
記住我
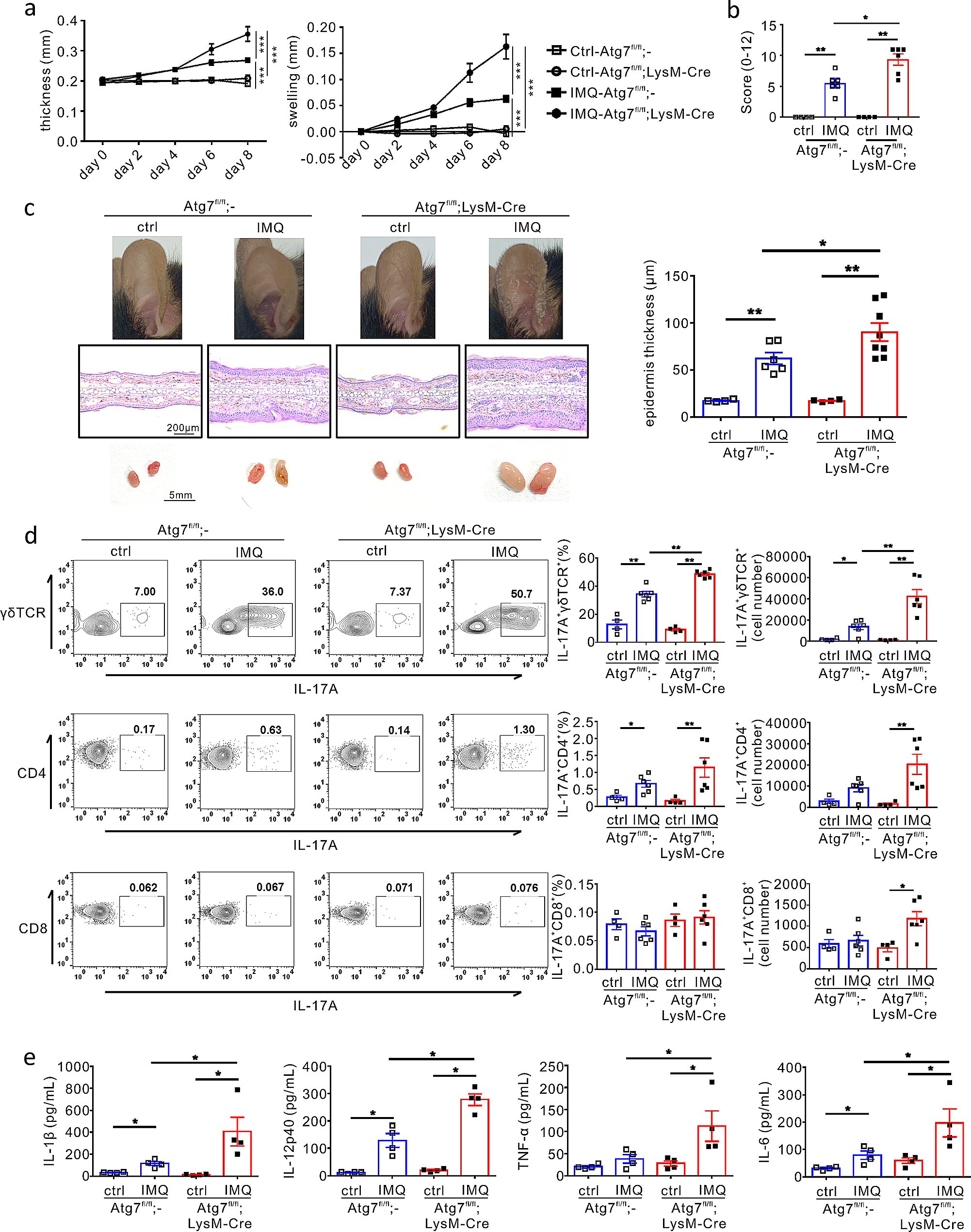





Here we summarize the individual lncRNAs that are involved in ocular angiogenesis, with more emphasis on those with in vivo functional studies. They are classified into pro-angiogenic and anti-angiogenic lncRNAs.
