
記住我






To test how severely mitochondrial perturbation affects the host, we individually silenced genes from OXPHOS complexes I-V (cI-cV) in the immune tissues (fat body and hemocytes) as well as ubiquitously (whole animal). From each of the five OXPHOS complexes, one to two genes were targeted in the knockdown experiments (Fig. 1A, Additional file 1: Table S1 & S2). Ubiquitous knockdown (da-GAL4) of individual OXPHOS genes resulted in a severe larval developmental delay and was eventually lethal (Additional file 1: Table S3). In the case of the fat body-specific gene silencing (Fb-GAL4) the effects varied from developmental delay to lethality, depending on the OXPHOS complex. Fat body-specific knockdown of ND-75 (cI) and SdhD (cII) resulted in a development delay but viable adults, whereas knockdown of ox (cIII), UQCR-C1 (cIII), COX5B (cIV) and ATPsynCF6 (cV) were partially or fully lethal at the pupal stage (Additional file 1: Table S3). In contrast, knocking down the OXPHOS genes in hemocytes (HmlΔ-GAL4; He-GAL4) did not affect the development time or eclosion of flies.
Fig. 1
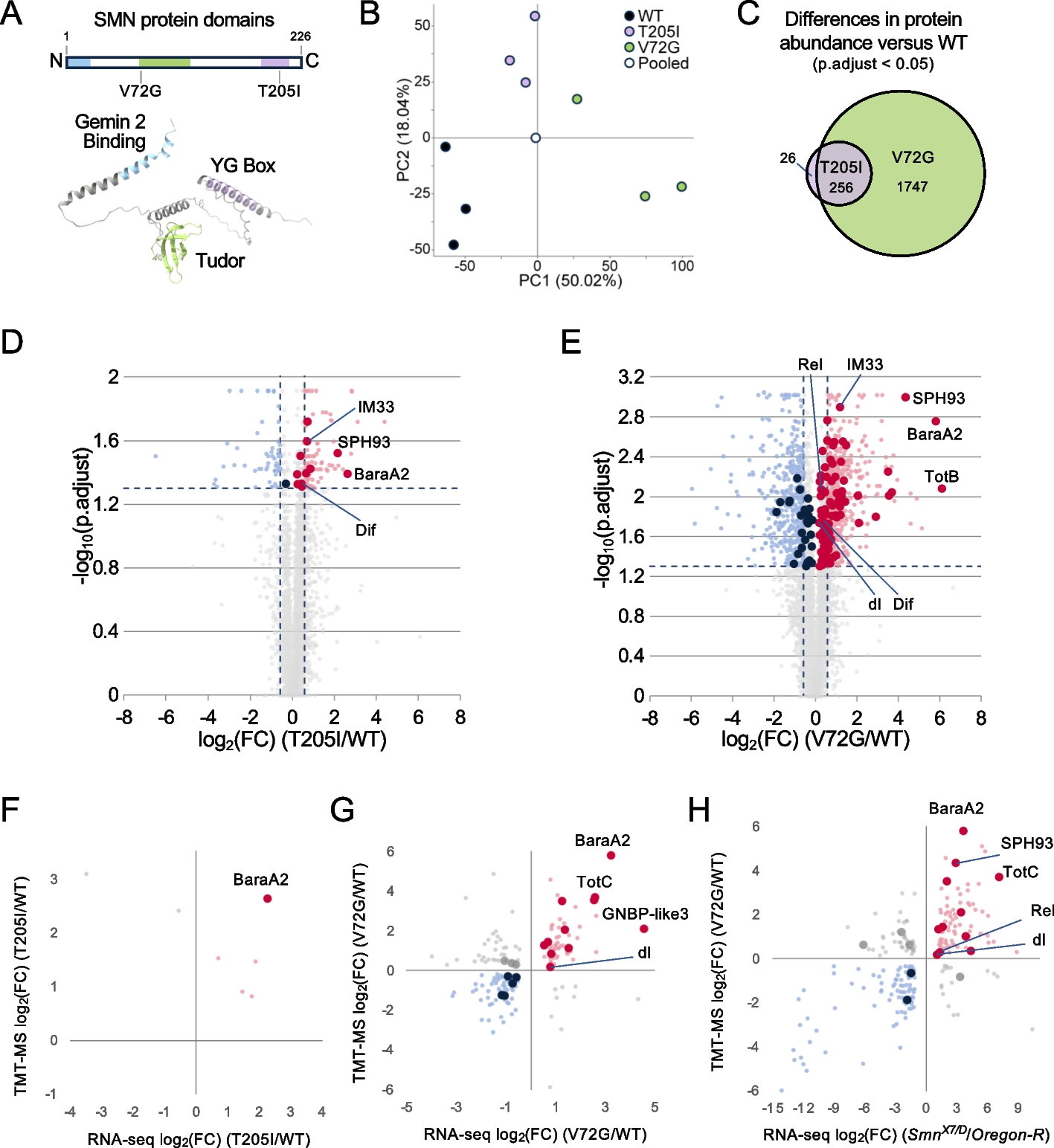
Knockdown of the OXPHOS complex genes in hemocytes affects the mitochondrial membrane potential, causes melanotic nodules and enhances the immune competence. A A schematic representation of oxidative phosphorylation (OXPHOS) complexes and the complex-specific knockdown target genes: ND-75 (NADH dehydrogenase (ubiquinone) 75 kDa subunit); SdhD (Succinate dehydrogenase, subunit D); ox (oxen); UQCR-C1 (Ubiquinol-cytochrome c reductase core protein 1); COX5B (Cytochrome c oxidase subunit 5B); ATPsynCF6 (ATP synthase, coupling factor 6). B Mitochondrial membrane potential was measured as a ratio of the MitoProbe™ TMRM signal intensity detected in the OXPHOS knockdown plasmatocytes to the signal detected in control plasmatocytes (n = 3, 2000–3500 eater-GFP-positive plasmatocytes per replicate). The data were analyzed using one sample t-test. C Examples of melanotic