
記住我



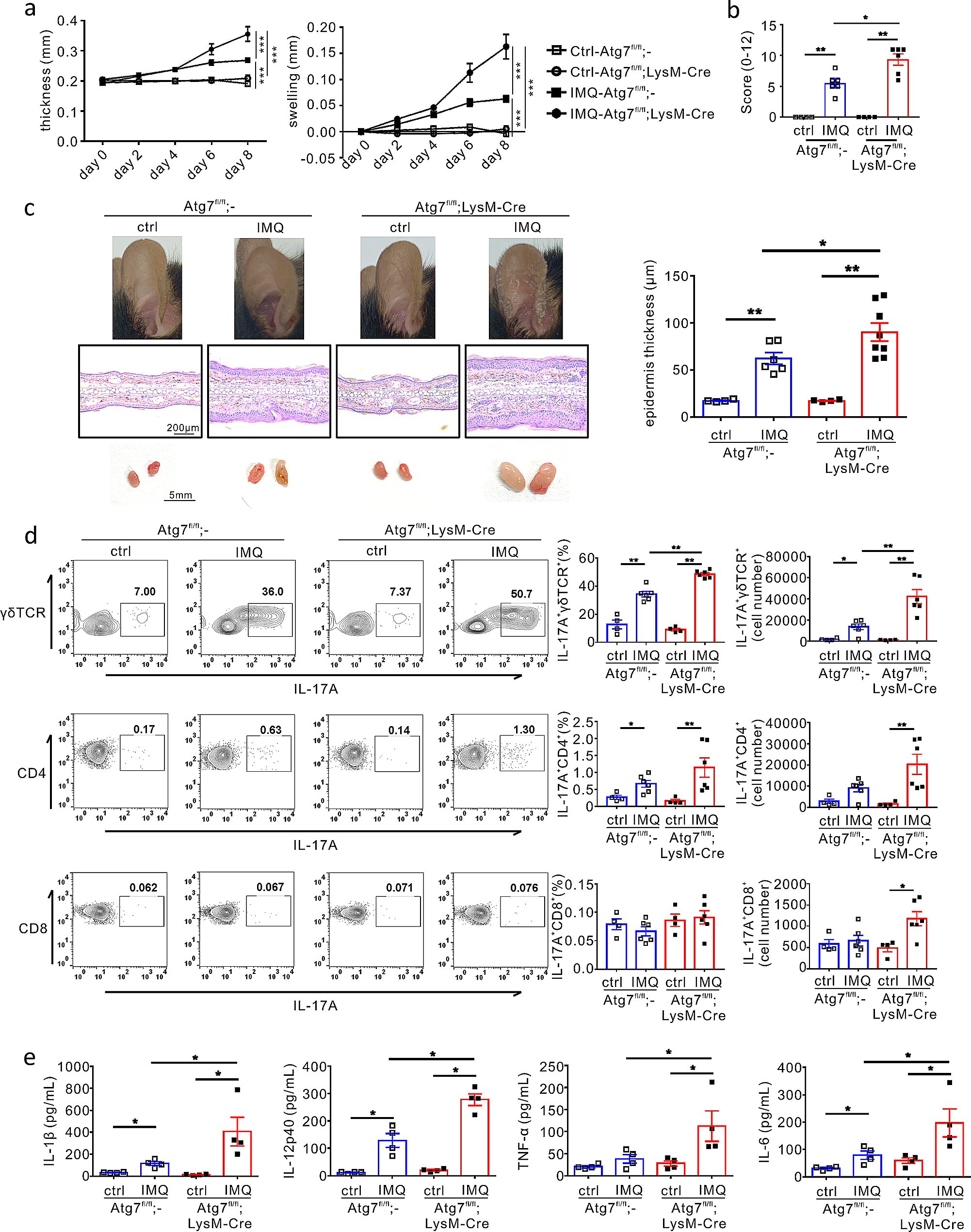



To explore the direct role of autophagy in myeloid cells in the pathogenesis of psoriasis, we developed a model of psoriasis using Atg7fl/fl;LysM-Cre mice, which selectively exhibit a deficiency of the autophagy gene Atg7 in myeloid cells such as neutrophils and macrophages [45]. Imiquimod (IMQ) cream was topically applied daily on the ears of the mice over 8 days, and skin inflammation was assessed by measuring the ear thickness every other day. Ear thickness and swelling were significantly increased upon IMQ application in both Atg7fl/fl;LysM-Cre and Atg7fl/fl mice; however, this increase was more significantly pronounced in the Atg7fl/fl;LysM-Cre mice than in the Atg7fl/fl mice (Fig. 1a). Consistent with this result, the Atg7fl/fl;LysM-Cre mice showed a more severe IMQ-induced PASI score and inflammatory response in the ears than the Atg7fl/fl mice (Fig. 1b and c upper), along with significantly aggravated epidermis thickness (Fig. 1c, middle and right). In addition, IMQ-induced swelling of skin-draining lymph nodes (dLNs) was more pronounced in the Atg7fl/fl;LysM-Cre mice than in the Atg7fl/fl mice (Fig. 1c bottom). These results suggest that autophagy in myeloid cells plays a protective role in the pathogenesis of psoriasis.
Fig. 1
Effect of impaired autophagy in myeloid cells on psoriatic skin inflammation. An IMQ-induced psoriasis model was established in Atg7fl/fl and Atg7fl/fl;LysM-Cre mice. The mice were sacrificed at the end of the experiment, and tissues were harvested for each analysis. (a) Ear thickness and swelling were measured every other day (n = 6–8 per group) ***P < 0.001, two-way ANOVA. (b) Cumulative clinical score (erythema, scaling, and thickness) on a scale from 0 to 12. All bars indicate the mean ± SEM. *P < 0.05 and **P < 