Proangiogenic lncRNAs (Table 2)Table 2 Pro-angiogenic lncRNAsANRILAntisense RNA to INK4 locus (ANRIL) is a significant lncRNA in cardiovascular diseases, diabetes, and cancer. ANRIL’s single nucleotide polymorphism associates with type II diabetes and coronary artery diseases [56]. It is detectable in human and mouse RECs and upregulated under HG conditions [57]. Also, it’s elevated in the retina of STZ-induced DR rats and in the serum and vitreous of DR patients [53, 58]. ANRIL silencing using siRNAs prevented HG-induced VEGF expression and vascular tube formation in hRECs, and ANRIL−/− mice are phenotypically normal but are protected from diabetes-induced VEGF upregulation [59]. Conversely, overexpression of ANRIL upregulates VEGF and promotes angiogenesis [60]. Mechanistically, ANRIL binds to both p300 and EZH2 of the Polycomb Repressive Complex (PRC) 2 to regulate VEGF expression. The upregulation of miR200b in si-ANRIL-transfected hRECs suggests the involvement of mir200b in the ANRIL-regulated VEGF expression. Combined with the data that VEGF is a target gene of miR-200b, and PRC2 represses miR-200b, ANRIL likely regulates VEGF expression by interacting with p300 and EZH2 in the PRC2 complex, which in turn represses miR-200b expression (Fig. 3) [61, 62]. In an independent study, the infection of si-ANRIL into STZ-induced DR rats reversed DR pathologies [58]. Downregulation of the NF-k pathway by si-ANRIL in the retina was confirmed by reduced P65 phosphorylation and the mRNA level of inflammatory markers (IL-1, IL-10, and MCP-1). In another relevant study, adenovirus mediated ANRIL overexpression increased Akt and eNOS [63]. These establish ANRIL as a key pro-angiogenic and pro-inflammatory lncRNA with implications in DR (Fig. 3).
Fig. 3
ANRIL lncRNA Promotes inflammation by regulating NF-kB pathway and enhances VEGF expression and angiogenesis through interacting with p300 and EZH2, which regulates miR200b. The target sequence in VEGFA that is complementary to miR200b seed region is CAGUAUU (1314–1320) in VEGFA 3’UTR
HOTAIRHOX antisense intergenic RNA (HOTAIR) was initially identified for its role in cancer progression and metastasis [64, 65]. It is highly conserved in mouse, rat and human. Its expression is upregulated in human RECs under HG condition, in the retina of STZ-induced DR mice, and in the vitreous humor and serum from proliferative DR patients [66, 67]. HOTAIR knockdown repressed REC proliferation and migration, and vessel-like structure formation under HG condition in vitro. Delivering of HOTAIR shRNA by AAV virus or HOTAIR siRNA alleviated retinal vessel impairment in STZ-induced DR mice and rats, as shown by decreased acellular capillaries, increased pericytes in the retina, and decreased retinal vascular leakage. Mechanistically, HOTAIR silencing represses the expression of multiple RNA transcripts implicated in angiogenesis, such as VEGFA, ANGPTL4, PGF (placental growth factor), and HIF1α. HOTAIR has been shown to regulate target gene transcription by binding to histone methylase PRC2 and histone demethylase LSD1 [68]. HOTAIR also binds to histone demethylase LSD1 and HIF1, or recruits RNA polymerase II and histone methyltrasferase EZH2 and acetylators (P300) to VEGF promoter and enhance its transcription. HOTAIR also binds to LSD1 to inhibit the H3K4me3 on the VE-cadherin promoter, thereby suppressing VE-cadherin transcription. Therefore, HOTAIR is a proangiogenic lncRNA with potential to regulate angiogenesis and vascular permeability in DR. Overall, HOTAIR functions as a scaffold for transcription factors that increase expression of pro-angiogenic genes (Fig. 4).
