
記住我
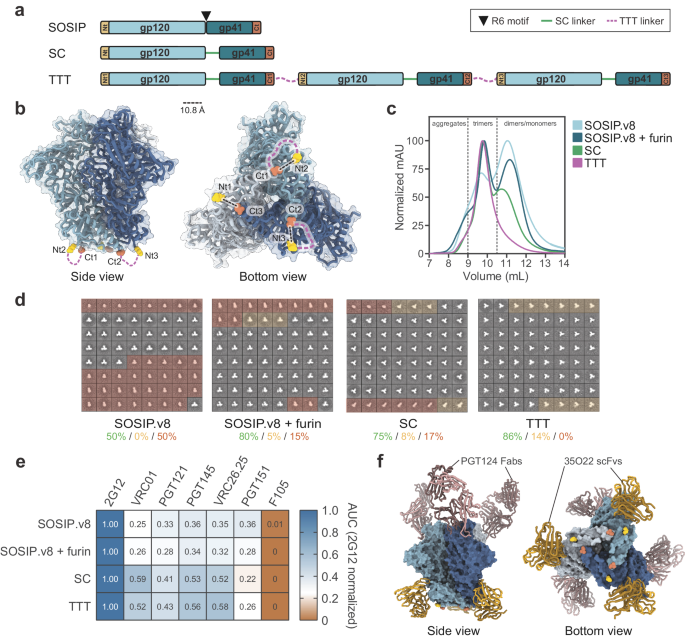
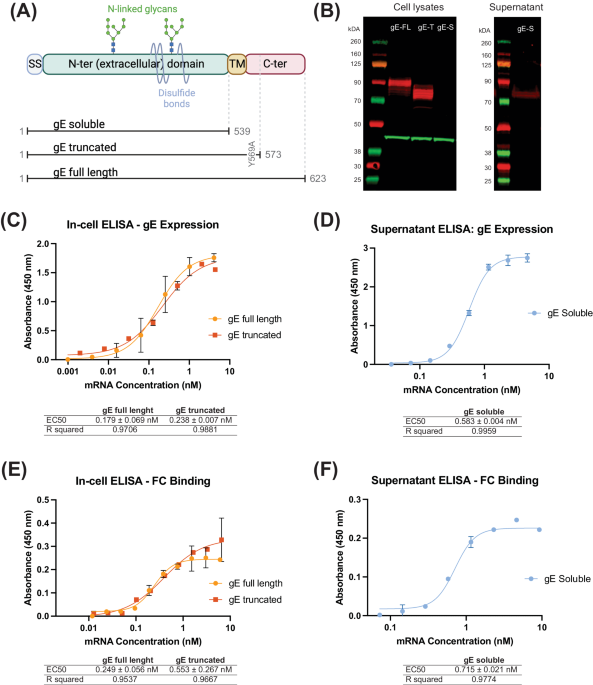
As reported elsewhere6, through personalized genotyping a total of 9 different IGHV1-2 genotypes were found among the 48 IAVI G001 trial participants (Fig. 1a, b). These consisted of combinations of the known alleles *02, *04, *05 and *06, in addition to allelic variant, IGHV1-2*02_S4953, that is distinct at the nucleotide level but encodes the same amino acid sequence as the *02 allele. For the subsequent analyses in the current study, we therefore classified *02_S4953 as *02. A structural view and amino acid sequence alignment of the various IGVH1-2 alleles is shown in Supplementary Fig. 1. The frequency of each allele in the 48 trial participants showed that IGHV1-2*04 was most common, followed by IGHV1-2*02 (Fig. 1c). The one participant that did not produce a detectable VRC01-class response was found to be IGHV1-2 genotype *05/*06, i.e. lacking one of the required *02 or *04 VRC01-class alleles, at least one of which were present in the genotype of all other trial participants (Fig. 1b and Supplementary Table 1)6.