0.01, Mann–Whitney U-test. (c) Representative ear images (top), H&E-stained histological images (middle), and draining lymph nodes (bottom) from each mouse group. The statistical bar chart shows the epidermal thickness of the ear tissue. All dots indicate the number of samples. All bars indicate the mean ± SEM. *P < 0.05 and **P < 0.01, Mann–Whitney U-test. (d) Flow cytometric analysis of the indicated IL-17 A-producing T cells in the draining lymph nodes. The number of samples is indicated by dots. All bars indicate mean ± SEM. *P < 0.05 and **P < 0.01, Mann–Whitney U-test. (e) The levels of IL-1β, IL-12p40, IL-6, and TNF-α in harvested ear tissues from each group of mice, as determined using ELISA. All dots indicate the number of samples. All bars indicate mean ± SEM. *P < 0.05 and **P < 0.01, Mann–Whitney U-test
Considering the critical role of γδ T cells and T helper 17 cells (Th17 cells) in the pathogenesis of psoriasis [21], we assessed the effect of autophagy deficiency in myeloid cells on T cells isolated from dLNs in Atg7fl/fl;LysM-Cre or Atg7fl/fl mice using flow cytometry. γδ T cells are the major cellular source of IL-17 A in the IMQ-induced mouse model of psoriasis [18]. In both groups of mice, IMQ application significantly increased the population of IL-17 A+γδ T cells, which was further significantly elevated by autophagy deficiency, in terms of percentage and absolute numbers (Fig. 1d upper). In comparison, IL-17 A+CD4+ T cells populations in the dLNs were clearly increased in both groups upon IMQ application but were not significantly affected by autophagy deficiency (Fig. 1d middle). Similarly, autophagy deficiency exhibited a marginal effect on the population of IL-17 A+CD8+ T cells in dLNs of IMQ-treated mice, with a significant effect only on the absolute number (Fig. 1d bottom).
