
記住我
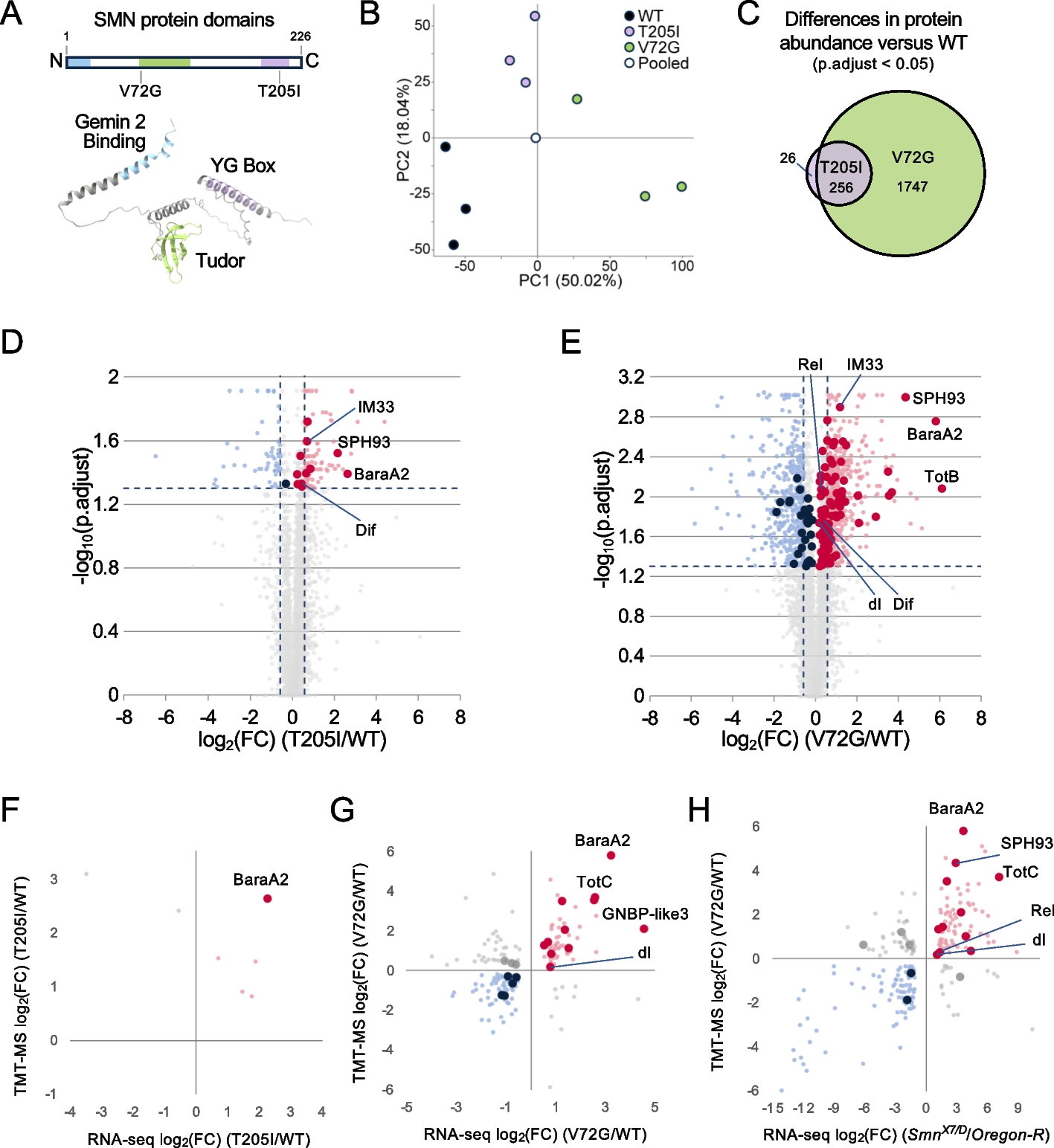






Most insect ORs are expressed in the adult antennae and, usually, only a few ORs are expressed in larvae. Some ORs expressed in both larvae and adults have been shown to exert key functions, for instance in D. melanogaster, Spodoptera littoralis, and H. armigera [16, 26, 27]. We thus searched for such ORs in P. xylostella, as targets for large screening to identify new active volatile compounds. First, the expression profile of 54 PxylORs in the heads (including antennae) of larvae at different instars (first, second, third female, third male, fourth female, and fourth male) and in the antennae of female and male adults were determined (Fig. 1A, Additional File 1: Fig. S1). The P. xylostella ortholog of the universal OR co-receptor (Orco), which is necessary for OR functioning, was found to be expressed in all samples examined. Most pheromone receptors (PRs) were specifically or highly expressed in the male antennae (PxylOR1, PxylOR3, PxylOR4, PxylOR5, PxylOR6, PxylOR7, and PxylOR41) (Additional File 1: Fig. S1).
