiting “transient” expansion in response to V160 (Fig. 4c). Trajectory clusters 37 and 52 also showed a V160-induced clonal expansion but demonstrating very slow contraction post the last dose, and we defined these two trajectory clusters as exhibiting “durable” expansion in response to V160 (Fig. 4d, Supplementary data file 2). The “durable” clonotypes (n = 196) collectively constituted about 5% of total TCR sequences at the peak that occurred at month 12, whereas the “transient” clonotypes were more clonally diverse (n = 697) but their prevalence peaked at month 7, reaching only 1% of TCR sequences (Fig. 4e). Overall, using longitudinal TCR profiling, we identified two groups of vaccine-responsive T cells with “durable” or “transient” expansion kinetics.
Fig. 4: V160 elicits durable polyclonal CD8 T cell response.
a–e Time-course blood TCR profiling of a 3-dose V160 subject (S26) at eight time points with dotted lines indicating V160 injections (D1, M2, and M6). a Clonal fractions of three most prevalent clones are shown with CDR3β sequences. b–d Trajectory clustering of time-course TCR profiles. b Three representative clusters of non-responsive T cell clones. Clusters of T cell clones exhibiting (c) “transient” or (d) “durable” clonal expansion in response to V160 vaccination. n denotes the number of unique T cell clones. e Combined clonal fractions of durable and transient T cell clones. f TCR CDR3β sequence clusters are shown as sequence logos, with the number of T clones assigned to each CDR3β sequence cluster and each trajectory cluster shown as a dot plot. g Percentages of V160-responsive CD4 or CD8 T cells in scRNA-seq/scTCR-seq data were identified using the CDR3β as a natural barcode. Significance was assessed by Fisher’s exact test. h Proportions of V160-responsive CD4 and CD8 T cells that were “durable” and “transient” clones. Significance was assessed by Fisher’s exact test. Error bars in g, h indicate mean ± standard error of the mean. i Average percentages of IE-1 or pp65 specific CD4 or CD8 T cells in month 9 (blue circle) or 18 PBMCs (orange circle) from 2-dose (n = 5) and 3-dose (n = 5) subjects were compared side-by-side for expression of four effector molecules (CD107a, IFN-γ, IL-2, and TNF-α) by ICS flow cytometry. Data were background subtracted. Individual responses in i are shown in Supplementary Fig. 6. j Distributions of responsive CD4 vs. CD8 T cells positive for 1, 2, 3, or 4 of the four effector molecules in month 9 vs. 18 PBMCs following IE-1 or pp65 antigen stimulation. The average percentage of total responsive T cells in each group was shown in the center of the pie chart. Data were background subtracted. Individual responses in j are shown in Supplementary Fig. 8.
To validate that the “durable” and “transient” clonotypes are indeed HCMV specific, we analyzed CDR3β sequences for recurring motifs. We identified a consensus CDR3β sequence CASSRLAxxxDTQYF that matches public TCR sequences in VDJdb24, which have been verified to interact with the HLA-B*08:01-restricted IE-1 epitope (QIKVRVDMV)25. Similarly, the consensus sequence CAxxxxGAYNEQFF also matches a public CDR3β sequence against the same IE-1 epitope (QIKVRVDMV) restricted by HLA-B*08:0126. We also identified a clonotype with the CDR3β sequence CASSYSGNTEAFF, which has been previously shown to interact with the HLA-B*07:02-restricted pp65 epitope (TPRVTGGGAM)27,28. Furthermore, several consensus CDR3β sequences are shared by both “durable” and “transient” clonotypes (Fig. 4f). Taken together, our results show that V160 elicits highly polyclonal T cells.
V160 induces durable cytotoxic CD8 T cell responseAfter the vaccine-reactive T cell clonotypes were identified, we sought to better understand these T cells. We annotated the T cells in the scRNA-seq/scTCR-seq data with vaccine-responsive status that were determined from longitudinal TCR profiling by using the CDR3β nucleotide sequences as natural barcodes. By integrating our datasets this way, we can now compare the “durable” vs. “transient” T cells in the single-cell expression data. About 6.45% (45 out of 697) “transient” clones and 48.98% (96 out of 196) “durable” clones matched to scRNA-seq/scTCR-seq data, in which the percentage of responsive CD8 T cells is predominantly higher than responsive CD4 T cells (Fig. 4g). Additionally, V160 responsive CD8 T cells were mostly enriched for “durable” clones, whereas V160 responsive CD4 T cells were mostly enriched for “transient” clones (Fig. 4h).
