
記住我



To assess the animals’ anxiety/avoidance-like behavior, elevated plus maze (EPM) and open field (OF) tests were employed.
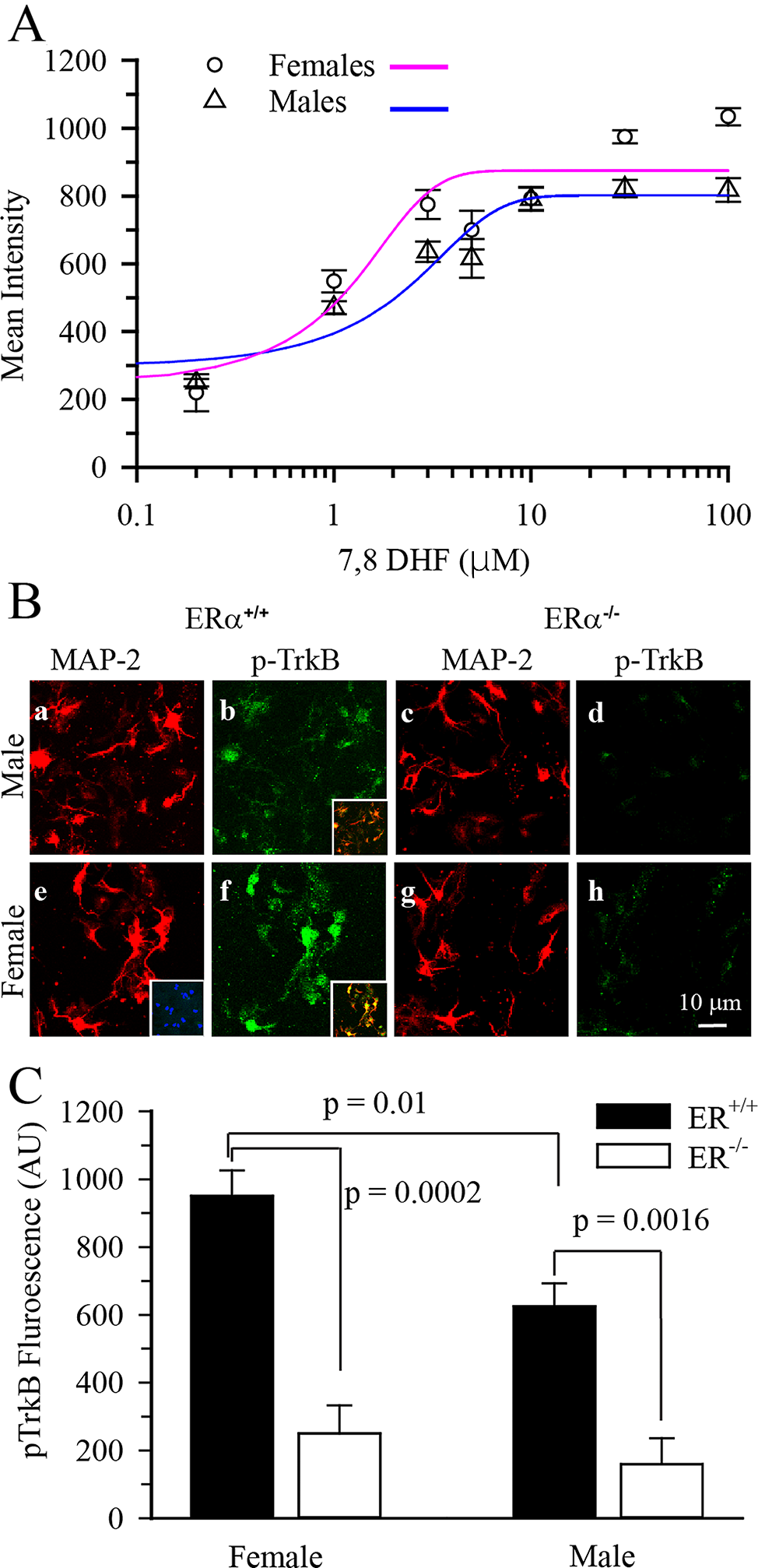
On EPM: A comparison between the male and female anxiety index revealed a significant sex effect (F(1,44) = 5.2, p = 0.0275, η2 = 0.1050). SPS males displayed a significantly higher anxiety index than SPS females. No sex differences were observed in the anxiety indices of the unstressed controls (Fig. 2A). Irrespective of sex, animals in the SPS group showed divergent responses to the traumatic stress, with females displaying clearer separation (Fig. 2A). The anxiety index of the SPS treated males ranged from 0.7–1, whereas that of the females ranged from 0.5–0.9. Due to the observed variations in both sexes, animals in the SPS group with an anxiety index two standard deviations above the mean of their respective controls were subdivided into SPS-susceptible (SPS-S) subgroups, while the rest were grouped as SPS-resilient (SPS-R) [39, 40, 52, 53]. Following group subdivisions, significant group (F(2,42) = 30.92, p < 0.0001, η2 = 0.5955) and sex (F(1,42) = 12.03, p = 0.0012, η2 = 0.2220) effects were observed in the anxiety index with no interaction. Animals in the SPS-S subgroup of both sexes showed a significantly higher anxiety index than their respective controls and SPS-R subgroups (Fig. 2B). Interestingly, SPS-R females displayed a significantly lower anxiety index than SPS-R males (Fig. 2B). When the percentage duration in the open arms was calculated, a significant group effect (F(2,42) = 27.48, p < 0.0001, η2 = 0.5667) and an interaction between sex and group (F(2,42) = 3.410, p = 0.0424, η2 = 0.1392) were observed. Animals in the SPS-S subgroup, regardless of sex, spent significantly less time in the open arms of the maze compared to their respective controls and SPS-R subgroups. SPS-R females spent significantly more time in the open arms than SPS-R males (Fig. 2C). Additionally, the frequency of open arm entries showed significant group (F(2,42) = 26.63, p < 0.0001, η2 = 0.5591) and sex (F(1,42) = 14.7, p = 0.0004, η2 = 0.2592) effects, with no significant interaction. Male and female rats in the SPS-S subgroups displayed significantly fewer entries into the open arms than their respective controls and SPS-R subgroups. However, females showed more frequent entries into the open arms (OAs) than their respective male groups (Fig. 2D).
Fig. 2
Sex differences in anxiety-like behavior and social impairments following SPS. Anxiety-like behavior tested on EPM: A Anxiety Index before group subdivisions, B Anxiety Index following group subdivisions, C % duration in open arms (OA), D % entries into open arms (OA). Anxiety-like behavior tested on OF: E duration in the center of the arena, F number of entries into the center of the arena, G number of rears. Social behavior tested on SI: H time spent interacting with a juvenile rat of same sex, I number of interactions initiated by the test rat towards the juvenile rat. Each symbol represents the value for an individual animal (blue bars—males: control n = 10, SPS-R n = 7, SPS-S n = 7; pink bars—females: control n = 10, SPS-R n = 8, SPS-S n = 6)
On OF: The time spent in the center of open field showed significant group and sex interaction (F(2,42) = 3.567, p = 0.0371, η2 = 0.1446). Male rats in the SPS-S subgroup spent significantly less time in the center compared to their controls, while no differences were observed among females. Additionally, control males spent significantly more time in the center of the arena than control females (Fig. 2E). Regarding the frequency of entries into the center, a significant group effect was found (F(2,42) = 4.482, p = 0.0172, η2 = 0.1757). SPS-S males displayed fewer entries into the center compared to control and SPS-R males. However, no differences were observed among females (Fig. 2F). An additional measure of anxiety-like behavior on the OF is rearing behavior [54]. The analysis of the number of rears on the open field demonstrated a significant group effect (F(1,42) = 10.29, p = 0.0026, η2 = 0.1962), with female controls showing more rears compared to control males (Fig. 2G).