Fig. 1
PxylOR16 is expressed in all the larval and adult stages of Plutella xylostella and is specifically tuned to heptanal. A Tissue expression patterns of P. xylostella OR genes. The cDNA templates for PCR analyses were from larval heads (first, second, third female, third male, fourth female, and fourth male instar larvae) and adult antennae (male adults: MA, female adults: FA). W water control. Among the 54 P. xylostella ORs, only PxylOR16 and PxylOR27 were detected in all larval stages and in adults. Actin was used for cDNA quality control. B Inward current responses of PxylOR16/PxylOrco Xenopus oocytes to plant volatile compounds (10–4 M). C No ligand was identified for PxylOR27/PxylOrco. D Inward current responses of PxylOR16/PxylOrco Xenopus oocytes stimulated with a range of heptanal concentrations. E Response profile of PxylOR16/PxylOrco Xenopus oocytes to a panel of 71 odorants (n = 6). F Dose–response curve of PxylOR16/PxylOrco Xenopus oocyte responses to heptanal. Heptanal EC50 = 1.757 × 10–5 M. Error bars indicate SEM (n = 6)
Interestingly, only two ORs, PxylOR16 and PxylOR27, were expressed in all examined larval stages and adult antennae of both sexes (Fig. 1A), suggesting they may have important biological functions throughout the insect life cycle. We subsequently expressed these two ORs in Xenopus oocytes to investigate their responsivity to a panel of 71 plant volatile compounds (Additional File 1: Table S1) using two-electrode voltage clamp. The oocytes expressing PxylOR16/Orco were extremely sensitive to heptanal, and they also responded weakly to two other volatiles (octyl acetate and 1-heptanol) (Fig. 1BE). In dose–response studies, we assayed the responses of PxylOR16 to a range of concentrations of heptanal and observed the lowest measurable response at a concentration of 1 × 10−6 M (Fig. 1D). The EC50 value of PxylOR16 for heptanal was 1.757 × 10−5 M (Fig. 1F). These results suggest that PxylOR16 and its ligand (heptanal) are of great significance to P. xylostella. Oocytes expressing PxylOR27 did not respond to any of the 71 examined plant volatile compounds (Fig. 1C).
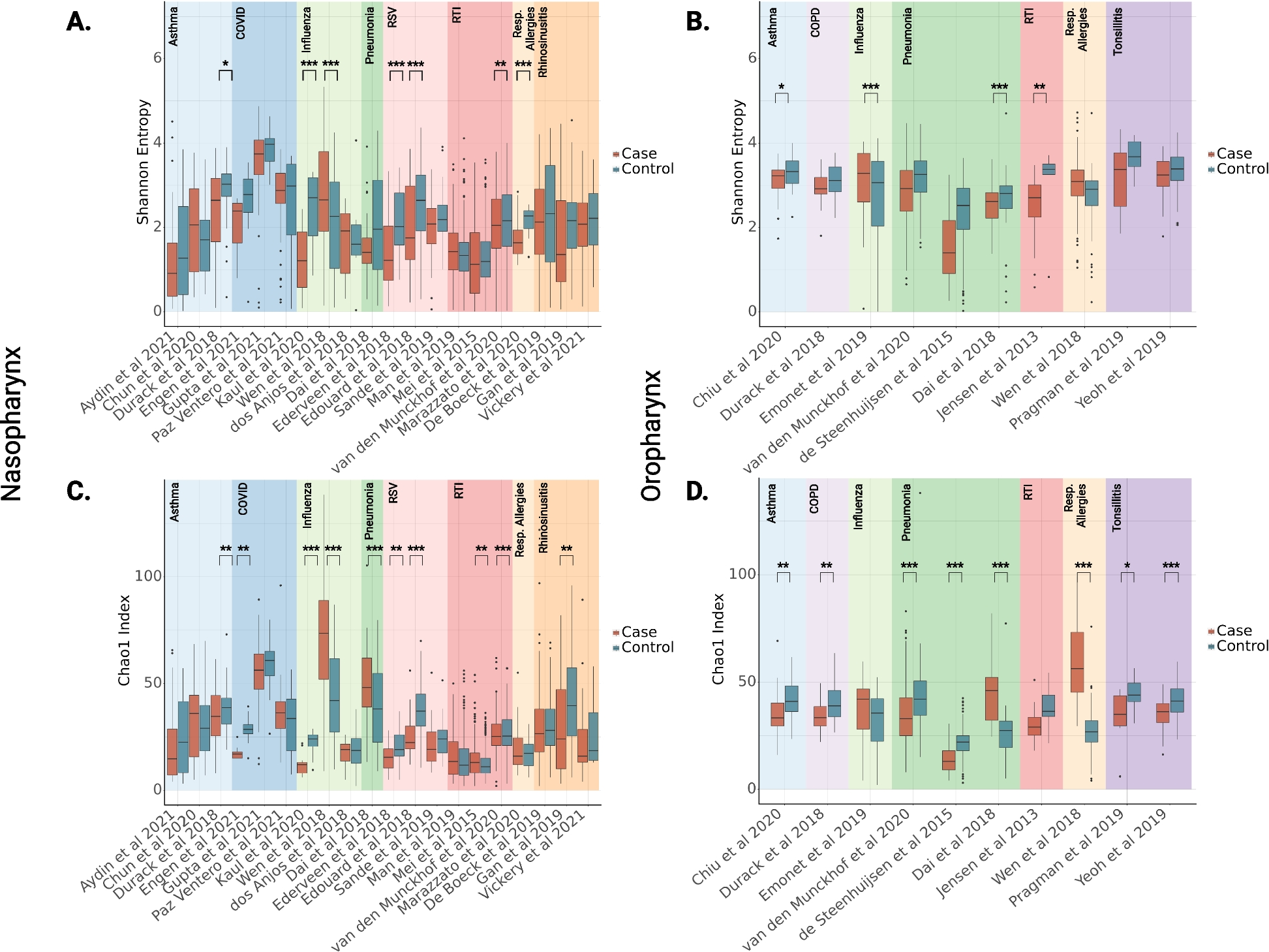
PxylOR16 knockout mutants show impaired electrophysiological responses to heptanalSince heptanal appeared to strongly activate PxylOR16, we wondered whether this compound would also trigger a strong antennal response in vivo, as measured by EAG. Because the antennae of P. xylostella larvae are too small, we were not able to carry out EAG experiments on larvae. We thus recorded the electrophysiological responses of adult female and male antennae to heptanal doses of 10 ng, 100 ng, 1 μg, and 10 μg (Fig. 2B, left). Both female and male antennae displayed a dose-dependent EAG response to heptanal, with responses increasing with increasing heptanal doses. At the highest dose (10 μg), the EAG response values were maximal, with mean response values of 0.77 mV and 0.52 mV for wild-type females and males, respectively. At all doses except the lowest one (10 ng), the male EAG response values were significantly higher than those of females (Fig. 2B, left).We next constructed a PxylOR16-knockout strain using CRISPR/Cas9 genome editing in a traditional and common widely used method in many insects including P. xylostella [28], H. armigera [16], Locusta migratoria [24, 29], and Eupeodes corollae [30]. In all of these works, only one homozygous mutant strain was used. Therefore, knockout mutants (PxylOR16−/−) were obtained with a 5-nt insertion and a 1-nt deletion in the second exon, which