nodules found from hemocyte-targeted cV knockdown animals. Nodules are marked with arrowheads. Scale bars 500 µm. D Quantification of melanotic nodules detected in the hemocyte-targeted OXPHOS gene knockdown larvae (n = 100). * = 1% of the driverless background control larvae had melanotic nodules. E Mean percentage of melanized wasp eggs/larvae found in the controls and in larvae with OXPHOS knockdown in hemocytes (n = 150). E’ Melanization response to wasp infection in the animals with ND-75 and ATPsynCF6 knockdown in the fat body (n = 150). The data were analyzed using logistic regression with a binomial distribution with replication as a random factor. ns = not significant, * p < 0.05, ** p < 0.01, *** p < 0.001. (Schematic Fig. 1A modified from [10])
Taken together, the effect of OXPHOS perturbation varied according to the affected OXPHOS complex and the target tissue. Fat body-specific knockdown of cI-cII genes resulted in a milder effect on the viability than cIII-cV gene knockdown, while hemocyte-specific knockdown did not affect the development time or eclosion of the flies. Given the central role of hemocytes in cell-mediated innate immunity, we focused on the effects of OXPHOS perturbation in larval hemocytes on cellular innate immune response.
Knockdown of OXPHOS genes in hemocytes decreases the mitochondrial membrane potential and induces the formation of melanotic nodulesFirst, we verified that the RNAi constructs efficiently silenced the expression of their target genes in hemocytes. ND-75, SdhD, UQCR-C1 and ATPsynCF6 expression was significantly reduced, ranging from 57 to 91% of the control levels in females (Additional File 2: Fig. S1A) and from 69 to 94% in males (Additional File 2: Fig. S1A’). We were not able to measure the RNAi efficiencies of ox and COX5B, as the RNAi constructs’ hairpin sequence covers most of the gene length, leaving no space to design appropriate reverse transcription quantitative real-time PCR (RT-qPCR) primers for the remaining gene sequence.