On SI: Social interaction (SI) test was used to evaluate the level of active interaction between the test animals of each subgroup and a novel juvenile rat of the same sex. Analysis of the time spent interaction revealed significant group (F(2,42) = 9.513, p = 0.0004, η2 = 0.3115) and sex (F(1,42) = 48.25, p < 0.0001, η2 = 0.5342) effects with no interaction. Males in the SPS-S subgroup spent less time interacting with the juvenile rat compared to their controls and SPS-R subgroup. In females, however, differences were only evident between SPS-S and SPS-R subgroups. When the time spent interacting was compared between the sexes, males, irrespective of the groups, spent more time interacting than the females (Fig. 2H). Similarly, the number of interactions showed significant group (F(2,42) = 7.297, p = 0.0019, η2 = 0.2575) and sex (F(1,42) = 12.78, p = 0.0009, η2 = 0.2336) effects with no interaction. Male rats in the SPS subgroup initiated significantly fewer approaches towards the juvenile rats compared to their controls, whereas SPS-S females displayed fewer interactions compared to both SPS-R subgroup and unstressed controls. Additionally, SPS-R females demonstrated more frequent social interactions than SPS-R males (Fig. 2I).
Sex-specific differences in net weight gain following SPS exposureAs another measure of the stress response, animal body weight measurements were taken at the time of the SPS stressors, as well as 14 days afterward and the net weight gain from the day of the SPS stressors was calculated. While the group effect approached significance (F(2,42) = 3.083, p = 0.0563, η2 = 0.1273), two-way ANOVA revealed significant sex effect (F(1,42) = 142.1, p < 0.0001, η2 = 0.7711) and interaction between the two factors (F(2,42) = 5.483, p = 0.0077, η2 = 0.2061). Male rats in SPS subgroups gained significantly less weight than their unstressed controls, however the net weight gain of the SPS-S subgroup was lower than that of the SPS-R subgroup. Moreover, males in all groups gained more weight than did the females. No group differences were observed among the females (Fig. 3).
Fig. 3
Sex differences in net weight gain. All animals were weighed on the day of SPS (day 15) and two weeks later (day 31), and the net weight gain was calculated. Each symbol represents the value for an individual animal (blue bars—males: control n = 10, SPS-R n = 7, SPS-S n = 7; pink bars—females: control n = 10, SPS-R n = 8, SPS-S n = 6)
Sex-specific differences in adrenomedullary system of male and female rats before and after exposure to SPSTo assess differences in the stress response of male and female rats, the urinary epinephrine levels were measured before and 30 min into the immobilization step of SPS. A comparison of the basal urinary epinephrine levels showed significant group effect (F(2,31) = 7.212, p = 0.0027, η2 = 0.3173). Before exposure to SPS, male rats in the SPS-S subgroup displayed significantly higher levels of basal urinary epinephrine than their respective controls and the SPS-R subgroup. No differences were observed among the females (Fig. 4A). During the immobilization step of SPS, a significant group effect (F(1,22) = 13.49, p = 0.013, η2 = 0.3792) was observed, with the urinary epinephrine levels being higher in the SPS-S subgroups of both sexes compared to their respective SPS-R subgroups (Fig. 4B).
Fig. 4
Sex differences in urinary epinephrine levels before SPS and 30 min into the immobilization step of SPS. Urine samples were collected before SPS and 30 min into immobilization step of SPS, to measure urinary epinephrine levels of individual animals. A Basal relative epinephrine levels, B relative epinephrine levels 30 min into the immobilization step of SPS. Each symbol represents the value for an individual animal. Due to technical difficulties, urine samples were not collected from every single animal. Moreover, during the immobilization step urine samples were not collected from the controls as they were not exposed to stress. Each symbol represents the value for an individual animal. Before SPS (blue bars—males: control n = 5, SPS-R n = 5, SPS-S n = 6; pink bars—females: control n = 10, SPS-R n = 7, SPS-S n = 4). Following SPS (blue bars—males: SPS-R n = 6, SPS-S n = 6, and pink bars—females: SPS-R n = 8, SPS-S n = 6)
Sex-specific differences in the expression of tight junction protein in ventral hippocampus and medial prefrontal cortex following SPS exposureChanges in blood–brain barrier (BBB) permeability are frequently reported in mood disorders [55]. We indirectly evaluated the permeability of the ventral hippocampus (vHipp) and medial prefrontal cortex (mPFC) by quantifying the levels of the tight junction protein claudin-5, known as the gatekeeper of neurological functions [56]. In males, the SPS-S subgroup exhibited significantly lower expression levels of claudin-5 compared to SPS-R subgroup in vHipp (F(2,15) = 4.019, p = 0.04) and compared to unstressed controls in mPFC (H(3,18) = 10.18, p = 0.0019), suggesting increased BBB permeability (Fig. 5A,5C). While in the females, the SPS-R group displayed higher expression of claudin-5 than that of the unstressed controls in the vHipp (H(3,22) = 13.35, p = 9.480, p = 0.0044) and higher than that of both unstressed controls and SPS-S subgroup in mPFC (F(2,18) = 4.667, p = 0.0233). Notably, claudin-5 levels in the SPS-S females were comparable to those observed in the controls for both regions (Fig. 5B, D).