Fig. 1: IGHV1-2 genotype, allele, and pre-vaccination IgM repertoire distributions for IAVI G001 trial participants.
a The IGHV1-2 allele content in each study participant was determined by sequencing bulk IgM libraries and inferring the IGHV allele content in each case with IgDiscover. Quantitative analyses of mRNA expression levels and HCDR3 frequencies followed. b Number of each genotype and c number of each allele, for all trial participants (n = 48). d Number of vaccine recipients per group, out of 18, with each allele (*02, *04, *05, and *06), with the *02 variant *02_S4953 classified as *02. P-values are based on a Fisher’s exact test with values >0.05 marked as not significant (NS). For all 48 participants, pre-vaccination e mRNA expression frequencies and f unique HCDR3 frequencies for each IGHV1-2 allele are shown as points color-coded by allele and grouped by homozygous and heterozygous genotype. Each point represents a trial participant, with heterozygous participants represented by two points. Thick lines are median values and boxes are the 25% and 75% quantiles. g Correlation between mRNA count and unique HCDR3 count; h Ratio of unique HCDR3 count to mRNA count versus mRNA count; i Correlation between frequency of mRNA expression and frequency of unique HCDR3s; and j Ratio of the unique HCDR3 frequency to the mRNA frequency versus the mRNA frequency, are shown for pre-vaccination repertoires. Points are shape- and color-coded by IGHV1-2 allele as shown in the legend of (g–j). Pearson correlation coefficients (r) for counts and frequencies are shown in (g and i), respectively. In panels h and j, the solid line is the median ratio, and the shaded region shows the inter-quartile range.
We observed that the distributions of *02 and *04 alleles between dose groups were substantially uneven (Fig. 1d). In the high dose group, 72% of participants (13 of 18) had an *02 allele, compared to 28% (5 of 18) in the low dose group (P-value = 0.02). Conversely, in the low dose group, 94% of participants (17 of 18) had an *04 allele, compared to 44% (8 of 18) in the high dose group (P-value = 0.003). The *05 and *06 alleles were less prevalent and had similar frequencies between dose groups (Fig. 1d). The imbalance in *02 and *04 between dose groups potentially impacted the relative strengths of the VRC01-class responses, because a previous study had shown that the frequency of eOD-GT8-specific naive precursors in the germline repertoire was higher for individuals encoding the *02 allele compared to those encoding the *04 allele15. Therefore, we hypothesized that the dose effect observed in this trial depended on IGHV1-2 allelic differences between the groups.
IGHV1-2 allele frequencies differ in the germline repertoireWe calculated the mRNA expression frequencies of different IGHV alleles in the naive repertoire for each trial participant by counting the per-allele unique molecular identifiers (UMIs) introduced during the cDNA synthesis of IgM libraries (Fig. 1a and Supplementary Data 1). The frequencies for each IGHV1-2 allele are shown separately for homozygous and heterozygous genotypes in Fig. 1e. The mean per-allele mRNA expression of *02 was similar in homozygous and heterozygous individuals, at 3.1% (95% CI: 2.7–3.6%) and 3.3% (95% CI: 2.9–3.8%), respectively (Supplementary Table 2). Usage of *04 was ~4-fold lower, at 0.9% (95% CI: 0.7 to 1.1%) in homozygous participants and 0.7% (95% CI: 0.6 to 0.8%) in heterozygous participants (Supplementary Table 2). Comparison of the per-allele frequencies between homozygous and heterozygous participants suggested that allele usage was proportional to zygosity, with homozygotes having approximately twice the usage of *02 or *04 than heterozygotes (Fig. 1e and Supplementary Table 3). Alleles *05 and *06, which do not have the required VRC01-class binding motif, had very low usage, 0.09% (95% CI: 0.03–0.14%), and intermediate usage, 2.4% (95%CI: 1.9–3.0%), respectively (Supplementary Table 2). The differences in relative frequencies of IGHV1-2 allele usage we observed are consistent with data from two previous studies15,16 and from recent analysis of allele frequencies of expressed IGHV genes within a collection of previously published IgM libraries17,18.