To verify that the silencing of the OXPHOS genes resulted in mitochondrial perturbation, we measured the mitochondrial membrane potential (ΔΨm) as a readout of mitochondrial activity. ΔΨm was measured from plasmatocytes using a membrane potential sensitive MitoProbe dye. As a control, hemocytes were treated with carbonyl cyanide 3-chlorophenylhydrazone (CCCP), an uncoupler of oxidative phosphorylation in mitochondria causing a drastic drop in the ΔΨm (Additional File 2: Fig. S1B). We found that cIII-cV knockdown hemocytes had significantly lower MitoProbe signal intensity than the untreated control hemocytes, corresponding to a 40–80% decrease in ΔΨm (Fig. 1B, Additional File 2: Fig. S1B). In contrast, knockdown of cI-cII did not have a significant effect on the MitoProbe signal (Fig. 1B).
Next, we investigated if silencing of the OXPHOS genes alters the cell-mediated innate immune response of the host. Salminen et al. [31] showed that a mtDNA-encoded OXPHOS cIII gene cytochrome b (mt:cyt-b) variant was associated with the formation of melanotic nodules in Drosophila. Melanotic nodules are melanized hemocyte aggregates and are indicative of a pre-activated cell-mediated immune system [32]. We found that silencing any of the OXPHOS genes in hemocytes led to the formation of melanotic nodules to varying degrees, examples of which are shown in Fig. 1C. The prevalence of the nodules ranged from 2 to 14% in the GD library RNAi lines (Fig. 1D), and the finding was further confirmed, although with a milder phenotype, with an additional set of RNAi lines from the KK library with a different genetic background (Additional File 3: Fig. S2A). Again, the strongest phenotypes were observed with cIII-cV gene knockdowns (Fig. 1D). In the control animals, 0–1% of the larvae exhibited melanotic nodules (Fig. 1D).
Hemocyte-targeted OXPHOS gene knockdowns did not affect the development time or eclosion. Nevertheless, perturbation of OXPHOS decreased the mitochondrial membrane potential in hemocytes and activated cell-mediated innate immunity, which could potentially affect other life-history traits of the flies. Therefore, we tested whether cIII and cV gene knockdowns in hemocytes had an effect on the lifespan of the flies. While the male longevity was not affected by the cIII or cV knockdown, females with cIII knockdown exhibited significantly decreased lifespan (Additional File 3: Fig. S2B). The reduced lifespan in females upon cIII knockdown in hemocytes might suggest that there are differences in the demands for hemocyte metabolism in adult flies between the sexes.
OXPHOS perturbation in hemocytes enhances the cell-mediated encapsulation response against parasitoid waspsBecause melanotic nodules indicate activation of the cellular innate immune response [32], we examined if OXPHOS perturbation in hemocytes affected the cell-mediated response against parasitoid wasp infection. We infected the knockdown and control larvae with Leptopilina boulardi parasitoid wasps and scored the percentage of larvae with a successful immune response (fully melanized wasp larvae). Knockdown of the OXPHOS genes resulted in an enhanced encapsulation response when compared to the controls (Fig. 1E). A similar trend was observed when knocking down the OXPHOS genes using the KK RNAi lines (Additional File 3: Fig. S2C), although the effect was again less pronounced than in the GD RNAi lines.
Next, we tested if OXPHOS perturbation in the fat body also resulted in an altered response to wasp infection. Fat body specific (Fb-GAL4) knockdown of ND-75 (cI) had the mildest larval development delay when compared to other OXPHOS complex knockdowns, and ATPsynCF6 (cV) knockdown had one of the most severe effects on viability (Additional file 1: Table S3). In both knockdown lines, the encapsulation response was drastically reduced when compared to the control (Fig. 1E’). Taken together, OXPHOS perturbation in the fat body had a negative impact on viability and led to an increased susceptibility to immune challenge, indicating the importance of tightly regulated mitochondrial function in this tissue. In contrast, hemocyte-targeted OXPHOS perturbation did not affect development and was beneficial for the host upon an immune challenge. This suggests that OXPHOS acts as a moderator of hemocyte function under normal conditions.