Fig. 4
HOTAIR lncRNA promotes angiogenesis by binding to PRC2 and LSD, which regulates VEGF-A, ANGPTL4, PFGF and HIF1α transcription
HOTTIP(HOXA transcript at the distal tip) (HOTTIP), located as the distal 5’ tip of the HOXA locus, is a critical oncogenic lncRNA [69]. It functions as an important mediator of chromatin activation by recruiting WD repeat-containing protein (WDR5) and mixed lineage leukemia (MLL) (Fig. 5). HOTTIP expression is significantly upregulated in STZ-induced DR mice and rats. Adenovirus-delivered shRNA against HOTTIP alleviated diabetes-induced visual function decline and apoptosis of retinal cells and reduced the expression of ICAM-1 and VEGF in the retina. Mechanistically, it functions by activating the p38-MAPK pathway in retinal ECs [70]. Its underlying mechanism in DR is yet to be discovered.
Fig. 5
HOTTIP lncRNA promotes inflammation and angiogenesis by activating p38/MAPK pathway, as well as by interacting with WDR5 and MLL, which regulates ICAM-1 and VEGF transcription
H19H19, one of the early discovered lncRNAs, is maternally expressed in the H19-Igf2 locus and overexpressed in multiple cancers [71, 72]. A differentially methylated H19 upstream region determines the reciprocal expression of H19 from the maternal allele and Igf2 from the paternal allele. As a lncRNA, H19 also serves as a source of miR-675, which restricts Igf1r expression. Additionally, H19 acts as ceRNA for multiple miRNAs, being linked to multiple pathological processes, including inflammation, angiogenesis, neurogenesis, and fibrosis progression [71, 73, 74]. Regarding ocular angiogenesis, H19 is markedly upregulated in the vascularized corneal and can be induced by bFGF in ECs. It promotes EC angiogenesis by sponging miR-29c that targets VEGFA [75]. In the context of DR, downregulation of H19 was observed in vitreous humor samples from individuals with DR, in the retina of diabetic mouse models and in HG-induced hREC and ARPE-19 cells [76,77,78]. Conversely, Fawzy et al. observed H19 upregulation in the plasma of diabetic patients compared to healthy subjects, with no significant differences between patients with and without DR [79]. H19 overexpression prevented HG-induced endothelial–mesenchymal transition (endMT), while its silencing led to endMT phenotypes similar to HG induction. In H19−/− mice, endMT phenotypes, shown by reduced Cd31 and Ve-Cad expression, increased Fsp1 and α-SMA expression, and extravascular IgG staining, were also observed. Mechanistically, H19 represses endMT by suppressing TGF-β1 and by repressing the MAPK–ERK1/2 signaling pathway. Beyond endMT, H19 also inhibits HG-induced expression of inflammatory cytokines, such as TNF-α, IL-1β and IL-6 in ARPE-19 cells, by sponging miR-19b which targets silence information regulator factor related enzymes 1 (SIRT1). These studies suggest H19 is a proangiogenic lncRNA but can inhibit multiple pathological processes, including endMT and inflammation, in DR models (Fig. 6). Further in vivo studies are needed to test the role of H19 in ocular angiogenesis, especially proliferative DR or neovascular AMD.