introduced a premature stop codon in the coding sequence (Fig. 2A). This PxylOR16-knockout P. xylostella strain (PxylOR16−/−) was using for the further comparing of electrophysiological and behavioral responses with the wild-type (WT) strain. We also used EAG to investigate the electrophysiological response of PxylOR16−/− moths to heptanal. As a control, we tested the response of female and male mutant moths to trans-2-hexen-1-ol, which is not a ligand of PxylOR16, and that is known evoke significant electrophysiological responses in the antennae of both female and male P. xylostella adults. As expected, we observed no difference between the responses of PxylOR16−/− moths and WT moths to this compound (Fig. 2B, right). However, the EAG signals of female and male PxylOR16−/− moths in response to heptanal were significantly reduced compared with those of WT female and male moths, at all examined heptanal doses (Fig. 2B, left). In the abovementioned results, the EAG responses to heptanal and trans-2-hexen-1-ol at different concentrations have been registered on the same individual, whether WT or PxylOR16−/−. These results support the hypothesis that PxylOR16 is involved in heptanal detection.
Fig. 2
PxylOR16 knockout mutants show impaired electrophysiological responses to heptanal. A Schematic diagram of the sgRNA target in Exon II of PxylOR16. The target sequence is shown in blue, the PAM sequence is marked in red, and the non-homologous insertion in the genome in yellow. Deleted bases are represented by dashes. PxylOR16 mutants show impaired electrophysiological responses to heptanal. B Electrophysiological responses measured as electroantennograms of Plutella xylostella antennae to heptanal and trans-2-hexen-1-ol in wild-type (WT) animals and in PxylOR16 knockout mutants generated by CRISPR/Cas9 (PxylOR16−/−). Left, dose-dependent electroantennographic (EAG) responses of female and male moths. Heptanal was used at doses ranging from 10 ng to 10 μg. WT female and male antennae exhibited dose-dependent EAG responses to heptanal, with responses increasing with increasing heptanal doses. EAG responses of female and male PxylOR16−/− moths to heptanal were far lower than those of WT female and male moths at all heptanal doses. Right, as a control, we tested the responses of female and male moths to trans-2-hexen-1-ol, which is not a ligand of PxylOR16. As expected, we observed no differences in the responses to this compound between PxylOR16−/− moths and WT moths. Error bars indicate SEM (n = 10). Different letters indicate significant differences among insects (two-way ANOVA followed by Tukey’s pairwise test; P < 0.05)
Heptanal elicits obvious avoidance behaviors in P. xylostella larvae and adultsAs PxylOR16 was expressed in both larvae and adults, and having confirmed that PxylOR16 responded to heptanal, we next investigated the effect of this ligand on the behavior of P. xylostella adults and larvae. Behavioral experiments were conducted independently on female and male adults. Adult behavioral responses were tested using a Y-tube two-choice bioassay, while larvae responses were tested using a Petri dish assay (static air). As a control, we selected sex pheromone (Z-11-hexadecenal) to verify whether there is a difference in recognition of sex pheromones between the WT and PxylOR16−/−. The results of behavioral experiments showed that both WT and PxylOR16−/− male moths were significantly attracted to Z-11-hexadecenal, and there was no significant difference between the two strains (Additional File 1: Fig. S2).