OXPHOS perturbation in hemocytes results in immune cell activationHaving established a link between mitochondrial perturbation in hemocytes, formation of melanotic nodules and an enhanced immune response against parasitoid infection, we tested if silencing the OXPHOS genes in hemocytes affects hemocyte numbers or differentiation. We utilized an in vivo hemocyte reporter system (msn-mCherry and eater-GFP; “Me” for short) to distinguish hemocyte subpopulations based on the levels of the mCherry and GFP fluorescence as described in Anderl et al. [28]. As expected, in the control larvae mainly plasmatocytes were present, as shown by high levels of eater-GFP expression (Fig. 2A). In contrast, knocking down OXPHOS cIII, cIV and cV-related genes resulted in an immune activation of the hemocytes, with uninfected larvae producing infection-specific hemocytes, including lamellocytes, identified by high msn-mCherry expression and a lack of the eater-GFP expression (Fig. 2A-B). In addition to hemocyte activation, total hemocyte numbers were significantly increased upon cIV and cV knockdowns, without a change in basal state plasmatocyte numbers (Fig. 2C-C’), but with an increase in activated plasmatocytes (Fig. 2C’’), which are characterized by high GFP expression together with mCherry fluorescence, which is often localized in cytoplasmic foci [28]. These hemocytes are not present in high numbers in healthy larvae, but appear after wasp infestation or genetic activation of the hemocytes [28]. Numbers of lamelloblasts, the putative lamellocyte precursor cell type [28], were increased by cIII and cV knockdowns (Fig. 2C’’’), whereas prelamellocytes (Fig. 2C’’’’), and to an even greater extent lamellocytes (Fig. 2C’’’’’), were increased upon cIII, cIV and cV gene knockdowns (Fig. 2C’’’’). cI and cII knockdown had mild effects on hemocyte populations (Fig. 2A, C–C’’’’’). Knocking down the OXPHOS genes using the KK library RNAi constructs showed similar results concerning hemocyte activation (Additional File 4: Fig. S3A-A’’’’’), except for knockdown of ND-75, which resulted in the formation of prelamellocytes and lamellocytes (Additional File 4: Fig. S3A’’’’-A’’’’’), indicating that cI silencing may also induce lamellocyte formation in specific genetic backgrounds.
Fig. 2
Knockdown of the OXPHOS genes in hemocytes induces immune cell activation. A Flow cytometry analysis of larval hemocytes using the plasmatocyte (eater-GFP, y-axis) and lamellocyte (msn-mCherry, x-axis) in vivo hemocyte reporters. While the control animals mainly had plasmatocytes (high GFP), the knockdown of various OXPHOS genes induced immune-activated hemocyte types, including activated plasmatocytes (high GFP, low mCherry) and lamellocytes (high mCherry intensity, marked in red), and occasionally also lamelloblasts (low GFP) and prelamellocytes (low GFP, low mCherry). B Heatmap (n = 30) showing the average total hemocyte and lamellocyte (lc) count in wGD control and hemocyte-targeted OXPHOS knockdown samples. C–C’’’’’ Quantification of total hemocytes and hemocyte types classified based on eater-GFP and msn-mCherry expression when knocking down the OXPHOS genes (n = 30). Total = total circulating hemocyte count, pc = plasmatocytes, act pc = activated plasmatocytes, lb = lamelloblasts, pre lc = pre-lamellocytes, lc = lamellocytes. cIII gene = UQCR-C1. The data on hemocyte counts were analyzed using a generalized linear model with a negative binomial distribution. Error bars indicate standard error of the mean. Asterisks indicate the statistical difference between the OXPHOS knockdowns and the control. D Mitochondrial membrane potential in plasmatocytes of wasp-infected larvae and of larvae with hepCA or Toll10b overexpression in hemocytes, as a ratio to the MitoProbe™ TMRM signal intensity in the control. ns = not significant, * p < 0.05, ** p < 0.01, *** p < 0.001
Wasp infection is known to cause both elevation in hemocyte numbers and hemocyte activation. We tested if hemocyte-specific cI, cIII or cV knockdown has additional effects on the hemocyte populations 48 h after wasp infection compared to wasp-infected controls. In the wasp-infected larvae, OXPHOS knockdown did not alter hemocyte profiles and produced similar numbers of lamellocytes as the wasp-infected control larvae (Additional File 4: Fig. S3B-B’’’’’). The exceptions were a small decrease in the numbers of activated plasmatocytes in the cIII knockdown larvae, and an increase in lamelloblast count in the cV knockdown larvae (Additional File 4: Fig. S3B’’).