Fig. 5
Sex-specific alterations in claudin-5 expression in the brain following SPS exposure. Ventral hippocampus (vHipp) and medial prefrontal cortex (mPFC) of each animal were dissected and Western blot was performed to analyze the expression of tight junction protein claudin-5. Expression of claudin-5 in A vHipp of males and B females, C mPFC of males and D females. Representative Western blots are shown. The images have been previously published in Neurobiology of Stress [39]. The analysis was performed using one-way ANOVA, as the samples from males and females were not run simultaneously. The values are normalized to their respective controls taken as 1. Claudin-5 protein expression data in the vHipp. of females, and in the mPFC of males were non-parametrically distributed and were analyzed using Kruskal–Wallis test followed by FDR corrected multiple comparison test. The rest of the data passed the normality test and were analyzed using one-way ANOVA, followed FDR corrected multiple comparison test. Each symbol represents the value for an individual animal. vHipp (blue bars—males control n = 7, SPS-R n = 6, SPS-S n = 5; pink bars—females control n = 10, SPS-R n = 6, SPS-S n = 6);mPFC (blue bars—males control n = 7, SPS-R n = 6, SPS-S n = 5; pink bars—females control n = 8, SPS-R n = 7, SPS-S n = 6)
Pre-existing and trauma-induced differences in the gut microbial composition in male and female ratsAlpha diversity was higher in females than in males both before and after exposure to SPSBefore exposure to SPS. Analysis of alpha diversity using OTUs revealed significant effects of both group (F(2,32) = 3.489, p = 0.0426, η2 = 0.1793) and sex (F(1,32) = 551.2, p < 0.0001, η2 = 0.9459), with strong interaction between the two factors (F(2,32) = 4.427, p = 0.0201, η2 = 0.2162). While no group differences were observed in the OTU numbers among males, SPS-S females exhibited a significantly lower number of OTUs compared to SPS-R and control females. When the numbers were compared between the sexes, females, regardless of their groups, had significantly higher OTU counts than males (Fig. 6A).
Fig. 6
Sex differences in alpha diversity before and after SPS. Alpha diversity was measured using OTU counts A before SPS, B after SPS. Fecal samples collected from each rat were sent for 16S sequencing. Due to technical difficulties stool samples were not collected from every single animal. Each symbol represents the value for an individual animal. Before SPS (blue bars—males control n = 7, SPS-R n = 6, SPS-S n = 5; pink bars—females control n = 9, SPS-R n = 6, SPS-S n = 5), following SPS (blue bars—males control n = 7, SPS-R n = 6, SPS-S n = 4; pink bars—females control n = 8, SPS-R n = 8, SPS-S n = 5)
Following SPS exposure. Two-way ANOVA revealed significant sex (F(1,32) = 241.9, p < 0.0001, η2 = 0.8835) and group (F(2,32) = 3.740, p = 0.0347, η2 = 0.1895) effects in OTU counts. Similar to the findings of before SPS exposure, OTUs remained significantly higher in females compared to males across all three groups. However, a notable change was observed in females after SPS exposure. Contrary to the pre-SPS results, the SPS-S subgroup exhibited significantly higher OTU numbers compared to SPS-R and unstressed control groups (Fig. 6B).
Beta diversity showed distinct separations between the sexes before and after exposure to SPSBefore SPS exposure. The Aitchison distance matrix, used as a measure of beta diversity [57], resulted in a PCA plot that displayed a distinct separation between male and female unstressed control groups (Fig. 7A), as well as between the SPS-S subgroups (Fig. 7C). Interestingly, there was no discernible separation observed between SPS-R males and females, indicating a similarity in microbial diversity in these two groups (Fig. 7B).