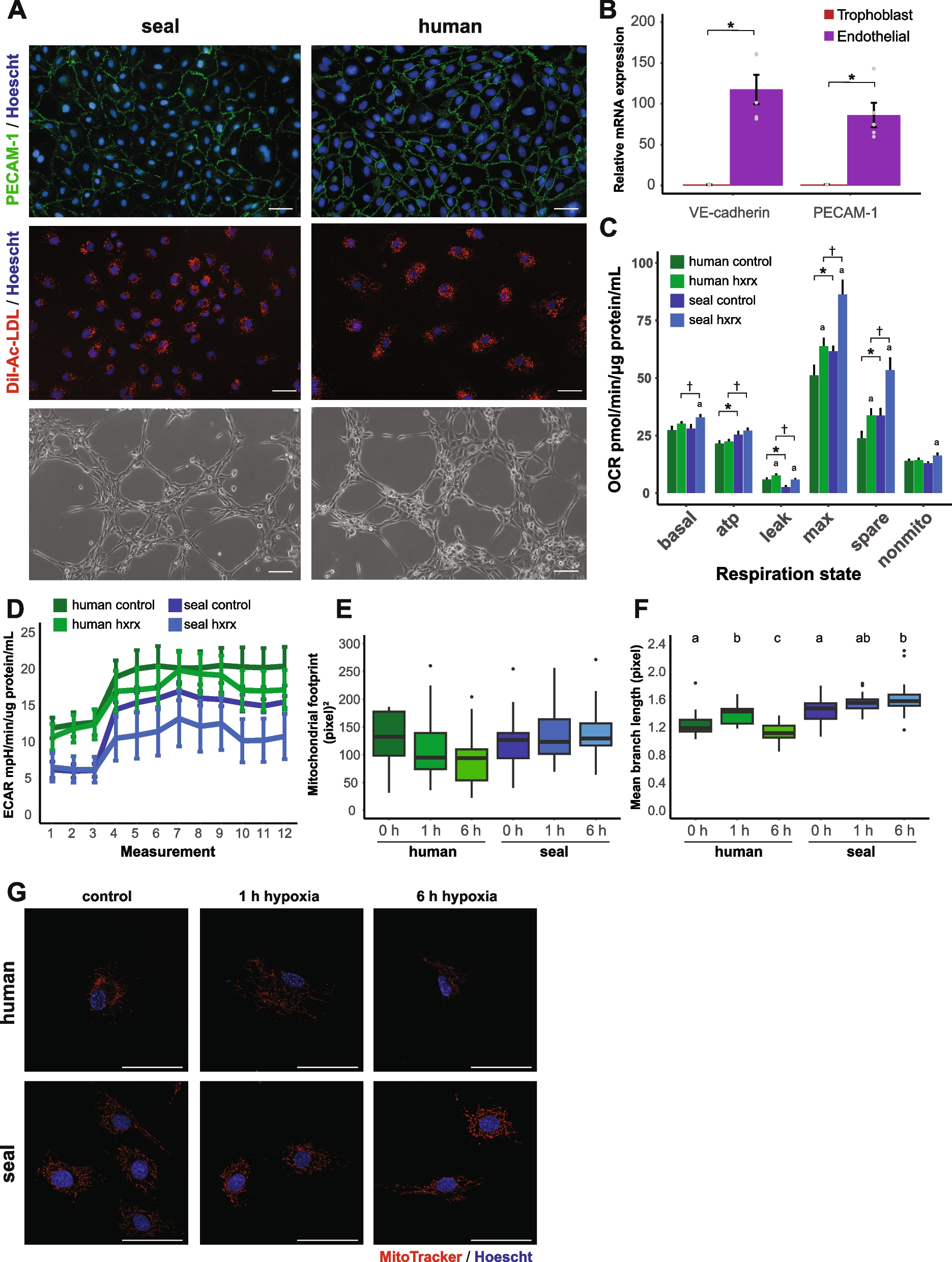
According to the Y-tube two-choice bioassay for adults, we first validated this assay by performing control experiment in which insects were exposed to filter paper without any odorant added on both sides. We observed that neither WT nor PxylOR16−/− adults displayed any selection preference (Fig. 3A). Then, we tested adult behavioral responses to heptanal at different concentrations. The experiment revealed that male moths displayed significant avoidance to heptanal compared with the paraffin oil control at doses of 100 ng, 1 μg, and 10 μg (Fig. 3A, left). The same avoidance behavior was observed for female moths, but only at doses of 1 μg and 10 μg (Fig. 3A, right), suggesting that female and male moths have different sensitivities to heptanal, in accordance with the EAG results.