Overall, hemocyte-specific OXPHOS perturbation leads to increased hemocyte counts and immune activation prior to infection. The hemocyte profile induced by wasp infection was mostly unaltered by the OXPHOS knockdown.
Decrease in the mitochondrial membrane potential is not a universal indicator of immune cell activationcIII-cV gene knockdowns caused a decrease in ΔΨm in plasmatocytes, while cI and cII gene knockdowns did not. In parallel, cIII-cV knockdowns led to the formation of melanotic nodules, elevated immune cell count and immune cell activation, which was not the case with the cI and cII gene knockdowns. This prompted us to test whether a decrease in the circulating plasmatocyte ΔΨm occurs when they begin to transdifferentiate into lamellocytes. We selected three model systems that are known to induce the formation of lamellocytes: parasitoid wasp infection, and the expression of constitutively active forms of hemipterous (hepCA) and Toll (Toll10b, Tl10b) [33]. There were no significant differences in ΔΨm compared to the control, although there was a trend for lower ΔΨm in hepCA and Tl10b plasmatocytes (Fig. 2D). Based on this result, we cannot conclude with certainty that a drop in ΔΨm is a common feature of hemocyte activation. However, we do not exclude the possibility that a drop in ΔΨm might be required in hemocytes attached to tissues or in those forming the melanized capsules.
UQCR-C1 (cIII) knockdown hemocyte transcriptomes cluster according to time point and infection statusWe harnessed RNA sequencing to study the mechanism behind OXPHOS perturbation that leads to immune cell activation. For this, we chose cIII gene UQCR-C1 due to its strong effects on the immune phenotypes studied here. We sampled UQCR-C1 knockdown hemocytes from uninfected and wasp-infected male larvae and their controls. Because UQCR-C1 knockdown in hemocytes induces lamellocyte differentiation (Fig. 2), we first identified a timepoint at which lamellocytes are not yet present in the hemolymph, in order to capture transcriptional changes prior to and after the appearance of mature lamellocytes. Based on previous information on the lamellocyte differentiation after wasp infection [28], we profiled the hemocytes of UQCR-C1 knockdown larvae at 90, 92 and 96 h after egg lay (AEL). While lamellocytes were detected in the circulation both 92 h and 96 h AEL, at 90 h AEL lamellocyte numbers did not differ from the age-matched control larvae (Additional File 5: Fig. S4A’’’’’ for lamellocyte counts, Additional File 5: Fig. S4A-A’’’’ for other hemocyte types). In addition, UQCR-C1 knockdown plasmatocytes exhibited reduced ΔΨm at 90 h AEL (Additional File 5: Fig. S4B-B’), showing that the mitochondrial perturbation is present already at that timepoint. The 90 h AEL timepoint was therefore selected as the sample collection timepoint, and is hereafter referred to as the “early” timepoint. In addition, we collected UQCR-C1 knockdown and control hemocytes from late 3rd instar larvae (approximately 120 h AEL) without and with parasitoid wasp infection (48 h post infection), referred to as “late” and “late infected”, respectively (Fig. 3A). Of note, “late infected” refers to a comparison between UQCR-C1 knockdown hemocytes collected from wasp-infected larvae and control hemocytes also from wasp-infected larvae.