On the basis of the aforementioned findings regarding the association of autophagy deficiency in myeloid cells with exacerbated skin inflammation and increased populations of IL-17 A-producing T cells, especially γδ T cells, we sought to understand the molecular mechanism underlying this effect. To this end, we examined the production of pro-inflammatory cytokines, which play a crucial role in the induction and differentiation of IL-17 A+γδ T cells and Th17 cells [1, 2]. We observed a significant increase in the production of IL-1β, IL-12p40, and IL-6 in the IMQ-treated ears of mice compared with that in untreated mice, whereas the TNF-α level was only significantly elevated in IMQ-treated Atg7fl/fl;LysM-Cre mice (Fig. 1e). Notably, all these cytokines further increased in the Atg7fl/fl;LysM-Cre mice compared with those in Atg7fl/fl mice (Fig. 1e), indicating that autophagy loss in myeloid cells promoted pro-inflammatory cytokine production, thereby contributing to the increased population of IL-17 A-producing T cells, particularly IL-17 A+γδ T cells. Collectively, these results suggest a protective role of autophagy in myeloid cells in the pathogenesis of skin inflammation in the IMQ-induced mouse model of psoriasis.
IL-1β dysregulation in autophagy-deficient macrophages is associated with neutrophilic inflammationTo further investigate how autophagy exacerbates IMQ-induced psoriatic skin inflammation, we assessed the mRNA expression levels of various cytokines and chemokines in the ears of IMQ-treated mice. Similar to the protein level, the mRNA expression of Il1b was significantly increased following IMQ treatment, and this increase was further augmented by myeloid cell-specific autophagy deficiency (Fig. 2a). In comparison, IMQ treatment significantly increased the mRNA expression of Il12p40, Il6, and Tnfa in the mouse ears; however, autophagy deficiency did not increase the expression further (Fig. 2a). Notably, the mRNA expression of Cxcl2, a critical chemokine for neutrophil recruitment into inflamed tissue [5], was significantly elevated by IMQ treatment and was further increased by myeloid cell-specific autophagy deficiency (Fig. 2a). In comparison, no significant difference was observed in the mRNA expression of Cxcl1, another chemokine for neutrophil recruitment (Supple. Figure 1), and Cxcr2, encoding the cognate receptor for CXCL2 (Fig. 2a). In addition, autophagy deficiency did not affect the mRNA expression of Ccl2 for macrophage recruitment or that of Ccl5 for the recruitment of leukocytes, such as T cells, eosinophils, basophils, monocytes, NK cells, dendritic cells, and mast cells (Suppl. Figure 1). IMQ treatment significantly elevated the gene expression of Il23 and Il1a, which are critical cytokines for the induction and differentiation of IL-17 A+γδ T cells and Th17 cells [1, 2]; however, autophagy deficiency did not further increase their expression (Fig. 2a). Additionally, we observed no significant effect of autophagy deficiency on the expression of diverse cytokines, including IL-1 family cytokines (Il18 and Il33), Th17-cell-inducing TGF-β1 (Tgfb1), pro-inflammatory cytokines (Il12p35 and Ifnb), Th1 cytokine (Ifng), and Th2 cytokine (Il4, Il5, and Il13) (Supple. Figure 1). In addition, despite the involvement of prostaglandins in pathogenic Th17 cell-driven inflammation including psoriasis [46], impaired autophagy did not affect the expression levels of Pgts2, encoding COX-2; mPges-1 (a prostaglandin E2 synthase); and Hpgds (a prostaglandin D2 synthase) in myeloid cells (Supple. Figure 1). Thus, these results raise the possible involvement of IL-1β and CXCL2 dysregulation, induced by autophagy deficiency in myeloid cells, in promoting neutrophilic inflammation. In support, treatment with an antagonist of CXCR2 (SB225002) that blocks the effect of CXCR2 ligand CXCL2 for neutrophil recruitment significantly alleviated ear thickness and swelling in Atg7fl/fl psoriatic mice and, notably, in Atg7fl/fl;LysM-Cre psoriatic mice with myeloid autophagy deficiency (Supple. Figure 2a). This therapeutic efficacy was further validated by a significant decrease in the PASI score, epidermis thickness and swelling of dLNs (Supple. Figure 2b and c).