Fig. 7
Sex differences in Beta diversity before and after SPS. Beta diversity was assessed using Aitchison distance. Before SPS A male vs female controls, B males vs females SPS-R subgroup, C males vs females SPS-S subgroup. After SPS: D male vs female controls, E males vs females SPS-R subgroup, F males vs females SPS-S subgroup. Due to technical difficulties stool samples were not collected from every single animal. Each symbol represents the value for an individual animal. Before SPS (blue bars—males control n = 7, SPS-R n = 6, SPS-S n = 5; pink bars—females control n = 9, SPS-R n = 7, SPS-S n = 5), following SPS (blue bars—males control n = 7, SPS-R n = 6, SPS-S n = 4; pink bars—females control n = 8, SPS-R n = 8, SPS-S n = 6)
Following SPS exposure. A clear separation between control male and female rats persisted in the PCA plot after two weeks (Fig. 7D). However, unlike the pre-SPS results, the SPS-R male and female subgroups now exhibited a clear separation (Fig. 7E), whereas no distinct separation was observed within the SPS-S subgroups (Fig. 7F).
Sex-specific differences were observed at the genus level in males and females before and two weeks following SPS exposureGenus-level analysis of the gut microbial composition revealed that a total of 68 genera were shared between males and females, yet ten genera were exclusively present in females, and one genus was present only in males both before and after SPS exposure (Fig. 8A).
Fig. 8
Sex differences in the gut microbial composition at genus level before and after SPS. A Venn diagram depicting shared and sex specific genera, B heat map of the genera which showed significant pre-existing group differences in each sex as well as in both sexes, C relative abundance of Lactobacillus, D relative abundance of Lachnospiraceae_Incertae_Sedis, E heat map of the genera which showed significant SPS-induced group differences in each sex and in both sexes, F relative abundance of Barnesiella. Due to technical difficulties stool samples were not collected from every single animal. Each symbol represents the value for an individual animal. Before SPS (blue bars—males control n = 6, SPS-R n = 5, SPS-S n = 5; pink bars—females control n = 9, SPS-R n = 6, SPS-S n = 7/6), following SPS (blue bars—males control n = 6, SPS-R n = 5, SPS-S n = 4; pink bars—females control n = 7, SPS-R n = 7, SPS-S n = 6)
Pre-existing differences: Among these 68 shared genera, sex-specific group differences were observed in males and females (Fig. 8B). However, two genera, namely, Lactobacillus, and Lachnospiraceae_Incertae_Sedis, showed significant group differences in both sexes (Fig. 8B). The relative abundance of the genus Lactobacillus showed significant sex and group interaction (F(2,32) = 11.52, p = 0.0002, η2 = 0.4182). In male rats, its abundance was significantly higher in the SPS-R subgroup than in the SPS-S subgroup, whereas in female rats, the abundance of Lactobacillus was significantly lower in the SPS-R subgroup than in the SPS-S subgroup and unstressed controls (Fig. 8C). When the abundance of Lactobacillus was compared between the sexes, female rats in the SPS-R subgroup showed significantly lower abundance than SPS-R males (Fig. 8C). Similarly, the relative abundance of the genus Lachnospiraceae_Incertae_Sedis showed a strong interaction between group and sex (F(2,31) = 6.833, p = 0.0035, η2 = 0.3067). SPS-S males had significantly higher abundance of Lachnospiraceae_Incertae_Sedis compared to SPS-R males, whereas in females, the opposite was observed, with SPS-S subgroup showing significantly lower abundance than SPS-R subgroup (Fig. 8D). Moreover, when the abundance was compared between the sexes, SPS-R males showed significantly lower abundance of Lachnospiraceae_Incertae_Sedis than SPS-R females, whereas SPS-S males showed higher abundance than SPS-S females (Fig. 8D).