Fig. 3
Transcriptomic analysis of UQCR-C1 knockdown hemocytes. A Samples used for RNA sequencing. Hemocytes were collected from male larvae with a hemocyte-specific OXPHOS complex III knockdown (HmlΔ-GAL4; He-GAL4/UAS- UQCR-C1 RNAi) and the controls (HmlΔ-GAL4; He-GAL4/wGD) at 90 h after egg lay (“early” timepoint), 120 h after egg lay (“late” timepoint) as well as from wasp-infected larvae at 120 h after egg lay (“late infected” timepoint). B PCA plot of the RNA sequencing data (n = 3). C UQCR-C1 expression levels in the RNA sequencing samples expressed as counts per million (CPM) showing the efficiency of the knockdown. Figure 3A created with BioRender.com
Principal component analysis (PCA, Fig. 3B) showed a clear clustering of the transcriptome samples along the PC2 according to timepoint and (to a lesser degree) the wasp-infection status. In addition, wasp-infected samples clustered based on their genotype (UQCR-C1 knockdown or control) along PC1. The knockdown of UQCR-C1 was verified based on the counts per million (CPM) values of the RNA sequencing data for all three treatment groups, with a drop in UQCR-C1 expression to 20–45% of the control value (Fig. 3C).
UQCR-C1 knockdown causes upregulation of lamellocyte-enriched marker genes at both timepointsUQCR-C1 knockdown in hemocytes led to distinct changes in the transcriptome. A subset of genes was differentially expressed in all treatments/timepoints when UQCR-C1 was silenced, and multiple genes were differentially regulated only at specific timepoints or upon infection (all significantly differentially expressed genes are listed in Additional File 6). Because UQCR-C1 silencing led to lamellocyte differentiation, many differentially expressed genes were likely to be linked to lamellocyte function, especially at the late timepoint. We utilized a comprehensive list of 304 lamellocyte-enriched marker genes based on the hemocyte single-cell RNA sequencing studies compiled by Hultmark and Ando [34] and cross-referenced these genes with our early and late timepoint UQCR-C1 samples. Indeed, many lamellocyte-enriched genes were upregulated in uninfected UQCR-C1 knockdown samples at the late, and, interestingly, also at the early timepoint (114 and 113 genes, respectively; Additional File 7). There was a large degree of overlap in lamellocyte-enriched genes upregulated at late and early timepoint (94 genes). When comparing infected UQCR-C1 knockdown samples to infected control samples, only 14 lamellocyte genes were upregulated. For comparison, in the wasp-infected control hemocytes, 223 lamellocyte genes were upregulated compared to the uninfected controls (Additional Files 6 and 7). Even though both early and late timepoint UQCR-C1 knockdown hemocytes showed similar upregulation of the lamellocyte marker genes, they clustered separately based on the PCA (Fig. 3B). These data indicate that even though at the early timepoint mature lamellocytes are not yet detected in the larvae, the activation program leading to them is already evident. Next, we studied further the transcriptome profiles between UQCR-C1 knockdown and control samples.
UQCR-C1 knockdown leads to gene expression changes characteristic of the mitochondrial unfolded protein responseWe utilized the Flymine [35] tool to analyze the differentially expressed genes and to construct curated lists of genes of interest based on Gene Ontology (GO) term enrichment. Significant GO terms and genes included in them are listed in Additional File 8. Genes that were consistently differentially expressed when UQCR-C1 was knocked down were of a particular interest since those represent a core set of genes responding to UQCR-C1 silencing, regardless of the infection status or timepoint. Across all of the timepoints and treatments, there was an upregulation of glycolytic enzymes including lactate dehydrogenase, indicating more active use of glycolysis by the cells starting already at an early timepoint prior to lamellocyte formation (Fig. 4A; Additional File 6). Many translation-related genes were also upregulated, including multiple aminoacyl-tRNA synthetases and several genes functioning in mitochondrial translation (Fig. 4A; Additional File 8). This, along with the upregulation of multiple mitochondrial genes involved in heme synthesis, OXPHOS assembly and mitochondrial fusion, might be an effort to compensate for the mitochondrial defect caused by the cIII disruption. Several heat shock response genes (Fig. 4A) and glutathione-S-transferases (Fig. 4A and Additional File 9: Fig. S5A) were upregulated, suggesting a possible stress response in the hemocytes upon OXPHOS knockdown. Of note, genes involved in the same processes (aerobic glycolysis, translation of mitochondrial proteins, heat shock response) are upregulated by the mitochondrial unfolded protein response (UPRmt) in the nematode Caenorhabditis elegans and the yeast Saccharomyces cerevisiae [36, 37], indicating that this phenomenon could also occur in our model. The UPRmt is a cellular stress response involving a mitochondria-to-nucleus signaling pathway that induces a specific transcriptional program. The UPRmt protects cells from stress that originates from mitochondrial perturbation, such as OXPHOS dysfunction, mitochondrial protein misfolding, decrease of mitochondrial membrane potential or ATP depletion [36]. Hence, it is likely that the hemocyte targeted UQCR-C1 knockdown induces the UPRmt.