Fig. 2
Association of myeloid autophagy with IL-1β expression and neutrophilic inflammation. (a) Effect of autophagy deficiency in myeloid cells on the mRNA expression of cytokines related to psoriasis pathogenesis. Relative mRNA levels corresponding to the indicated proteins, as determined using qRT-PCR and normalized to β-actin. All dots indicate the number of samples. All bars indicate the mean ± SEM. *P < 0.05 and **P < 0.01, Mann–Whitney U-test. (b) Representative immunostaining for IL-1β, CD68, or Ly6G in the ear tissue from each mouse group. The bar charts show the IL-1β-positive area colocalized with each cell marker and the number of cells for each cell type. Scale bars = 50 μm. All dots indicate the number of samples. All bars indicate the mean ± SEM. *P < 0.05 and **P < 0.01, Mann–Whitney U-test. (c, d) Effect of impaired autophagy on the levels of IL-1β, IL-12p40, IL-6, and TNF-α in R848-stimulated peritoneal macrophages from Atg7fl/fl or Atg7fl/fl;LysM-Cre mice, as determined using ELISA (c) and qRT-PCR (d). Data are representative of at least three independent experiments conducted with duplicates. All bars indicate the mean ± SD. **P < 0.01, Student’s t-test
Autophagy regulates IL-1β release by degrading inflammasome components such that the loss of autophagy leads to uncontrolled IL-1β production [33,34,35]. In addition, the IL-1β/CXCL2 axis promotes neutrophil recruitment [16]. Thus, we hypothesized that dysregulation of IL-1β production by impaired myeloid autophagy primarily contributes to the recruitment of myeloid cells, including neutrophils, into the inflamed skin lesion. To this end, we first examined the localization of IL-1β in association with surface markers for macrophages and neutrophils in the ears of psoriasis-induced mice. Upon IMQ treatment, IL-1β production was apparently increased from CD68+ macrophages and, to a lesser extent, from Ly6G+ neutrophils and was significantly upregulated by autophagy deficiency (Fig. 2b). The dominant colocalization of IL-1β in CD68+ macrophages rather than Ly6G+ neutrophils suggests that macrophages were the predominant producers of IL-1β linked to autophagy deficiency in our mouse model of psoriasis, consistent with the previous finding that activated monocytes/macrophages are a principal source of IL-1β [47]. Furthermore, the numbers of CD68+ macrophages and Ly6G+ neutrophils in the skin lesions were markedly increased by autophagy deficiency (Fig. 2b), suggesting that myeloid autophagy coordinates both IL-1β production and neutrophilic inflammation.
We examined the direct contribution of macrophage autophagy to the production of inflammatory cytokines such as IL-1β, IL-12p40, IL-6, and TNF-α in vitro. To this end, peritoneal macrophages were isolated and stimulated with R848, a ligand for Toll-like receptors (TLR) 7 and 8. Subsequently, the production of these cytokines in the culture supernatant was measured using ELISA. In activated macrophages, the production of IL-1β, but not that of IL-12p40, IL-6, and TNF-α, was significantly elevated by autophagy deficiency (Fig. 2c). In comparison, the mRNA expression of these cytokines, including Il1b, was not affected by autophagy deficiency in activated macrophages (Fig. 2d), consistent with prior studies on autophagy-mediated targeted pro-IL-1β degradation for IL-1β secretion [33,34,35]. Collectively, our findings suggest that IL-1β dysregulation in autophagy-deficient myeloid cells, particularly macrophages, is a potential mechanism underlying aggravated neutrophilic inflammation in psoriasis.