To validate the results from scRNA-seq and determine the durability of virus-specific T cell responses induced by V160, we did a side-by-side ICS flow cytometry experiment to compare the percentages of IE-1 and pp65 responding CD4 and CD8 T cells in month 9 and month 18 PBMCs from the same subject, with 5 subjects from the 2-dose group and 5 subjects from the 3-dose group. Individual responses were shown in Supplementary Fig. 6. A predominantly higher percentage of antigen-specific CD8 T cells were detected than CD4 T cells in both month 9 and month 18 PBMCs, which is consistent with our scRNA-seq analysis. Interestingly, the average percentages of IE-1 and pp65 responding CD4 and CD8 T cells in the 2-dose group remained stable or showed an upward trend from months 9 to 18. The average percentages of IE-1 and pp65 responding CD4 and CD8 T cells in the 3-dose group had an obvious drop at month 18, mainly due to two high responders (S26 and S28), but still remained at a comparable level to the 2-dose group (Fig. 4i, Supplementary Fig. 6). Notably, one subject (S23) in the placebo group showed dramatically increased T cell responses from month 9 to 18. The titers of HCMV antigen-specific IgG in serum samples of this subject also significantly increased from months 9 to 18 (Supplementary Fig. 7), which indicates HCMV infection. The combinatorial analysis demonstrated that the proportions of total IE-1 responding CD8 T cells and pp65 responding CD4 T and CD8 T cells that co-expressed ≥2 effector molecules at month 18 are comparable to those at month 9 (Fig. 4j, Supplementary Fig. 8). Collectively, these results demonstrate that V160-induced cytotoxic T-cell responses are durable from month 9 to month 18 with a preserved polyfunctionality.
V160-induced CD8 T cells exhibit a dominant effector phenotypeTo better understand the vaccine-responsive “durable” and “transient” clones, we identified the location of these cells in the T-distributed stochastic neighborhood embedding (TSNE) transcriptomic map (Fig. 5a). We found that the “durable” clones are predominantly found in effector cytotoxic T cell clusters 7 and 11, whereas the “transient” clones are predominantly found in the CD4 T cells clusters 5 and 9 (Fig. 5b). Based on single-cell transcriptomic profiles, these vaccine-responsive CD8 T cells were predominantly effector memory T cells of the TEMRA subset, especially for the “durable” clones (Fig. 5c). Furthermore, differential expression analysis of “durable” vs. non-responsive T cells revealed that “durable” clones greatly upregulate many effector-function related chemokines and cytokines such as CCL4, XCL1, XCL2, CCL4L2, and IFNG, as well as cytotoxic protein GZMB (Fig. 5d, Supplementary data file 3). In addition, the “durable” clones also upregulate transcription factors T-bet (TBX21) and eomesodermin (EOMES) and the fractalkine-binding chemokine receptor CX3CR1, which are also expressed by CD8 effector memory T cells induced by HCMV infection in humans29. Conversely, the “transient” vs. non-responsive T cells revealed that the “transient” clones upregulate modulators such as Annexin A1 (ANXA1) and Annexin A2 (ANXA2), as well as adhesion molecules such as Vimentin (VIM), LGALS1 (Galectin 1) and LGALS3 (Galectin 3) (Fig. 5e, Supplementary data file 4).
Fig. 5: V160-responsive “durable” expanded T cells were dominated by a TEMRA phenotypes.