We then counted the number of unique heavy chain complementarity determining region 3 (HCDR3) sequences within IGHV1-2 mRNAs, a measure of the number of unique IGHV1-2 B cells in each personalized library (Fig. 1a and f and Supplementary Data 1). We found that, regardless of IGHV1-2 allele, the number of unique IGHV1-2 HCDR3s was proportional to the number of IGHV1-2 mRNA molecules (Fig. 1g, h), and the frequencies in the repertoire were also proportional (Fig. 1i, j). Indeed, ratios of unique HCDR3 counts to mRNA counts (Fig. 1h), a measure of 1/(B cell receptor cell surface density), were similar for *02 and *04 alleles (median 0.23 and 0.24, respectively; P-value for difference = 0.53), which suggests that the surface density was not appreciably different between these two alleles. Additionally, ratios between HCDR3 frequency and mRNA frequency for *02 and *04 (Fig. 1j) were also similar (0.93 and 0.96, respectively; P-value for difference = 0.19), which also suggested that surface density was similar in the two cases. Thus, different IGHV1-2 alleles had different frequencies of unique precursor B cells, with *02 higher than *04 (Fig. 1f). Furthermore, *02 or *04 homozygotes had approximately twice the frequency of allele-specific unique precursors as heterozygotes (Fig. 1f). Higher bnAb-precursor frequency has been shown to lead to higher bnAb-precursor-derived vaccine responses in mouse models19,20,21. Therefore, the higher frequency of unique IGHV1-2 precursors in *02 compared to *04 individuals suggested that the stronger VRC01-class responses in the IAVI G001 high dose group could be due at least in part to the greater representation of *02 in that group. Taken together, these results supported including the *02 and *04 allele counts as independent predictors for the frequency of vaccine-induced VRC01-class IgG B cells in our statistical models.
One model best explains the difference in VRC01-class responses between dose groupsTo look for genotype-specific effects in the vaccine response data, we first analyzed differences in post-vaccination VRC01-class B cell frequencies among trial participants that were *02/*02, *02/*04, *04/*04, or *04 heterozygous with either of the non-productive *05 or *06 alleles, and we found no significant differences after adjusting for multiple comparisons (Supplementary Table 4). Similarly, we compared across dose groups with sufficient data for two genotypes: (i) *02/*04 and (ii) *04/*05 or *04/*06 (pooled genotype), and we found that neither comparison was significant (Supplementary Table 5). However, the small numbers of participants within each genotype and dose group meant that the analysis had low sensitivity. To increase sensitivity to detect genotype-specific effects, we used statistical modeling to analyze pooled data under the mechanistic assumption that allele effects were additive.
We modeled the VRC01-class response frequency using four candidate models. Each model was defined by one or more frequency parameters:
Model 1 (Null): A single frequency for all vaccine recipients.
Model 2 (Dose): Two frequencies, one for the low dose and another for the difference between high and low dose.
Model 3 (Allele): Two frequencies, one each for the *02 and *04 alleles.
Model 4 (Full): Three frequencies, one each for the *02 and *04 alleles at low dose, and another for the allele-independent difference between high and low dose.
As described in Leggat et al.6, for all trial participants (n = 48), the frequencies of VRC01-class BCRs were measured for three sample types at seven sample collection time points: (i) PBMC IgG memory B cells (MBCs) at weeks 4, 8, 10, and 16; (ii) lymph node IgG germinal center (GC) B cells at weeks 3 and 11; and (iii) PBMC IgD- plasmablasts (PBs) at week 9. Here, we modeled count data of VRC01-class BCRs using a quasi-Poisson generalized linear model. The models were fit separately for each of the seven sample collection time points and ranked based on a Quasi-likelihood version of Akaike’s second-order information criterion (QAICc)22.