Fig. 4
Knocking down UQCR-C1 in hemocytes leads to changes in the hemocyte transcriptomes. Venn diagrams show genes that are upregulated (A) and downregulated (B) upon UQCR-C1 knockdown in hemocytes, and heatmaps illustrate differential expression (as log2FC to wGD hemocytes) of selected genes specific to all samples of UQCR-C1 knockdown hemocytes, or unique to specific timepoint/treatment. Differentially expressed genes were grouped according to Flymine GO term pathway enrichment analysis. Asterisks indicate statistically significant difference in gene expression compared to wGD control. Gene symbols written in red encode mitochondrial proteins
Among the commonly downregulated genes, the only significant GO term was the cellular compartment term “extracellular region” (Additional File 8). However, manual curation of the data revealed downregulation of multiple innate immune response-related genes in all of the UQCR-C1 knockdown samples, including the extracellularly located modular serine protease (modSP) and Gram-positive Specific Serine protease (grass), both involved in the activation of the Toll signaling pathway (Fig. 4B; Additional File 6). As a comparison, under control conditions, wasp infection induced the upregulation of a large number of genes involved in the humoral immune response, including AMPs such as multiple Bomanins (also known as Immune-induced molecules, IMs), Daisho, Drosomycin (Drs), Cecropin A1 (CecA1) and Attacin B (AttB). Furthermore, Toll pathway inducer spatzle (spz) and its activators Spatzle-Processing Enzyme (SPE) and easter (ea) were all upregulated, as was Toll (Tl) itself (Additional Files 6 and 8). The downregulation of a set of humoral immune genes in response to UQCR-C1 knockdown could be explained by the presence of an immuno-suppressive mechanism aimed at reducing excessive inflammation response brought on by the activation of cell-mediated innate immunity (shown as the upregulation of lamellocyte-enriched marker genes), which could be especially important in immune-challenged animals. A group of genes involved in fatty acid metabolism was also downregulated, possibly signifying that there is a switch in cell metabolism, and that the UQCR-C1 knockdown hemocytes might rely on glycolysis for energy production.
In addition to this core set of differentially expressed genes, we looked at the changes occurring only at a certain timepoint or only when hemocytes were derived from wasp-infected larvae. At the early timepoint, knocking down UQCR-C1 led to an upregulation of an additional set of chaperones, heat shock proteins and protein folding-related genes, both cytosolic and mitochondrial (Fig. 4A; Additional File 6). This indicates that a stress response to UQCR-C1 knockdown is launched already at 90 h AEL. The downregulated genes included an additional group of fatty acid metabolism-related genes, again highlighting early changes in cellular metabolism.
At the late timepoint, there was an upregulation in fatty acid metabolism genes, including some that were downregulated at the early timepoint (Fig. 4A; Additional File 6). Multiple gluconeogenesis-related genes were upregulated, possibly to produce more free glucose to fuel the glycolytic pathway. Several genes connected to mitochondria and mitochondrial maintenance were upregulated, including those involved in mitophagy and mitochondrial calcium import, again suggesting a mitochondrial stress response. Among the downregulated genes was a group of innate immune response genes, mostly encoding AMPs such as several Attacins and Cecropins (Fig. 4B).
In UQCR-C1 knockdown wasp-infected samples, compared to wasp-infected controls, a large group of genes involved in mitochondrial translation and mitochondrial transport were upregulated (Additional Files 6 and 8, Fig. 4A). As mentioned above, increased expression of genes in these groups is a feature of the UPRmt [36, 37]. Another large group of genes that was upregulated was that of genes involved in cell cycle regulation. Among the downregulated genes of interest were genes related to glycogen metabolism, gluconeogenesis and fatty acid metabolism (Fig. 4B).