Fig. 3
Heptanal elicits obvious avoidance behaviors in both Plutella xylostella larvae and adults. Preference index of P. xylostella wild-type (WT) and CRISPR/Cas9 PxylOR16 knockout (PxylOR16−/−) adults and larvae for heptanal. Preference indexes = (number of choices at treatment – number of choices at control) / number of total choices. A Preference index of WT and PxylOR16−/− mutant female and male moths for control (filter paper vs filter paper) and for filter paper + paraffin oil vs filter paper + heptanal in a Y-tube olfactometer (n = 16). WT male moths exhibited significant avoidance to heptanal at doses of 100 ng, 1 μg, and 10 μg (left). Avoidance was also observed for WT female moths, but only at doses of 1 μg and 10 μg (Welch’s t-test; NS, no significant difference, P > 0.05; *** P < 0.001) (right). PxylOR16−/− mutant female and male moths did not show a preference for one side of the device (Welch’s t-test; NS, no significant difference, P > 0.05). B Preference index of WT and PxylOR16−/− mutant female and male moths for blank (one side blank vs a piece of Brassica pekinensis), for control (a piece of B. pekinensis vs a piece of B. pekinensis), and for a piece of B. pekinensis + paraffin oil vs a piece of B. pekinensis + heptanal in a Y-tube olfactometer (n = 16). WT and PxylOR16−/− mutant female and male moths were significantly attracted to B. pekinensis, compared with blank. WT male moths were significantly less attracted to B. pekinensis + heptanal at doses of 100 ng, 1 μg, and 10 μg than to B. pekinensis alone (with paraffin oil) (left). Female moths were also less attracted to B. pekinensis + heptanal than to B. pekinensis alone but only at heptanal doses of 1 μg and 10 μg (Welch’s t-test; NS, no significant difference; P > 0.05; *** P < 0.001) (right). The PxylOR16−/− mutant female and male moths lost their avoidance responses to heptanal significantly, compared with WT adult female and male moths (Welch’s t-test; NS, no significant difference; P > 0.05). C Preference indexes of 10 WT and mutant female and male third instar larvae for blank (one side blank vs a piece of B. pekinensis), for control (a piece of B. pekinensis vs a piece of B. pekinensis), and for a piece of B. pekinensis + paraffin oil vs a piece of B. pekinensis + heptanal in a 10-cm diameter plastic Petri dish (n = 28). Both sexes of WT and mutant larvae were attracted to B. pekinensis. Significantly more female and male third instar larvae were located in the control area (B. pekinensis + paraffin oil) than in the heptanal-supplemented area (B. pekinensis + heptanal) at all heptanal doses (100 ng, 1 μg, and 10 μg on filter paper) except for 10 ng (Welch’s t-test; NS, no significant difference, P > 0.05; *** P < 0.001). As shown for PxylOR16−/− adults, detection of heptanal was abolished in PxylOR16−/− larvae (Welch’s t-test; NS, no significant difference, P > 0.05)
Then, we performed a second experiment on adults using the Y-tube two-choice bioassay to test the response of P. xylostella adults to heptanal in the presence of the host Brassica pekinensis leaf (1 × 1 cm2, the oviposition material for adults used in our laboratory) as the attractant in both tube arms. In the one side blank experiment, we added a piece of B. pekinensis in one arm and kept the other arm blank. Both WT and PxylOR16−/− adults exhibited a preference for B. pekinensis over the blank (Fig. 3B, Blank). This result showed that the B. pekinensis leaf was attractive to adults, whatever the genotype. When adults were challenged with B. pekinensis on both sides, neither WT nor PxylOR16−/− adults displayed any selection preference (Fig. 3B, Control). When we compared the choice between B. pekinensis and B. pekinensis + heptanal, we found that WT male moths were significantly less attracted to B. pekinensis + heptanal than to B. pekinensis alone (with paraffin oil), at heptanal doses of 100 ng, 1 μg, and 10 μg (Fig. 3B, left). Female WT moths were also less attracted to B. pekinensis + heptanal than to B. pekinensis alone, but only at heptanal doses of 1 μg and 10 μg (Fig. 3B, right), indicating that male moths were still more sensitive to heptanal than female moths in the presence of B. pekinensis. Consistent with our finding that PxylOR16 mediates P. xylostella perception of heptanal, PxylOR16−/− adults in the Y-tube two-choice bioassay showed no preference under any examined heptanal concentration (Fig. 3AB).
Larvae behavioral responses were tested using a Petri dish assay (static air). As for the Y-tube assay conducted in adults, we first conducted a control one side blank experiment in which a piece of B. pekinensis (1 × 1 cm2, the food source for larvae used in our laboratory) was added on one side of the dish, while keeping the other side blank. This result showed the food source was also attractive to larvae (Fig. 3C, Blank). When larvae were challenged with B. pekinensis on both sides, neither WT nor PxylOR16−/− larvae displayed any selection preference (Fig. 3C, Control). In view of these results, we used the Petri dish assay to test the response of P. xylostella larvae to heptanal (heptanal alone vs. paraffin oil control) in the absence of food (B. pekinensis) and found that P. xylostella larvae did not make effective choices. We next tested the response of P. xylostella larvae to heptanal in the presence of a piece of B. pekinensis using the Petri dish assay. Significantly more female and male third instar larvae were located in the control area (B. pekinensis + paraffin oil) than in the heptanal-supplemented area (B. pekinensis + heptanal) at all heptanal doses (100 ng, 1 μg, and 10 μg on filter paper) except at 10 ng (Fig. 3C). To verify whether PxylOR16 was responsible for this effect in the larvae, we challenged PxylOR16−/− larvae in the Petri dish assay. As shown for PxylOR16−/− adults, detection of heptanal was abolished in PxylOR16−/− larvae (Fig. 3C). Taken together, these results suggest that PxylOR16 regulates the heptanal avoidance behavior of P. xylostella larvae and adults.