Autophagy deficiency-mediated aggravation of psoriasis is IL-1 dependentA recent study highlighted the pivotal role of the IL-1β–IL-1R signalling pathway in psoriasis pathogenesis [20]. Thus, to probe the pathogenic role of IL-1β dysregulation in psoriasis, we assessed the effect of IL-1 blockade on psoriatic skin inflammation linked to autophagy deficiency. Mice were systemically administered anti-IL-1R1 antibody, which blocks the interaction of IL-1β with its cognate receptor IL-1R during the development of psoriasis. IL-1 receptor blockade significantly alleviated ear thickness and swelling in both Atg7fl/fl;LysM-Cre and Atg7fl/fl psoriatic mice, regardless of autophagy deficiency (Fig. 3a). Notably, this therapeutic efficacy was most evident in Atg7fl/fl;LysM-Cre mice, which exhibited an abrogation of pathogenic effect linked to autophagy deficiency (Fig. 3a and b). Similarly, epidermis thickness and swelling of dLNs were significantly decreased in both mouse groups, with the most significant effect in the Atg7fl/fl;LysM-Cre mice (Fig. 3c). As a control experiment, treatment with anti-IL-1R1 antibody did not affect ear thickness and swelling in both Atg7fl/fl;LysM-Cre and Atg7fl/fl mice without IMQ treatment (data not shown).
Fig. 3
Effect of IL-1-receptor blockade on psoriatic skin inflammation. Atg7fl/fl or Atg7fl/fl;LysM-Cre mice were injected with anti-IL-1R1 antibody or control IgG and sacrificed at the end of the experiment. The tissues were harvested for each analysis. (a) Ear thickness and swelling were measured every other day (n = 8–10 per group) ***P < 0.001, two-way ANOVA. (b) Cumulative clinical score (erythema, scaling, and thickness) on a scale from 0 to 12. All bars indicate the mean ± SEM. *P < 0.05, Mann–Whitney U-test. (c) Representative ear images (top), H&E-stained histological images (middle), and draining lymph nodes (bottom) from each mouse group. The bar chart shows the epidermal thickness of ear tissue. All dots indicate the number of samples. All bars indicate the mean ± SEM. *P < 0.05 and **P < 0.01, Mann–Whitney U-test. (d) Flow cytometric analysis of the indicated IL-17 A-producing T cells in the draining lymph nodes. All dots indicate the number of samples. All bars indicate the mean ± SEM. *P < 0.05 and **P < 0.01, Mann–Whitney U-test. (e) The levels of IL-1β, IL-12p40, IL-6, and TNF-α were determined using ELISA in harvested ear tissues from each mouse group. All dots indicate the number of samples. All bars indicate the mean ± SEM. *P < 0.05 and **P < 0.01, Mann–Whitney U-test
We investigated the effects of IL-1 blockade on IL-17 A-producing T cells. Following IMQ treatment, both mouse groups showed an increase in IL-17 A+γδ T cell population, which was further increased by autophagy deficiency (Fig. 3d upper). Systemic blockade of IL-1 receptor significantly reduced the IL-17 A+γδ T cell population in terms of percentages and numbers, particularly in autophagy deficiency. Similarly, the same treatment significantly reduced the percentages and numbers of IL-17 A+CD4+ T cells and IL-17 A+CD8+ T cells in dLNs from the Atg7fl/fl;LysM-Cre mice (Fig. 3d middle and bottom). These results suggest a key role of IL-1R signalling in the pathogenesis of psoriasis, linked to autophagy deficiency through its significant effect on IL-17-producing T cells.
We assessed the effect of IL-1 receptor blockade on the production of inflammatory cytokines promoting psoriasis pathogenesis in the ears of IMQ-treated mice. In support of the key role of IL-1β in IMQ-induced psoriasis, blockade of IL-1 receptor significantly diminished the production of IL-12p40 as well as IL-1β in psoriasis-induced skin lesions (Fig. 3e). Although not significant, the production of IL-6 and TNF-α was also apparently reduced by IL-1R blockade in IMQ-treated Atg7fl/fl;LysM-Cre mice (Fig. 3e). These results suggest that IL-1β dysregulation linked to myeloid autophagy dysfunction can induce psoriatic skin inflammation.