The QAICc model selection criterion ranked the models consistently at all four MBC timepoints, with the following best-to-worst order: (i) Allele; (ii) Full; (iii) Dose; and (iv) Null (Supplementary Fig. 2a and Supplementary Table 6). This model ranking indicated that the differential distributions between the dose groups of IGHV1-2 alleles *02 and *04 better explained the VRC01-class B cell frequencies post vaccination than either dose alone or a simple overall average frequency among vaccine recipients estimated by the Null model. Furthermore, the Allele model ranked higher than the Full model at all MBC timepoints, which indicated there was no detectable dose effect after accounting for the allele effect. At the week 3 GC and week 9 PB time points, the Allele model was also selected as the best model, but the ranking of the remaining models changed (Supplementary Fig. 2a and Supplementary Table 6). For the week 11 GC samples the Null model ranked highest, suggesting that neither dose nor allele adequately explained the observed variation in those samples.
Parameter estimates from the Allele model were consistently higher for the per-allele contribution of *02 to the VRC01-class response than for the contribution of *04 (Supplementary Fig. 2b and Supplementary Table 7). The Full model estimated similar per-allele effects as the Allele model but also estimated effects for high versus low dose that were close to zero and had confidence intervals that included zero in all cases (Supplementary Fig. 2b and Supplementary Table 7). Thus, the data provided no support for a true dose effect, and these results suggested that the count of *02 and *04 alleles best explained the variation in the VRC01-class B cell response to eOD-GT8 60mer.
We then compared experimentally measured VRC01-class frequencies to the Allele-model-estimated mean VRC01-class responses by genotype (Fig. 2 and Supplementary Table 8). To make comparisons in an ordered manner, we ranked IGHV1-2 genotypes based on the frequency of IGHV1-2 mRNA expression in the naive repertoire: first, we grouped genotypes that had a single *05 or *06 allele together since these alleles lack the necessary VRC01-class motif; next, we ranked the genotypes based on our observation that *02 precursors were approximately four-fold more common than *04 precursors. This resulted in our most-to-least favorable ranking of genotypes for induction of a VRC01-class response: (i) *02/*02; (ii) *02/*04; (iii) *02/*05 or *02/*06; (iv) *04/*04; and (v) *04/*05 or *04/*06. The genotype-specific medians of the experimentally measured VRC01-class frequencies generally followed this same ordering (Fig. 2), and the model-estimated mean VRC01-class response also followed this ordering, except for the week 10 MBC timepoint where the order of *04/*04 and *02/*05 or *02/*06 was reversed (Fig. 2 and Supplementary Table 8). Hence the model captured the dependence of the VRC01-class response frequency on genotype. However, there was substantial heterogeneity in the responses that was not explained by genotype (Fig. 2). This variation was potentially attributable to other factors that can influence the strength of immune responses, including but not limited to sex, age, additional genetic factors, and immune history. Nevertheless, our modeling indicated that the effect of dose was negligible after accounting for IGHV1-2 allele content.
Fig. 2: Model estimates and 95% confidence intervals (CIs) from the Allele model for each genotype and time point.
Estimates and CIs for the frequency of VRC01-class IgG B cells at each time point by genotype are shown as open diamonds and vertical lines, respectively. Thick lines are median values and boxes are the 25% and 75% quantiles. Experimentally measured frequencies for each participant are shown as color- and shape-coded points by dose group as indicated by the legend. Genotypes containing the *05 and *06 alleles are grouped together (e.g., *02/*05 or *02/*06), because the estimated mean response from the Allele model depends only on the count of *02 and *04 alleles. Week 11 germinal center (GC) results are not shown since the Null model ranked higher than the Allele model for that sample time point.