Heptanal does not participate in the direct interaction between P. xylostella and its host plantHeptanal is a common plant volatile, so we firstly speculated that heptanal may mediate interactions between P. xylostella and host plants. We conducted a series of behavioral experiments to test if heptanal participates in the direct interaction between P. xylostella and Brassica vegetables. We chose Brassica plants for experiments because it is the most important genus of cruciferous plants and also the favorite host plant of P. xylostella. In the past 20 years, P. xylostella has become the most destructive insect pest on Brassica vegetables [21]. Therefore, we selected a common Brassica vegetable (B. parachinensis) for the following experiments.
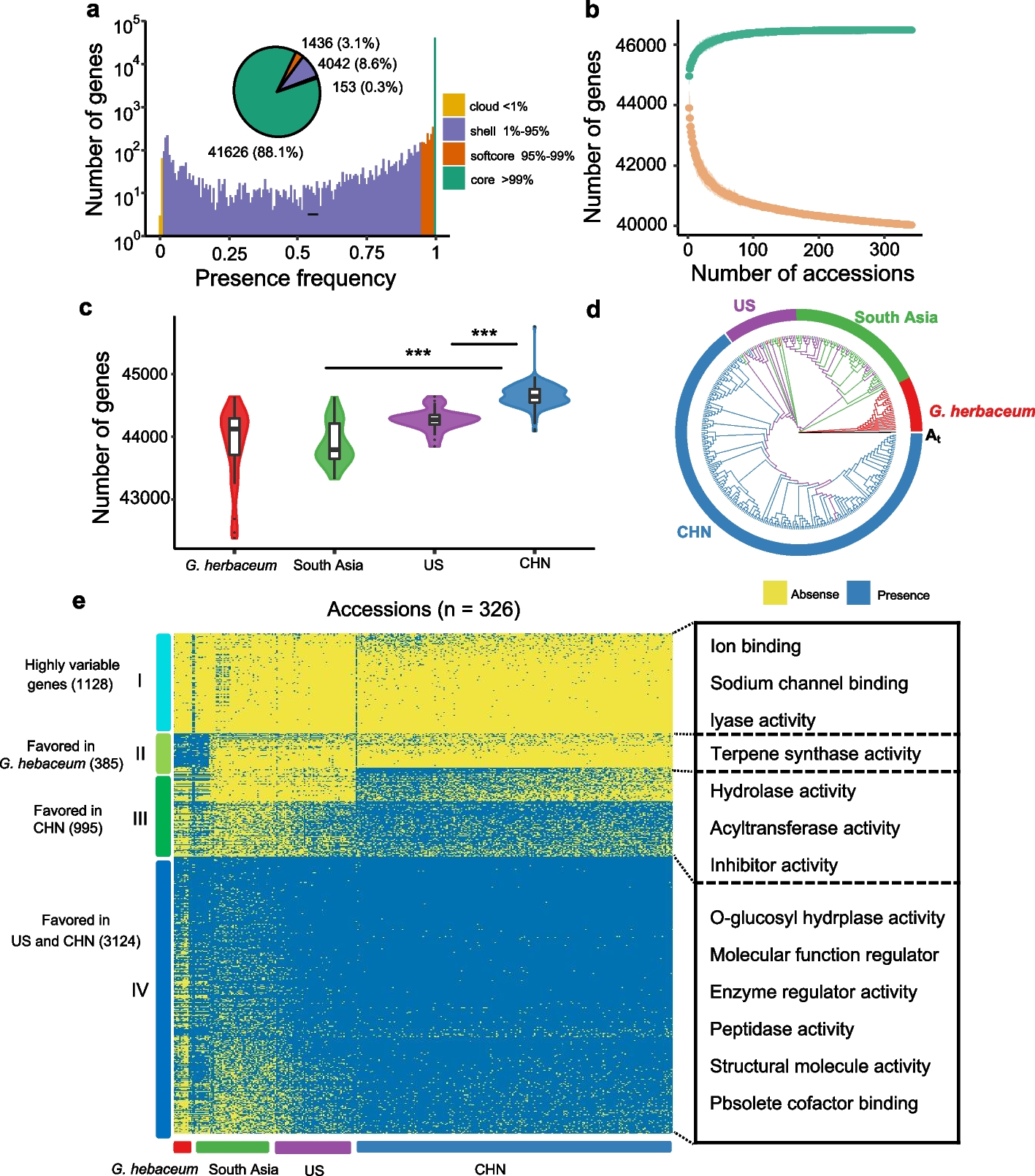
The effect of heptanal in the presence of the host plant B. parachinensis on the adults and larvae were tested by using Y-tube two-choice bioassay and a screening device (with airflow) separately. The responses of P. xylostella adults and larvae to a healthy B. parachinensis plant were tested in one side blank experiments. The healthy B. parachinensis plant did have a significant attracting effect on both WT and PxylOR16−/− adults and larvae, and neither WT nor PxylOR16−/− insects displayed any preference in control experiments when B. parachinensis was present in both sides (Fig. 4AB).