OXPHOS perturbation-induced hemocyte activation is not dependent on ROSThe disruption of OXPHOS, especially complexes I and III, leads to an increased electron escape, creating increased amounts of reactive oxygen species (ROS) (reviewed in [38]). An excess of ROS can overload the antioxidant enzymes, leading to the oxidation of macromolecules [39]. We looked at the expression levels of genes involved in ROS production and detoxification in our RNA sequencing data. Superoxide, the precursor of most types of ROS, is degraded by the superoxide dismutases (Sod), which catalyze its conversion to hydrogen peroxide (H2O2). H2O2 is further broken down by Catalase (Cat) in the cytosol [40]. Expression of neither Sod1, Sod2 and Sod3, nor Cat, was significantly affected by UQCR-C1 knockdown (Additional File 9: Fig. S5A). Immune-regulated Catalase (Irc) was not affected by UQCR-C1 knockdown in uninfected samples, but was downregulated in UQCR-C1 knockdown hemocytes upon wasp infection when compared to wasp-infected control hemocytes (Additional File 9: Fig. S5A). In contrast, under the control conditions Irc was upregulated upon wasp infection (Additional File 6), suggesting a suppression of this induction in wasp-infected UQCR-C1 knockdown hemocytes. Peroxiredoxins (Prx) are sensitive to the levels of H2O2, and function in neutralizing it and other peroxides in the cell [41]. Cytosolic Prx2 (also known as Jafrac1) was consistently upregulated by UQCR-C1 knockdown, while others were either downregulated or their expression did not change (Additional File 9: Fig. S5A). Peroxidasin (Pxn) was not affected, but globin1 (glob1), also encoding a peroxidase, was upregulated at the early and late timepoints after UQCR-C1 knockdown (SFig. 5A) as well as after wasp infection in control hemocytes (Additional File 6), indicating that it might be specific for activated hemocytes or lamellocytes. H2O2 can also function as a signaling molecule by oxidizing cysteine residues on proteins, and can be produced via the NOX/DUOX system on demand [42]. NADPH oxidase (Nox) and Dual oxidase (Duox) were downregulated at the early timepoint with UQCR-C1 knockdown. Multiple Glutathione transferase D and -E (GstD and GstE) genes were upregulated at both early and late timepoints with UQCR-C1 knockdown, and some also after wasp infection (Additional File 9: Fig. S5A). Glutathione transferases act as a general defense system against various genotoxic and cytotoxic electrophilic compounds, including ROS, by catalyzing conjugation of glutathione to these compounds [43]. As mentioned above, Gst expression is also a feature of the UPRmt. Taken together, the RNA sequencing data show some upregulation of ROS scavenging system genes, indicating that there may be a modest increase in ROS upon UQCR-C1 knockdown in hemocytes.
To further study the connection between the immune cell activation phenotype and ROS, we used three approaches. First, ROS was measured in the OXPHOS cI, cIII and cV knockdown hemocytes using the CellROX™ green probe. While feeding antioxidant N-acetylcysteine (NAC) to the larvae reduced the levels of ROS in hemocytes (Additional File 9: Fig. S5B), knocking down the OXPHOS cI, cIII or cV did not change the average CellROX green signal intensity (Additional File 9: Fig. S5B’-C) or the proportion of CellROX-positive hemocytes (Additional File 9: Fig. S5C’). Second, we checked whether knocking down antioxidant-encoding genes in hemocytes could phenocopy the effects of UQCR-C1 knockdown on the formation of melanotic nodules, hemocyte activation and the immune response against wasps. To this end, we knocked down genes encoding cytosolic Sod1, the mitochondrial Sod2 [44, 45] or Cat in hemocytes. This did not induce the formation of melanotic nodules (0% prevalence in Sod1 and Sod2 knockdowns, 1% in Cat knockdown, data not plotted) or lamellocyte formation (Additional File
留言 (0)