*02 has higher frequencies in both the naive repertoire and the modeled VRC01-class responseFrom the Allele model, we computed the relative contribution of *02 versus *04 alleles to the post-vaccination VRC01-class B cell frequency at each time point, and we found ratios between 1.7 and 4.4 (Fig. 3a and Supplementary Table 9). The confidence intervals in all cases included 1.0, consistent with equal contributions from *02 and *04. However, when we computed differences, rather than ratios, in post-vaccination VRC01-class B cell frequencies, we found significantly greater contributions from *02 compared to *04 (Supplementary Table 10; differences significant for all timepoints except week 10). This suggested that the true ratios were >1. In the pre-vaccination IgM (naive) repertoire of vaccine recipients, we computed ratios for *02 versus *04 mRNA usage of 3.9 (95% CI: 3.0 to 5.3) among homozygous individuals and 4.2 (95% CI: 3.3 to 5.1) among heterozygous individuals, both of which were significantly >1.0 (P-values of <0.0001 and 0.0001, respectively) (Fig. 3b and Supplementary Table 11). Ratios of the frequencies of unique HCDR3s using *02 versus *04 in the pre-vaccination repertoire were similar to the mRNA frequency ratios (Fig. 3c and Supplementary Table 11), more directly indicating higher *02 naive B cell frequencies. Overall, mRNA usage and B cell frequency were higher for *02 than for *04 in the personal naive repertoires, and *02 was higher than *04 in the Allele-model-determined contributions to vaccine-induced VRC01-class B cell frequencies.
Fig. 3: Relative contribution of *02 versus *04 alleles to Allele-model-derived post-vaccination VRC01-class frequency, pre-vaccination IGHV1-2 mRNA expression level, and post-vaccination BCR assignments.
a Allele model estimates for the relative contribution of *02 versus *04 (as a ratio) to the post-vaccination VRC01-class frequency are shown with 95% confidence intervals (CIs) for germinal center (GC) B cell, memory B cell (MBC), and plasmablast (PB) samples taken at the indicated week (Wk) after first vaccination. b Experimentally measured pre-vaccination ratios of *02 to *04 mRNA expression levels for homozygous or heterozygous genotypes. For homozygotes, the ratio of means and CI for *02 and *04 individuals is shown. For heterozygotes, ratios for each individual are shape- and color-coded (N = 4 for low dose; N = 4 for high dose), and the overall mean ratio and CI are shown in black. c Experimentally measured pre-vaccination ratios of *02 to *04 unique HCDR3 frequencies for homozygous or heterozygous genotypes. Homo- and heterozygote data are displayed as in (b). d Ratio of *02 to *04 usage for germline allele assignments among post-vaccination BCRs recovered from eight vaccine recipients known to be heterozygous for *02 and *04 by pre-vaccination genotyping are shape- and color-coded (N = 4 for low dose; N = 4 for high dose), and the overall mean ratio and CI are shown in black.
*02 has higher frequencies among allele assignments for post-vaccination BCR sequencesConsidering that VRC01-class responses from *02/*04 heterozygous individuals involved direct competition between the two alleles, we assessed frequencies of VRC01-class post-vaccination BCRs with allele assignments of *02 or *04 from *02/*04 heterozygous participants. For each VRC01-class BCR, the IGHV1-2 allele was bioinformatically assigned, accounting for the personal IGHV1-2 genotype information, as described in Leggat et al.6. From 873 post-vaccination BCR sequences of VRC01-class IgG (MBC or GC) or IgD- (PB) B cells from eight *02/*04 heterozygous vaccine recipients, we computed the per-participant ratio of *02 to *04 as the assigned germline allele (Fig. 3d and Supplementary Table 12). Only four of 873 (0.46%) BCR assignments were ambiguous, hence allele ambiguity did not meaningfully affect our calculations. The overall median ratio of *02 to *04 was 7.3 (range, 2 to 17.7) (Supplementary Table 12), which was significantly >1.0 (P-value = 0.004). The ratio of *02 to *04 BCRs varied widely (0 to infinity) across different post-vaccination time points but was >1.0 in 89% (42/47) of cases (Supplementary Table 13). The observed BCR usage ratios (Fig. 3d and Supplementary Tables 12–13) were generally higher than the model-derived usage ratios (Fig. 3a and Supplementary Table 9) and the naive repertoire mRNA and HCDR3 frequency ratios (Fig. 3b, c and Supplementary Table 11). We tested for a dose effect in the data in Fig. 3d but found no significant effect (P-value = 0.154). IGHV1-2*02 differs from *04 by a single nucleotide (A*04/T*02, SNP rs112806369) and a single amino acid (Arg66*04/Trp66*02, Kabat residue numbering) (10,15,23). We documented several occurrences of the Arg66*04→Trp66*02 mutation, which is favorable for VRC01-class maturation, in post-vaccination VRC01-class BCRs from *04 homozygous individuals6. Thus, occurrence of this mutation might have inflated the *02 to *04 germline allele ratio in BCRs of heterozygous individuals shown in Fig. 3d. Overall, the *02 to *04 ratios among BCRs from heterozygotes were consistent with the conclusions of the Allele model in that both indicated stronger VRC01-class responses from the *02 allele. Those findings, combined with the fact that the naive repertoire mRNA and HCDR3 ratios demonstrated higher expression and B cell frequency for the *02 allele, suggested a simple potential explanation for the superiority of *02 over *04 for VRC01-class responses, namely that the higher frequency of *02-using B cell precursors translated into higher post-vaccination VRC01-class responses.
Naive repertoire predicts outcomeTo test the hypothesis that stronger VRC01-class responses resulted from higher B cell precursor frequencies, we looked for correlations between the VRC01-class response and the total frequency of IGHV1-2*02 or *04 B cells in the naive repertoire. Pooling across dose groups (Fig. 4), we found significant correlations at each time point, excluding week 11, with correlation coefficients ranging from 0.4 to 0.6 (Fig. 4, P-values ranging from 0.04 to 0.0003). Analyzing dose groups separately, both dose groups showed positive correlations at MBC timepoints, but the correlations were stronger and statistically significant only for the high dose group. At GC timepoints, we found significant positive correlations only in the low dose group (Supplementary Fig. 3). The difference in correlation strength could reflect a dose effect but could also be explained by the fact that the high dose group, which was over-represented by *02, generally had stronger VRC01-class responses with higher dynamic range and had higher standard deviation of HCDR3 frequency perhaps due to its wider range of genotypes. Whatever the explanation, the significant positive correlations in the pooled data demonstrated that the strength of the VRC01-class response increased monotonically with the total frequency of IGHV1-2*02 and *04 naive precursors. This finding, together with the observation that the experimentally measured naive precursor frequency was higher for *02 than for *04, indicated that the stronger VRC01-class responses for *02 were most likely due simply to higher naive B cell frequencies for *02. Thus, experimental data provided independent corroboration for our statistical modeling.
Fig. 4: Correlations between pre-vaccination IgM unique HCDR3 frequency (IGHV1-2*02 or *04) and the percent VRC01-class B cell response by visit.
Points are shape- and color-coded as shown in the legend. Spearman correlation coefficient (r) and P-values are displayed for each time point.
Alleles affect precursor affinitiesThe fundamental hypothesis of the germline-targeting priming strategy is that vaccine antigen affinity and avidity for rare bnAb-precursor B cells strongly impacts whether or not the vaccine can trigger GC and memory responses from those precursors. Hence, while our above analyses focused on precursor frequency, it was also important to consider whether affinity differences between alleles could explain the different response outcomes. To begin, we investigated the effect of the IGHV1-2 *05 and *06 alleles on the affinity of eOD-GT8 for VRC01-class precursors. Alleles *05 and *06 both possess Arg50 instead of Trp50, one of the critical VRC01-class germline residues that interacts with HIV gp120. We produced W50R variants of VRC01-class precursors that were originally *02 or *04, including inferred germline (iGL) precursors for two bnAbs (VRC01 and N6), iGLs for four post-vaccination BCRs in the IAVI G001 low dose group6, and four human naive precursors isolated by prior B cell sorting studies of HIV-unexposed individuals24,25. We then assessed eOD-GT8 binding affinity for the original and W50R-variant iGL precursors using surface plasmon resonance (SPR). The original precursors all bound to eOD-GT8, with a median KD of 120 nM, whereas only one of the W50R variants had detectable affinity, and the overall median KD was ≥100 µM (Fig. 5a and Supplementary Data 2). The only W50R binder (KD, 4.0 µM) derived from the highest affinity original Ab, VRC01 iGL (KD, 49 pM), with an ~80,000-fold loss in affinity due to W50R. These results provided an explanation for why eOD-GT8 60mer failed to induce VRC01-class responses in the one *05/*06 participant.