Following SPS exposure. To assess how gut microbial communities were altered after SPS, differences in gut microbial composition were evaluated. As before SPS, several sex-specific group differences were observed, with 2 genera showing alterations in both males and females (Fig. 8E). Yet only differences in the genus Barnesiella showed significant interaction between sex and group (F(2,29) = 8.635, p = 0.0011, η2 = 0.3725). Following exposure to SPS, the abundance of genus Barnesiella was significantly higher in SPS-S males compared to SPS-R males, whereas in females it was significantly lower in SPS-S subgroup compared to SPS-R and unstressed controls (Fig. 8F). Comparison of the genus Barnesiella between the sexes, revealed higher abundance in SPS-R females than in SPS-R males, and significantly lower abundance in SPS-S females than in SPS-S males (Fig. 8F).
Sex differences in gut microbial predictive functionality before and after SPSNext, the predictive functional profiles of the microbes were evaluated, as the link between microbial taxonomic composition and metabolic response is not direct [58]. Analyzing microbial functionality goes beyond understanding the composition of gut microbiota. It enables the comprehension of not only the potential biological activities and metabolic pathways, but also how the composition of the gut microbiota may influence or contribute to specific diseases and physiological processes.
Before SPS exposure. Cellular processes, environmental and genetic information processing, human diseases, carbohydrate, lipid, terpenoid, and polyketide metabolism, and xenobiotic degradation showed significant alterations in the SPS-R and SPS-S subgroups in both sexes, although the pathways involved in each domain were distinct for males and females (Fig. 9A). On the other hand, pathways involved in amino acid metabolism and biosynthesis of other metabolites showed group differences only in males, whereas pathways involved in glycan biosynthesis and metabolism showed group differences only in females (Fig. 9B). The only common pathway between males and females was apoptosis in cellular processes, which showed a significant group effect (F(2,32) = 3.560, p = 0.0402, η2 = 0.1822) and interaction between sex and group (F(2,32) = 11.51, p = 0.0002, η2 = 0.4189). In males, the pathways involved in apoptosis were higher in SPS-R subgroup than in their controls and SPS-S subgroup, whereas in females, it was lower in both SPS subgroups than in the unstressed controls. Moreover, apoptosis was higher in SPS-R males than in SPS-R females (Additional file 1: Fig. S2A).
Fig. 9
Sex differences in the gut microbial predictive functionality before and after SPS. A Heat map of the microbial predictive functionality which showed pre-existing group differences in both sexes (right side: general functional pathways, left side: specific domains in each pathway), B heat map of the microbial predictive functionality which showed pre-existing group differences in males or females only (right side: general functional pathways, left side: specific domains in each pathway), C heat map of the microbial predictive functionality which showed SPS-induced group differences in both males and females. Due to technical difficulties stool samples were not collected from every single animal. Each symbol represents the value for an individual animal. Before SPS (blue bars—males control n = 7, SPS-R n = 6, SPS-S n = 5; pink bars—females control n = 8, SPS-R n = 7, SPS-S n = 5), following SPS (blue bar—males control n = 7, SPS-R n = 6, SPS-S n = 4; pink bar—females control n = 7/8, SPS-R n = 7/8, SPS-S n = 5/6)
Following SPS exposure. Five pathways were found to be common between the sexes (Fig. 9C). Among these pathways, Sphingolipid metabolism (F(2,32) = 11.08, p = 0.0002, η2 = 0.4098), and glycosphingolipid biosynthesis-globo series (F(2,32) = 10.15, p = 0.0004, η2 = 0.3889) exhibited significant group differences only. On the other hand, other glycan metabolism showed both group (F(2,32) = 17.06, p < 0.0001, η2 = 0.5162) and sex (F(1,32) = 8.328, p = 0.0069, η2 = 0.2062) effects, with all three pathways being significantly higher in the SPS-S subgroup compared to SPS-R and unstressed controls, regardless of sex (Additional file 1: Fig. S2B–D). Sporulation demonstrated a significant sex effect (F(1,32) = 15.82, p = 0.0004, η2 = 0.3182) with the pathway being higher in control and SPS-R males than in females (Additional file 1: Fig. S2E). Finally, bacterial invasion of epithelial cells exhibited significant sex (F(1,32) = 43.58, p < 0.0001, η2 = 0.5760) and group (F(2,32) = 4.668, p = 0.0166, η2 = 0.2254) effects, along with an interaction between the two factors (F(2,32) = 4.802, p = 0.015, η2 = 0.2309). Irrespective of their groups, females showed strikingly higher levels of bacterial invasion than males. Notably, SPS-R females displayed higher levels of bacterial invasion compared to SPS-S and unstressed control females (Additional file 1: Fig. S2F).