Autophagy deficiency-mediated neutrophilic inflammation is IL-1 dependentTo further investigate the therapeutic mechanism of IL-1 receptor blockade, we assessed the mRNA expression of key cytokines and chemokines in the ears of psoriasis-induced mice. We observed a significant upregulation of the mRNA expression of Il1b and Cxcl2 by autophagy deficiency, as expected, which was abrogated by IL-1 receptor blockade (Fig. 4a). In addition, the mRNA expression of Cxcr2 was significantly reduced. In comparison, no significant changes were observed in the mRNA expression of Il12p40, Il6, Tnfa, Il23, and Il1a, following IL-1 receptor blockade.
Fig. 4
Neutrophilic inflammation in autophagy-deficient psoriatic mice is IL-1-dependent. (a) Effect of IL-1R1 signalling blockade on the mRNA expression of genes related to psoriasis pathogenesis. Relative mRNA levels corresponding to the indicated proteins, as determined using qRT-PCR and normalized to β-actin. All dots indicate the number of samples. All bars represent the mean ± SEM. *P < 0.05 and **P < 0.01, as determined using the Mann–Whitney U-test. (b) Representative immunostaining for IL-1β and Ly6G in the ear tissue from each mouse group. The bar chart shows the area of IL-1β positivity colocalized with the neutrophil marker and the number of neutrophils. Scale bars = 50 μm. The number of samples is indicated by dots. All bars indicate the mean ± SEM. *P < 0.05 and **P < 0.01, as determined using the Mann–Whitney U-test
We investigated the recruitment of neutrophils and their expression of IL-1β in the IMQ-induced murine model of psoriasis in the presence or absence of anti-IL-1R1 antibody injection. Colocalization of the augmented IL-1β staining with Ly6G+ neutrophils in autophagy deficiency was significantly reduced by the blockade of IL-1 receptor (Fig. 4b), correlating with the expression of Il1b and Cxcl2. In addition, the anti-IL-1R1 antibody significantly decreased the number of infiltrated Ly6G+ neutrophils in both Atg7fl/fl and Atg7fl/fl;LysM-Cre mice (Fig. 4b), supporting the role of IL-1β–IL-1R signalling in driving neutrophilic inflammation during psoriatic skin inflammation, particularly in the context of autophagy deficiency.
Saccharin suppresses IL-1β expression and alleviates autophagy-deficiency-linked skin inflammationOur previous study revealed that artificial sugars, particularly saccharin, exert an anti-inflammatory effect on macrophage IL-1β production in an autophagy-independent manner [48]. Based on this finding, we investigated whether saccharin ameliorates IMQ-induced psoriatic skin inflammation in Atg7fl/fl;LysM-Cre mice. To this end, we first examined the effects of saccharin on R848-stimulated macrophages in vitro and found that saccharin significantly and dose-dependently reduced the production of IL-1β, in particular, and IL-12p40 by R848-activated macrophages; this effect was comparable between WT and Atg7-deficient macrophages (Fig. 5a). To a lesser extent, the same treatment reduced the production of IL-6 and TNF-α, independent of autophagy deficiency. These results indicated that saccharin exhibits a preferential anti-inflammatory effect on IL-1β and IL-12p40 production, prompting us to examine the therapeutic efficacy of saccharin in psoriasis.