a Highlighting of responsive T cells with “transient” or “durable” expansion on the TSNE plot as in Fig. 2a. Statistical significance of the enrichment of each cluster for transient or durable clones was assessed by Fisher’s exact test. b Distributions of V160-responsive “transient” vs. “durable” expanded clones across the expression clusters. c Distributions of T memory subsets among V160-responsive “transient” or “durable” expanded clones of CD8 T cells, as determined based on single-cell expression profiles. Error bars in b and c indicate mean ± standard error of the mean. d and e Volcano plots of scRNA-seq differential gene expression analyses of responsive “durable” T cells versus non-responsive T cells (d), or responsive “transient” T cells versus non-responsive T cells (e). Each dot summarizes the result for a gene. FDR: false discovery rate. FC fold change. f Flow cytometry analysis of memory T cell subsets among IE-1 or pp65 specific CD8 T cells in months 9 and 18 PBMCs of subject S26 and in an HCMV+ donor. Antigen-specific T cells identified by IFN-γ expression are shown in red and overlaid with total CD8 T cells shown in gray. g Group average distributions of memory T-cell subsets among IE-1 or pp65 specific CD8 T cells in month 9 or 18 PBMCs from 2-dose (n = 5), 3-dose (n = 5), or HCMV+ (n = 7) groups. Group average percentage of IFN-γ+ CD8 T cells was shown in the center of the pie chart. Individual-level data of memory T cell subset distributions among IFN-γ+ CD8 T cells, IL-2+ CD8 T cells, TNF-α+ CD8 T cells, and CD107a+ CD8 T cells are shown in Supplementary Fig. 9.
We then validated the effector phenotype of the vaccine-responsive cytotoxic T cells using a flow cytometry assay. By including two additional cell surface markers (CCR7 and CD45RO) into the ICS flow cytometry assay30, we were able to differentiate the antigen-specific CD8 T cells into four subsets, the CD45RO−CCR7+ naïve T cells (naïve), CD45RO+CCR7+ central memory T cells (TCM), CD45RO+CCR7− effector memory T cells (TEM) and the CD45RO−CCR7− subset which corresponds to the TEMRA cells (Supplementary Fig. 1). We analyzed the distributions of four subsets among IE-1 or pp65 specific IFN-γ+, CD107A+, IL-2+, and TNF-α+ CD8 T cells in month 9 and month 18 PBMCs from the same subjects and compared to those in HCMV+ donors. For S26 in the 3-dose group, TEMRA phenotype accounts for 89.7% of the 3.4% IE-1 specific IFN-γ+CD8 T cells at month 9 and 88.2% of the 2.6% IE-1 specific IFN-γ+CD8 T cells at month 18; TEMRA phenotype accounts for 80.4% of the 1.5% pp65 specific IFN-γ+CD8 T cells at month 9 and 66.7% of the 0.8% pp65 specific IFN-γ+CD8 T cells at month 18 (Fig. 5f). The dominance of TEMRA phenotype among IFN-γ+CD8 T cells in S26 is consistent with our single cell analysis data. The average proportions of TEMRA phenotype in both 2-dose and 3-dose groups remain stable from month 9 to month 18, though the 2-dose group has lower proportions of TEMRA phenotype and higher proportions of TEM phenotype than the 3-dose group (Fig. 5g). In contrast, the TEMRA and TEM phenotypes together dominate the 1.5% IE-1 IFN-γ+CD8 T cells and 0.2% pp65 responding IFN-γ+CD8 T cells in HCMV+ donors, which are different to those in the 3-dose group but similar to the 2-dose group (Fig. 5f, g, supplementary Fig. 9). Notably, the proportions of total naïve CD8 T cells showed no significant change from month 9 to 18 and are significantly higher compared to HCMV+ donors (Supplementary Fig. 10), suggesting minimal perturbation of immune system by V160 vaccination when compared to natural HCMV infection. Together, these results demonstrate that V160-induced CD8 T cells comprise a stable population of TEMRA phenotype that persists for at least one year after the last dose of V160 vaccination.
Transcriptional signatures of durable CD8 T cells post V160 vaccinationWe next sought to determine the transcriptional signatures of V160 responsive “durable” CD8 T cells. We compared the single-cell transcriptomes of “durable” versus “transient” CD8 T cells (Fig. 6a). Compared to “transient” CD8 T cells, the “durable” CD8 T cells upregulated the expression of chemokines such as CCL4 and CCL5, which attract immune cells to the site of infection, as well as cytotoxic protein marker NKG7. The “durable” CD8 T cells also expressed higher levels of glycolysis-related enzymes such as PGAM1, PKM, and GAPDH (Fig. 6a, Supplementary data file 5). This preferential use of glycolysis for energy generation is a metabolism feature of effector CD8 T cells31. Gene set enrichment analysis showed that the “durable” CD8 T cells activated IL2-STAT5 signaling, which promotes differentiation into terminal effector cytolytic T cells32,33, as well as mTORC1 signaling, which is essential for regulation of T-cell glycolytic metabolism34. Furthermore, “transient” CD8 T cells activated TNF-α signaling via NF-κB which exerts potent pro-inflammatory functions, while “durable” CD8 T cells showed even higher levels of TNF-α signaling via NF-κB but significantly suppressed interferon-α response (Fig. 6b, c). This could be explained by a cross-regulating relationship between TNF and type I interferons, where TNF controls type I interferons under steady-state conditions35,36.