Fig. 4
Heptanal does not participate in the direct interaction between Plutella xylostella and its host plant. Preference index of P. xylostella wild-type (WT) and PxylOR16-knockout (PxylOR16−/−) adults and larvae to heptanal. A Preference index of WT and PxylOR16−/− mutant female and male moths for blank (one side blank vs an intact healthy Brassica parachinensis plant), for control (an intact healthy B. parachinensis plant vs an intact healthy B. parachinensis plant), and for an intact healthy B. parachinensis plant + paraffin oil vs an intact healthy B. parachinensis plant + heptanal in a Y-tube olfactometer (n = 16). WT and PxylOR16−/− mutant female and male moths were significantly attracted to healthy B. parachinensis plant, compared with blank. Three doses (100 ng, 1 μg, and 10 μg) and two doses (1 μg and 10 μg) of heptanal elicited avoidance behavior in WT female and male P. xylostella adults, (Welch’s t-test; NS, no significant difference, P > 0.05; *** P < 0.001). The numbers of PxylOR16−/− adults on each side of the Y-tube olfactometer were not significantly different at all heptanal doses tested, when comparing B. parachinensis + heptanal with B. parachinensis plant + paraffin oil (Welch’s t-test; NS, no significant difference, P > 0.05). B Preference indexes of 10 WT and mutant female and male third instar larvae for blank (one side blank vs an intact healthy B. parachinensis plant), for control (an intact healthy B. parachinensis plant vs an intact healthy B. parachinensis plant), and for an intact healthy B. parachinensis plant + paraffin oil vs an intact healthy B. parachinensis plant + heptanal in a screening device (n = 10). Both sexes of WT and mutant larvae were attracted to the B. parachinensis plant. WT female and male larvae (third instar larvae) preferred the healthy plant with paraffin oil to a healthy plant with heptanal at three doses (100 ng, 1 μg, and 10 μg), unlike PxylOR16−/− larvae (Welch’s t-test; NS, no significant difference, P > 0.05; *** P < 0.001). C GC–MS analysis of the volatiles produced by larval-infested plants and healthy plants; heptanal was not detected. Larval-infested B. parachinensis (top), healthy B. parachinensis (middle), and synthetic heptanal (bottom). D Preference index of WT and PxylOR16−/− mutant adults for control (intact healthy B. parachinensis plant vs intact healthy B. parachinensis plant) and for an intact healthy B. parachinensis plant vs a larval-infected B. parachinensis plant in a Y-tube olfactometer (n = 16). There was no difference in selection between the WT and PxylOR16−/− mutant; both preferred the larval-infected plant (Welch’s t-test; NS, no significant difference, P > 0.05; *** P < 0.001). E Preference index of WT and PxylOR16−/− mutant larvae for control (intact healthy B. parachinensis plant vs intact healthy B. parachinensis plant) and for an intact healthy B. parachinensis plant vs a larval-infected B. parachinensis plant in a screening device (n = 10). Like adults, WT and PxylOR16−/− mutant larvae both preferred the larval-infected plant (Welch’s t-test; NS, no significant difference, P > 0.05; *** P < 0.001)
In subsequent experiments, female and male moths were given a choice between an intact B. parachinensis plant without and with different doses of heptanal in a Y-tube assay. Three doses (100 ng, 1 μg, and 10 μg) and two doses (1 μg and 10 μg) of heptanal elicited avoidance behaviors in female and male P. xylostella adults, respectively (Fig. 4A). In larvae, we observed a preference of female and male larvae (third instar larvae) for a healthy B. parachinensis plant with paraffin oil over a healthy plant with heptanal at three doses (100 ng, 1 μg, and 10 μg) (Fig. 4B). The numbers of PxylOR16−/− adults and larvae on each side of the device were not significantly different at all heptanal doses tested (Fig. 4AB). To test whether heptanal is present in the host plant B. parachinensis, we examined the volatiles emitted by the healthy B. parachinensis. Briefly, we used solid-phase microextraction to collect volatiles emitted by healthy plants. Gas chromatography coupled to mass spectrometry (GC/MS) analysis revealed that no heptanal was detectable in healthy B. parachinensis volatiles (Fig. 4C).