Fig. 5
Effect of saccharin on psoriatic skin inflammation. (a) Effect of saccharin on the production of IL-1β, IL-12p40, IL-6, and TNF-α from R848-stimulated peritoneal macrophages in Atg7fl/fl or Atg7fl/fl;LysM-Cre mice, as determined using ELISA. Data represent at least two independent experiments conducted with duplicates. All bars indicate the mean ± SD. **P < 0.01, Student’s t-test. (b-f) Saccharin or vehicle was applied onto the ear of Atg7fl/fl or Atg7fl/fl;LysM-Cre mice every day prior to IMQ treatment. Mice were sacrificed at the end of the experiment, and tissues were harvested for each analysis. (b) Ear thickness and swelling were measured every other day (n = 8). ***P < 0.001, two-way ANOVA. (c) Cumulative clinical score (erythema, scaling, and thickness) on a scale from 0 to 12. All bars indicate the mean ± SEM. *P < 0.05, Mann–Whitney U-test. (d) Representative ear images (top), H&E-stained histological images (middle), and draining lymph nodes (bottom) from each mouse group. The bar chart shows the epidermal thickness of ear tissue. The number of samples is indicated by dots. All bars indicate the mean ± SEM. *P < 0.05 and **P < 0.01, Mann–Whitney U-test. (e) Flow cytometric analysis of the indicated IL-17 A-producing T cells in the draining lymph nodes. All dots indicate the number of samples. All bars indicate the mean ± SEM. *P < 0.05 and **P < 0.01, Mann–Whitney U-test. (f) The protein levels of IL-1β, IL-12p40, IL-6, and TNF-α in harvested ear tissues from each mouse group, as determined using ELISA. All dots indicate the number of samples. All bars indicate the mean ± SEM. *P < 0.05 and **P < 0.01, Mann–Whitney U-test
To this end, we induced a murine model of psoriasis in Atg7fl/fl;LysM-Cre or Atg7fl/fl mice in the presence or absence of saccharin treatment. Considering the favourable safety profile of saccharin [49] and easy access to psoriatic skin, saccharin was topically applied to the ears of mice 1 h before IMQ treatment every day for a total of 8 days. Saccharin treatment significantly decreased ear thickness and swelling in Atg7fl/fl psoriatic mice and, most notably, in Atg7fl/fl;LysM-Cre psoriatic mice (Fig. 5b). The PASI scores were also significantly decreased by saccharin treatment in both groups of psoriatic mice (Fig. 5c). This therapeutic efficacy of saccharin was further supported by the assessment of epidermis thickness and swelling of dLNs (Fig. 5d). In control experiment, saccharin treatment alone did not affect ear thickness and swelling in both Atg7fl/fl;LysM-Cre and Atg7fl/fl mice without IMQ treatment (data not shown).
Next, we analysed the effect of saccharin on IL-17-producing T cells, a key effector in psoriasis. As seen with IL-1 receptor blockade, saccharin treatment, effectively and significantly reduced the percentages and number of IL-17 A+γδ T cells, in particular, and IL-17 A+CD4+ and IL-17 A+CD8+ cells induced by autophagy deficiency (Fig. 5e). In addition, saccharin treatment significantly decreased the protein levels of IL-1β and IL-12p40 in IMQ-induced Atg7fl/fl;LysM-Cre mice, which was similar to the effect of IL-1 receptor blockade (Fig. 5f). The same treatment significantly suppressed the augmented mRNA expression levels of Il1b and Cxcl2, along with Cxcr2, in IMQ-induced Atg7fl/fl;LysM-Cre mice (Fig. 6a). Consistent with the significant effect of saccharin on Il1b and Cxcl2 expression, the infiltration of CD68+ macrophages and Ly6G+ neutrophils and their production of IL-1β in the psoriatic lesions were markedly reduced by saccharin treatment, particularly in autophagy deficiency (Fig. 6b). Considering that the efficacy of saccharin is comparable to that of IL-1 receptor blockade in reducing the pathological signs of psoriasis, the number of IL-17 A-producing T cells, and inflammatory cytokine levels, we speculate that saccharin is an eligible and alternative candidate for the topical treatment of psoriasis via its anti-inflammatory effect on IL-1β.