Fig. 6
H19 lncRNA Inhibits endMT by inhibiting TGF-β1 and MAPK-ERK1/2 pathway; inhibits inflammation by sponging miR-19b that targets SIRT1; enhances angiogenesis by sponging miR-29c that targets VEGF-A. The target sequence in SIRT1 that is complementary to miR-19-3p seed region is UUGCAC (1285–1291) in SIRT1 3’UTR. The target sequence in VEGFA that is complementary to miR-29c seed region is UGGUGCUA (1758–1765) in VEGFA 3’UTR
IPWIPW (Imprinted gene in the Prader-Willi syndrome (PWS) region) was initially identified in an epigenetic PWS disorder [80]. Located in the critical region of the PWS locus, IPW regulates the imprinted DLK1-DIO3 region [81]. Disruption of the IPW region is associated with neurogenic disorders in humans. IPW was found to be significantly up-regulated in the choroids in laser-induced CNV mouse model and in hypoxic ECs [82]. IPW silencing or overexpression reduced or increased EC viability, proliferation, migration, and tube formation in vitro, as well as choroid sprouting angiogenesis ex vivo. IPW silencing also inhibited laser-induced CNV in vivo. Mechanistically, IPW silencing caused upregulation of miR-370 but not the miR-409 and MEG3 genes in the DLK1-DIO3 locus. miR-370 has been shown to inhibit angiogenesis by targeting KDR and BMP-2 [82, 83]. IPW overexpression rescued the anti-angiogenic effect of miR-370 in the ex vivo sprouting angiogenesis model and in vivo laser-induced CNV model. These suggest IPW as a pro-angiogenic lncRNA that functions by repressing miR-370, and IPW silencing could be a promising strategy for treating neovascular ocular diseases (Fig. 7).
Fig. 7
IPW lncRNA enhances angiogenesis by repressing miR-370 at the DLK1-DIO3 locus, which targets KDR and BMP-2. The target sequence in KDR that is complementary to miR-370 seed region is CAGCAGG (79–86) in KDR 3’UTR. The target sequence in BMP-2 that is complementary to miR-370 seed region is AGCAGG (725–731) in BMP-2 3’UTR
MALAT1Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) was initially discovered as a predictor for lung cancer metastasis [84]. In an OIR murine model and HG-stimulated hRECs, MALAT1 was upregulated together with VEGF and HIF1a, while miR-203a-3p was downregulated. MALAT1 was also upregulated in the aqueous humor samples, and in fibrovascular membranes of diabetic patients [48]. Knockdown of MALAT1 blunted HG-induced proliferation, migration, and tube-like structure formation in human retinal microvascular ECs (hRMEC), mouse RMEC and HUVEC cells [39, 85,86,87,88]. Mechanistically, MALAT1 was found by several studies to function as ceRNA to sponge multiple miRNAs, including miR-203a-3p, miR-200b, miR-205-5p, miR-124-3p and miR-320a, which relieve the repression of angiogenic genes VEGF, HIF1 and EGR1 by these miRNAs. Genetic ablation of MALAT1 reduced EC proliferation and neonatal retina vascularization [89]. GapmeRs-induced pharmacological inhibition of MALAT1 led to decreased blood flow recovery and capillary density in a hindlimb ischemia model. In STZ-induced diabetic rats, MALAT1 knockdown by shRNA injection ameliorated DR phenotypes in vivo, including pericyte loss, capillary degeneration, microvascular leakage, and retinal inflammation [39]. In addition, inhibiting retinal MALAT1 alleviated retinal neurodegeneration in the STZ mouse model [90]. In the OIR mouse model, silencing of MALAT1 by intravitreal siRNA injection inhibited retinal neovascularization in vivo, shown by reduced neovascular tufts, non-perfusion region, and EC nuclear counts [91]. The expression of CCN1, phosphor-AKT, and VEGF, which was upregulated by hyperoxia in the retina, was shown to be downregulated by si-MALAT1. In addition, the expression of inflammatory cytokines IL-1β, IL-6, and TNF- α were reduced by MALAT1 silencing. These suggest that MALAT1 could sponge multiple miRNAs to promote angiogenesis and potentially inflammation. Consequently, MALAT1 may serve as a potential therapeutic target for DR and ROP (Fig. 8).