Fig. 6: Transcriptional signature of V160-responsive “durable” expanded CD8 T cells.
a Left, Volcano plots of scRNA-seq differential expression analysis of “durable” vs. “transient” expanded CD8 T cells. Right, Breakdown of the statistical significance of differential expression into the discrete component (expressed vs. non-expressed) and continuous component (changes in expression level given that the gene is expressed). Each dot summarizes the result for a gene. FDR: false discovery rate. FC: fold change. b Summary of gene set enrichment analyses for responsive “transient” vs. non-responsive, responsive “durable” vs. non-responsive, and responsive “durable” vs. “transient” expanded CD8 T cells. NES, normalized enrichment score. c Gene set enrichment plots for selected significant pathways in the differential expression of “durable” vs. “transient” expanded CD8 T cells. Dashed lines represent minimum and maximum cumulative enrichment scores. d Uniform manifold approximation (UMAP) plots of T cells from expression clusters enriched for “transient” or “durable” expanded V160-responsive clones identified in Fig. 4b. Cells are colored by V160 response type (top) or T cell type (bottom). e Left, Volcano plots of scRNA-seq differential expression analysis of V160 responsive “durable” CD8 T cells subcluster C vs. B in (d). Right, Breakdown down the statistical significance of differential expression into the discrete and continuous components. f Summary of gene set enrichment analyses for V160 responsive “durable” CD8 T cells subcluster C vs. all other CD8 T cells; subcluster B vs. all other CD8 T cells; or subcluster C vs. B. NES, normalized enrichment score. g Percentages of cells expressing selected genes within each V160 responsive T cell cluster. Significance was assessed by Fisher’s exact test. Error bars indicate the mean ± standard error of the mean. h Violin plots showing the distributions of log single-cell expression counts in each response group. Each filled dot represents the log expression of a cell expressing the gene. Open diamonds represent mean log expression. Significance was determined by the Kruskal–Wallis test.
To characterize the relationships among vaccine-responsive T cells more precisely, we performed a UMAP analysis of vaccine-responsive T cells along with T cells that belonged to the same previously determined single-cell expression clusters. This analysis revealed that the V160 responsive “durable” T cells consisted of two subpopulations that we denote as subcluster C and B (Fig. 6d), which correspond to original clusters 11 and 7. We then compared the transcriptional profiles of “durable” CD8 T cells from subcluster C versus B (Fig. 6e, Supplementary data file 6). Subcluster C cells had a dramatic increase in expression of chemokines and cytokines found in activated CD8 T cells, such as XCL1, XCL2, CCL4L2, CCL4, and IFNG, whereas these genes were not expressed in subcluster B (Fig. 6e, g). Consistently, subcluster C also upregulated the expression of glycolysis-related proteins, including PGAM1, PKM, and GAPDH, as well as cytotoxic marker GZMB (Fig. 6e). We speculate that these differences between subclusters C and B should represent V160 responsive cells with or without in vitro stimulation, as IE-1 and pp65 only contain a fraction of potential V160 T-cell epitopes. At the pathway level, “durable” CD8 T cells from subclusters B and C both activated TNF-α signaling via NF-κB and suppressed interferon-α response, whereas only subcluster C cells showed strong activations of many additional pathways such as mTORC1 signaling and IL2-STAT5 signaling (Fig. 6f), suggesting that TNF-α signaling via NF-κB may play a role in the maintenance of “durable” CD8 T cells induced by V160. Overall, “durable” CD8 T cells from subcluster C appeared to show the highest transcript expression of many inflammatory genes, whereas “durable” CD8 T cells from subcluster B showed intermediate expression levels, and “transient” CD8 T cells showed the lowest levels (Fig. 6g, h). Taken together, our results demonstrate that the “transient” and “durable” CD8 T cells induced by V160 vaccination activated distinctive transcriptional programs.
留言 (0)