Fig. 6
Effect of saccharin on myeloid expression of IL-1β and neutrophilic inflammation. (a) Effect of saccharin on the mRNA expression of genes related to psoriasis pathogenesis. Relative mRNA levels corresponding to the indicated proteins, as determined using qRT-PCR and normalized to β-actin. All dots indicate the number of samples. All bars indicate the mean ± SEM. *P < 0.05 and **P < 0.01, Mann–Whitney U-test. (b) Representative immunostaining for IL-1β and cell markers CD68 or Ly6G in the ear tissue from each mouse group. The bar chart shows the IL-1β-positive area colocalized with each cell marker and the number of cells for each cell type. Scale bars = 50 μm. All dots indicate the number of samples. All bars indicate the mean ± SEM. *P < 0.05, Mann–Whitney U-test
Autophagy-related gene dysregulation and correlation with the Th17 pathway in psoriatic skin lesionsTo extrapolate whether our findings from the murine model of psoriasis have human relevance, we analysed a public microarray dataset (GSE13355) on gene-expression profiles in skin-biopsy specimens from patients with psoriasis and healthy donor subjects [43]. We first assessed the expression profiles of genes related to autophagy and found that psoriatic skin lesions from patients (PP) showed increased expression of ATG7 and BECN1 compared with normal skin biopsies from healthy donors (NN) or normal skin biopsies from patients (NP) (Fig. 7a). In contrast, the expression of other autophagy-related genes, including ATG14, ULK1, ULK2, and SQSTM1 encoding p62, was significantly decreased in psoriatic skin lesions from patients (PP) compared with that in the other specimens (NN and NP). This reciprocal regulation of autophagy-related genes suggests a potential dysregulation of autophagy in human psoriatic lesions, corroborating a recent report of impaired autophagy due to lysosomal dysfunction in psoriasis lesions from patients [37]. Genes related to inflammasomes (NLRP3, CASP1, and PYCARD) and IL-1β–IL-1R1 signalling (IL1B, IL1R1, MyD88, IRAK1, IRAK2, and IRAK4) were significantly overexpressed in psoriatic skin lesions (Fig. 7a). In contrast, no significant changes were observed in the expression levels of IL1A and IRAK3, a down-regulator of IL-1β–IL-1R1 signalling (Fig. 7a). Thereafter, we investigated the correlation of Th17-related genes with these genes. Notably, the up-regulated expression of ATG7 and BECN1 exhibited moderate positive correlation, whereas the down-regulated genes (ATG14, ULK1, ULK2, and SQSTM1) demonstrated a remarkable negative correlation with the Th17 pathway (Fig. 7b). Inflammasome-related genes (NLRP3 and PYCARD) and IL-1β–IL-1R1 signalling-related genes (IL1B, MyD88, IRAK1, IRAK2, and IRAK4) were significantly and positively correlated with genes related to the Th17 pathway (Fig. 7b). To confirm these results, we analysed an independent microarray dataset (GSE30999) on gene expression profiles in non-lesion or psoriasis lesion samples from patients (Supple. Figure 3) [44]. Similarly, we observed a reciprocal dysregulation of autophagy-related genes (ATG7 vs. ATG14 and SQSTM1) and overexpression of genes related to inflammasomes (NLRP3, CASP1, and PYCARD) and IL-1β–IL-1R1 signalling (IL1B, IL1R1, MyD88, IRAK1, IRAK2, and IRAK4), which correlated with the Th17 pathway (Supple. Figure 3). Thus, these findings support a potential link between autophagy dysregulation and inflammasome activation for IL-1β production and the inflammatory Th17 pathway in human psoriatic skin inflammation.
Fig. 7
Gene expression analysis in human psoriatic skin biopsy specimens. (a) Gene expression profiles were examined in normal, non-lesion skin from healthy donors (NN), non-lesion skin from patients with psoriasis (NP), and psoriatic lesions from patients (PP) using the microarray gene expression dataset (GSE13355). (b) Correlation analysis between target genes and TH17 score. The TH17 score was generated by averaging the expression levels of the IL17A, IL17F, IL22, IL23A, IL23R, and IL12B genes. Black, blue, and red dots indicate normal, healthy, non-lesion skin (NN); non-lesion skin from patients with psoriasis (NP); and psoriatic lesions from patients (PP), respectively. P values were calculated using Mann–Whitney U-tests (a) or the Spearman correlation test (b)
留言 (0)