We found that an intact B. parachinensis plant had a significant attracting effect on P. xylostella and that its presence did not affect the heptanal avoidance behavior, so we further tested if the B. parachinensis plants damaged by P. xylostella feeding will release heptanal and evoke avoidance behavior. Female and male moths and third instar larvae were given a choice between an intact B. parachinensis plant and larval-infected B. parachinensis plant (larvae and feces were removed, same treatment in following experiments) in a Y-tube and in a screening device: the adults and larvae had a preference for larval-infected plants compared with heathy plants (Fig. 4DE). These results were not unexpected, since P. xylostella was previously reported to prefer P. xylostella larval-infected plants [31, 32]. Moreover, like the WT, the PxylOR16−/− adults and larvae also showed a preference for larval-infected plants (Fig. 4DE). Next, we also examined the volatiles emitted by the P. xylostella larval-infested B. parachinensis. GC/MS analysis revealed that no heptanal was detectable in larval-infested B. parachinensis volatiles (Fig. 4C).
These results suggest that heptanal cannot explain the direct interaction between P. xylostella and B. parachinensis (intact or pest infested). Similarly, many other Brassica plants do not release heptanal [33], so we speculate that heptanal is not involved in the interactions between P. xylostella and host plants as a plant volatile.
P. xylostella larvae and adults detect and avoid Cotesia vestalis odorantsHeptanal was previously reported to be a cuticular volatile emitted and sensed by some Cotesia parasitic wasp species, including C. glomerata and C. marginiventris, and this compound is a required component of their sex communication system [34, 35]. Both females and males of C. glomerata, a parasitic wasp of Pieris brassicae, release heptanal as a repellent pheromone. After mating, females release heptanal to repel males to avoid mating again, while the males release heptanal to repel conspecific males, thus reducing male–male competition on the natal patch and ensuring maximal mating success. The amount of heptanal released by C. glomerata was analyzed. It was previously found that about 3 ng can be obtained from 6 individuals by solvent extraction, and that about 50 ng can be obtained from 200 individuals by headspace extraction [34]. In a phylogeny of 25 species of Cotesia commonly used in laboratory and field research, C. vestalis is the closest to C. glomerata [36]. We therefore turned our attention to C. vestalis, which is an endoparasitoid wasp that is known to attack P. xylostella larvae; indeed, it is used as an agricultural control measure against P. xylostella [37].
We hypothesized that, like C. glomerata, C. vestalis releases heptanal. To test this hypothesis, we collected cuticular compounds of this wasp using dichloromethane for GC/MS analysis. Consistent with the hypothesis that C. vestalis releases heptanal, we could detect heptanal in the extracts of a mixture of 10 female and 10 male wasps, and the amount collected from these twenty wasps was 2.47 ± 0.55 ng (Fig. 5AB).
Fig. 5
Plutella xylostella larvae and adult detect and avoid Cotesia vestalis odorants. A GC/MS analysis of the volatiles produced by C. vestalis. Heptanal could be detected in extracts of C. vestalis. IS internal standard. B Synthetic heptanal. C Preference index of WT and PxylOR16−/− P. xylostella mutant adults (left) and larvae (right) for filter paper + dichloromethane vs filter paper + C. vestalis body wash in a Y-tube olfactometer and a 10-cm diameter plastic Petri dish, respectively (n = 16, adults; n = 28, larvae). Both sexes of WT larvae and adults of P. xylostella had avoidance responses to a C. vestalis body wash, but the PxylOR16−/− mutant did not (Welch’s t-test; NS, no significant difference; P > 0.05; * P < 0.05; *** P < 0.001). D Preference index of WT and PxylOR16−/− mutant adults (left) and larvae (right) for air + 20 live C. vestalis vs air in a Y-tube olfactometer and a screening device, respectively (n = 16, adults; n = 10, larvae). Both sexes of WT larvae and adults of P. xylostella showed avoidance of C. vestalis, but the PxylOR16−/− mutants did not (Welch’s t-test; NS, no significant difference; P > 0.05; * P < 0.05; ** P < 0.01)
Both D. melanogaster adults and larvae have an avoidance response to Leptopilina boulardi, which is a parasitic wasp of D. melanogaster larvae [26]. To test whether larvae and adults of P. xylostella have a similar avoidance response to C. vestalis odorants, we next conducted a behavioral experiment in two different experimental treatments: (1) a C. vestalis body wash obtained by extracting 20 wasps (females and males) with dichloromethane for 30 min and (2) 20 live C. vestalis wasps (females and males) placed in glass jars connected by a Y-tube or a screening device. For the first experimental treatment, a Y-tube two-choice bioassay and a Petri dish assay revealed that a C. vestalis body wash elicited avoidance responses in adults (Fig. 5C, left) and in larvae (Fig. 5C, right). Both adults (Fig. 5D, left) and larvae (Fig. 5D, right) of P. xylostella also showed avoidance behavior in response to live C. vestalis in a Y-tube two-choice bioassay and using a screening device.
In behavioral experiments using PxylOR16−/−, we found that both PxylOR16−/− larvae and adults no longer had an avoidance response to C. vestalis body wash or live C. vestalis (Fig. 5CD). These results show that P. xylostella larvae and adults use PxylOR16 to detect heptanal released by C. vestalis and thereby to regulate their avoidance to C. vestalis.
留言 (0)