Sex-specific differences in gut–brain modules and gut–metabolic modulesTo expand the functional analysis and the interaction between gut microbiota and the brain towards more targeted functional annotation frameworks, gut–brain modules (GBMs) and gut–metabolic modules (GMMs) were examined to assess the functional capabilities of gut microbes. These modules represent specific functional pathways that have been identified in the literature as being linked to gut–brain communication or microbiota metabolism, respectively [59, 60]. GBMs specifically target molecules that have the ability to traverse both the intestinal epithelium and the blood–brain barrier, indicating their potential role in influencing brain function. On the other hand, GMMs focus on gut-specific bacterial and archaic metabolic processes, with a particular emphasis on anaerobic fermentation, which plays a vital role in gut health and overall microbial function.
Gut–brain module before and after SPS exposureBefore exposure to SPS. Three gut–brain modules were shared between the sexes (Fig. 10A). Acetate synthesis I showed significant interaction between group and sex (F(2,32) = 6.198, p = 0.0053, η2 = 0.2791). Its levels were significantly higher in SPS-R males compared to unstressed controls and SPS-S males as well as SPS-R females (Additional file 1: Fig. S3A). Acetylcholine synthesis showed significant sex effect (F(1,32) = 12.12, p = 0.0015, η2 = 0.2756), with interaction between sex and group (F(2,32) = 5.378, p = 0.0097, η2 = 0.2513). The module was higher in SPS-R males compared to SPS-S and unstressed control males. While no group differences were seen among the females, acetylcholine synthesis was higher in controls and SPS-S females than in their respective groups in males (Additional file 1: Fig. S3B). Similarly, glutamate degradation I showed significant sex effect (F(1,32) = 24.97, p < 0.0001, η2 = 0.4385) and was significantly higher in females than in males, irrespective of the groups (Additional file 1: Fig. S3C).
Fig. 10
Sex differences in the gut–brain and gut–metabolic modules before and after SPS. A Heat map of the shared gut–brain module between males and females before SPS exposure, B heat map of the shared gut–brain module between males and females following SPS exposure, C heat map of the shared gut–metabolic module between males and females before SPS exposure, D heat map of the shared gut–metabolic module between males and females following SPS exposure. Due to technical difficulties stool samples were not collected from every single animal. Each symbol represents the value for an individual animal. Before SPS (blue bars—males control n = 6/7, SPS-R n = 5/6, SPS-S n = 5; pink bars—females control n = 8/9, SPS-R n = 6/7, SPS-S n = 5/6), following SPS (blue bars—males control n = 6/7, SPS-R n = 6, SPS-S n = 4; pink bars—females control n = 8, SPS-R n = 8, SPS-S n = 5/6). The values of acetyl-CoA to crotonyl-CoA were multiplied by 1000 to facilitate their integration into the heat map alongside the other data points
Following SPS exposure. Only 2 gut–brain modules were altered (Fig. 10B). Gamma-hydroxybutyric acid (GHB) degradation showed significant sex effect (F(1,32) = 27.42, p < 0.0001, η2 = 0.4604), and it was more significant in control and SPS-R females than in males of the same group (Additional file 1: Fig. S3D). On the other hand, glutamate degradation I showed significant group (F(2,32) = 3.693, p = 0.0361, η2 = 0.1871) and sex (F(1,32) = 37.09, p < 0.0001, η2 = 0.5363) effects. The module was higher in SPS-S males compared to SPS-R and unstressed control males. However, compared to females, glutamate degradation was significantly lower in males than in females (Additional file 1: Fig. S3E).