Fig. 8
MALAT1 lncRNA promotes angiogenesis by acting as ceRNA for miR203a-3p, miR-205-5p, miR-124-3p, and miR-320 that target VEGF, HIF1α and EGR1, and promotes inflammation by elevating IL-1β, IL-6 and TNF-α levels. The target sequences in HIF1α that is complementary to miR-320a and miR-203a seed region are AGCUUU (1846–1852) and AUUUCA (995–1001) in HIF1α 3’UTR. The target sequences in VEGFA that is complementary to miR-205-5p and miR-203-3p seed regions are AUGAAGG (167–173) and AUUUCA (1320–1326) in VEGFA 3’UTR. The target sequence in EGR1 that is complementary to miR-124-3p seed region is GUGCCUU (769–775) in EGR1 3’UTR
MIATMIAT was initially discovered as a myocardial infarction (MI)-associated transcript (MIAT) [92]. Its expression was shown to be upregulated in the plasma of DR patients compared to both patients without DR and healthy individuals [93]. It is also upregulated by HG in different cell lines, and in the retina of diabetic rats and STZ-treated mice [94,95,96]. MIAT knockdown significantly reduced EC proliferation but increased EC cell death under HG. It also inhibited EC migration and tube-like structure formation induced by TNF-α or VEGF. Silencing MIAT in STZ-induced diabetic rats alleviated diabetic-induced retinal neovascularization, vascular leakage, and inflammation in vivo. Mechanistically, MIAT enhances the expression of VEGF by acting as a miR-150-5p sponge. Under hyperglycemia or hypoxic stress that induces angiogenesis, MIAT is upregulated, relieving the miR-150-5p associated repression of VEGF [94]. Consistently, endothelial miR-150 has been shown to be an intrinsic suppressor of pathological ocular neovascularization [97]. These findings established MIAT as an endogenous miR-150-5p sponge, driving the stimulation of ocular angiogenesis (Fig. 9).
Fig. 9
MIAT lncRNA promotes angiogenesis by acting as ceRNA for mirR-150-5p that targets VEGF. The target sequences in VEGFA that is complementary to miR-150-5p seed region are UUGGGA (676–681) and GGGGA (789–794) in VEGFA 3’UTR
NEAT1Nuclear-enriched abundant transcript 1 (NE AT1), also known as nuclear paraspeckle Assembly Transcript 1, is an abundant, ubiquitously expressed lncRNA, which forms a scaffold for a specific RNA granule in the nucleus, the paraspeckle. It has been implicated in multiple neurodegenerative diseases and other diseases [98, 99]. Contrasting results were observed regarding the expression of NEAT1 in DR. While downregulated in the retinal of STZ-treated mice, but upregulated in the serum of DR patients, it displays both up- or down-regulation in HG-treated retinal cells [100,101,102]. NEAT1 is upregulated in laser-induced CNV and M2 macrophage [45]. NEAT1 silencing in hRECs ameliorated HG-triggered apoptosis, oxidative stress, and inflammation through inactivating TGF-β1 and VEGF-A. Intravitreal injection of NEAT1 smart silencer inhibited laser-induced CNV leakage and neovascularization. Mechanistically, NEAT1 acts as sponge to miRNA-148a-3p to regulate the expression of PTEN, which prevents resolving inflammation by inhibiting M1 to M2 macrophage polarization. In another study, NEAT1 was shown to promote gastric cancer cell angiogenesis by enhancing proliferation, migration, and tube-like structure formation ability of ECs, by sponging miR-17-5p which targets TGFβR2 directly, which in turn, upregulates angiogenic factors including VEGF-A [103]. Therefore, NEAT1 lncRNA could promote ocular angiogenesis as a ceRNA for several miRNAs miR-148-3p and miR-17-5p to regulate PTEN and TGFβR2 (Fig. 10).