Fig. 5: Affinity analyses of VRC01-class IGHV1-2 allele variants.
a Affinities of VRC01-class precursors from two bnAb iGLs, four human naive precursors, and four iGLs from week 3 post-vaccination BCRs from IAVI G001, with original *02 or *04 Abs on the left, and W50R variants of those Abs on the right. b Affinities of *02 (N = 71) and *04 (N = 47) iGLs from post-vaccination BCRs from IAVI G001. c Affinities of *02 VRC01-class iGL antibodies from IAVI G001 and for *04 (R66W) variants of the same antibodies (N = 28 each). Lines connect matched Ab variants. In (a, b), horizontal lines indicate median and interquartile range. In (a–c), all affinities were measured for eOD-GT8 monomer analyte, and all iGLs from IAVI G001 are from the low dose group.
To complement our findings above that VRC01-class *02 precursors had higher frequencies than *04 in IAVI G001, we evaluated whether the two precursor populations had differing affinities for eOD-GT8. We first re-analyzed the Leggat et al. data on eOD-GT8 affinities for inferred germline (iGL) variants of post-vaccination VRC01-class BCRs recovered from the low dose group. These iGLs were overwhelmingly (96%) derived from memory BCRs after the first or second vaccination, hence they represented precursors to vaccine responses that survived GC competition and became memory B cells in the blood. Leggat et al. reported statistics on all VRC01-class iGLs (median KD, 119 nM; N = 118), but here we examined the data for *02 and *04 iGL precursors separately. We found that *04 iGLs, with median KD of 59 nM and interquartile range of 5.8 nM to 480 nM (N = 47), had approximately six-fold higher affinities than *02 iGLs (median KD, 380 nM; interquartile range, 29 nM to 2.9 µM; N = 71) (Fig. 5b and Supplementary Data S2). However, using a repeated measures test to account for the fact that affinities for different antibodies from the same participant are potentially correlated (the N = 47 *04 Abs came from 14 participants, and the N = 71 *02 Abs came from 5 participants), the six-fold difference was not significant (p-value, 0.38). Furthermore, we tested directly whether *04 precursors had inherently higher affinity than *02 precursors as a consequence of the R66W mutation that distinguishes *02 (Arg66) from *04 (Trp66). Although this mutation is not located within the region of direct antibody-antigen contacts, the spatial location of this mutation suggested that an indirect (allosteric) effect on affinity was possible (Supplementary Fig. 1). To test for such an effect, we produced 28 different *02 VRC01-class iGL antibodies from the IAVI G001 low dose group, along with *04 (R66W) variants of the same antibodies, and we measured their affinities for eOD-GT8 monomer by SPR (Fig. 5c and Supplementary Data 2). We found that the original *02 iGLs (median KD, 310 nM; interquartile range, 30 nM–8.3 µM; N = 28) had indistinguishable affinities to their *04 variants (median KD, 280 nM; interquartile range, 23 nM–10 µM; N = 28; repeated measures test p-value, 0.64). We concluded that neither allele had a precursor affinity advantage over the other. Therefore, affinity differences between alleles could not explain the different response outcomes between the vaccine groups.
留言 (0)