Gut–metabolic module before and after SPS exposureBefore exposure to SPS. Seven GMM pathways were shared between the sexes (Fig. 10C). Methanogenesis from CO2 showed significant sex effect (F(1,32) = 13.35, p = 0.0009, η2 = 0.2931) and interaction between sex and group (F(2,32) = 3.459, p = 0.0437, η2 = 0.1785). While no sex-specific group differences were observed, unstressed controls and SPS-S females had significantly lower levels of methanogenesis than did their respective male rats (Additional file 1: Fig. S4A). Similarly, hydrogen metabolism showed significant sex effect (F(1,32) = 9.239, p = 0.0047, η2 = 0.2245) with significant interaction between sex and group (F(2,32) = 5.061, p = 0.0123, η2 = 0.2390). Hydrogen metabolism was lower in SPS-S males than in controls and SPS-R males; but was higher in SPS-R males than in SPS-R females (Additional file 1: Fig. S4B). Pentose phosphate pathway also showed sex (F(1,32) = 21.05, p < 0.0001, η2 = 0.397), and group (F(2,32) = 3.335, p = 0.0484, η2 = 0.173) effects, with an interaction between the two factors (F(2,32) = 7.459, p = 0.0022, η2 = 0.3182). It was lower in SPS-R males but higher in SPS-R females compared to their respective SPS-R and control groups. Moreover, the pathway was higher in SPS-R females than in SPS-R males (Additional file 1: Fig. S4C). Additionally Entner–Doudoroff pathway (F(1,32) = 22.97, p < 0.0001, η2 = 0.4185) (Additional file 1: Fig. S4D), acetyl-CoA to crotonyl-CoA (F(1,32) = 67.55, p < 0.0001, η2 = 0.6783) (Additional file 1: Fig. S4E), and glyoxylate/succinate (F(1,32) = 50.57, p < 0.0001, η2 = 0.6127) (Additional file 1: Fig. S4F) showed only sex effects and were all higher in females than in males, with acetyl-CoA to crotonyl-CoA showing the most marked differences between males and females. Finally, Pectin degradation II showed significant sex effect (F(1,32) = 11.78, p = 0.0017, η2 = 0.2680) and interaction between sex and group (F(2,32) = 4.371, p = 0.0210, η2 = 0.2144), and was significantly lower in females than in males (Additional file 1: Fig. S4G).
Following exposure to SPS. Five modules were shared between the sexes (Fig. 10D). Methanogenesis from CO2 (F(1,32) = 7.403, p = 0.0104, η2 = 0.1885) (Additional file 1: Fig. S5A), pectin degradation II (F(1,32) = 39.20, p < 0.0001, η2 = 0.5503) (Additional file 1: Fig. S5B), Glyoxylate/Succinate (F(1,32) = 30.54, p < 0.0001, η2 = 0.4886) (Additional file 1: Fig. S5C), and acetyl-CoA to crotonyl-CoA (F(1,32) = 60.65, p < 0.0001, η2 = 0.6544) (Additional file 1: Fig. S5D) were similar to before SPS, with the acetyl-CoA to crotonyl-CoA module remaining strikingly higher in females compared to males. However, lactaldehyde degradation (F(1,32) = 6.940, p = 0.0129, η2 = 0.1785) (Additional file 1: Fig. S5E) showed sex effect and alterations only after SPS and was higher in SPS-R females than in SPS-R males.
Sex differences in cecal bacterial metabolites (short-chain fatty acids) following SPSThe cecal weight at the time of dissection (day 39) was similar among the groups and between sexes (Additional file 1: Fig. S6A) however, the cecal SCFA levels were different. Cecal acetate levels showed significant group (F(2,23) = 4.977, p = 0.016, η2 = 0.3021) and sex (F(1,23) = 5.476, p = 0.0283, η2 = 0.1922) effects. In male rats, the levels of acetate were significantly lower in the SPS-S subgroup compared to SPS-R and unstressed controls; however, no differences were observed among females. When the levels were compared between males and females, SPS-R females showed significantly lower levels than did SPS-R males (Fig. 11A). The levels of propionate did not differ among the groups in males and females, yet significant sex effect (F(1,23) = 8.936, p = 0.0066, η2 = 0.2808) was observed, with SPS-S females having significantly higher levels of propionate than SPS-S males (Fig. 11B). Although a sex effect (F(1,23) = 5.904, p = 0.0233, η2 = 0.2042) was observed in the levels of cecal butyrate, multiple comparison tests did not show any differences within or between the sexes (Additional file 1: Fig. S6B).
Fig. 11
留言 (0)