Fig. 10
NEAT1 lncRNA enhances angiogenesis by acting as ceRNA for miR-19-5p that targets TGB-βR2 and therefore enhancing VEGF-A expression. It also prevents resolving inflammation by inhibiting M2 macrophage polarization by acting ceRNA for miRNA-148a-3p that targets PTEN. The target sequences in PTEN that is complementary to miR-148-3p seed region are UGCACUG (2254–2260) and UGCACUG (3151–3158) in PTEN 3’UTR. The target sequence in TGF-βR2 that is complementary to miR-17-5p seed region is GCACUUU (268–275) in TGF-βR2 3’UTR
TUG1Taurine upregulated 1 (TUG1) was established as an oncogenic or tumor suppressor lncRNA by different studies [104, 105]. TUG1 knockdown suppressed tumorigenesis and tumor-induced angiogenesis in mouse hepatoblastoma or glioblastoma xenograft models [106, 107]. Further studies showed that TUG1 functions a ceRNA for multiple miRNAs, including mitigating the function of miR-34a-5p and miR-299 in repressing VEGFA expression, the function of miR-204-5p in repressing JAK2/STAT3 pathway, the function of miR-145-5p in repressing CCN1 expression, and the function of miR-29c-3p in regulating PDGF-BB/Wnt signaling [106,107,108,109,110,111]. In a diabetic limb ischemia mouse model, injection of TUG1 lentivirus stimulated angiogenesis [111]. In the mouse OIR model, intravitreal injection lentivirus expressing TUG1 shRNA reduced avascular and neovascular areas and reduced inflammation shown by reduced expression of the inflammatory factors IL-1β, IL-6, and TNF-α in the retina. These suggest that TUG1 silencing could hold a beneficial role in addressing retinal vascular diseases (Fig. 11).
Fig. 11
TUG lncRNA enhances angiogenesis by acting as ceRNA for miR-34a-5p and miR-299 that targets VEGF, miR-204-5p that targets JAK2, miR-145-5p that targets CCN1 and miR-29c-3p that targets PDGF-BB. It also promotes inflammation by elevating TNF-α, IL-1β and IL-6. The target sequences in VEGFA that is complementary to miR-34-5p and miR-299-3p seed regions are CACUGCC (859–865) and CCCACAU in VEGFA 3’UTR. The target sequence in JAK2 that is complementary to miR-204-5p seed region is AAGGGA (1453–1459) in JAK2 3’UTR. The target sequence in CCN1 that is complementary to miR-145-5p seed region is ACUGGA (572–577) in CCN1 3’UTR. The target sequence in PDGFB that is complementary to miR-29c-3p seed region is UGGUGCU (132–138) in PDGFB 3’UTR
Anti-angiogenic lncRNAs (Table 3)Table 3 Anti-angiogenic lncRNAsMEG3Maternally expressed gene 3 (MEG3) is an imprinted lncRNA gene that is expressed in both humans and mice. In many human tumor and tumor cell lines, its expression is lost, indicating that it has a tumor supressive role. MEG3 has anti-proliferative and pro-apoptotic activities partly through stimulating P53 accumulation [112, 113]. MEG3 expression is downregulated in the serum of diabetic and DR patients, STZ-induced diabetic mice, and in RECs under HG conditions. MEG3 overexpression inhibited EC proliferation, survival, and networking, while MEG3 knockdown had the opposite effect, suggesting an anti-angiogenic function of the lncRNA [114]. However, a positive role for MEG3 in VEGF-induced angiogenesis was also reported [115]. Maternal deletion of the Meg3 gene in mice results in skeletal muscle defects and perinatal death [112]. Upregulation of VEGFA gene and genes in VEGF pathways were detected in Meg3−/− embryos, consistent with increased cortical microvascular density in the knockout embryos. MEG3 knockdown by specific shRNAs aggravated retinal vessel dysfunction in vivo, shown by serious capillary degeneration, increased microvascular leakage, and inflammation. Mechanistically, MEG3 knockdown enhances PI3K/AKT signaling, while MEG3 overexpression represses high glucose-induced TGF-β and VEGF expression [114,
留言 (0)