
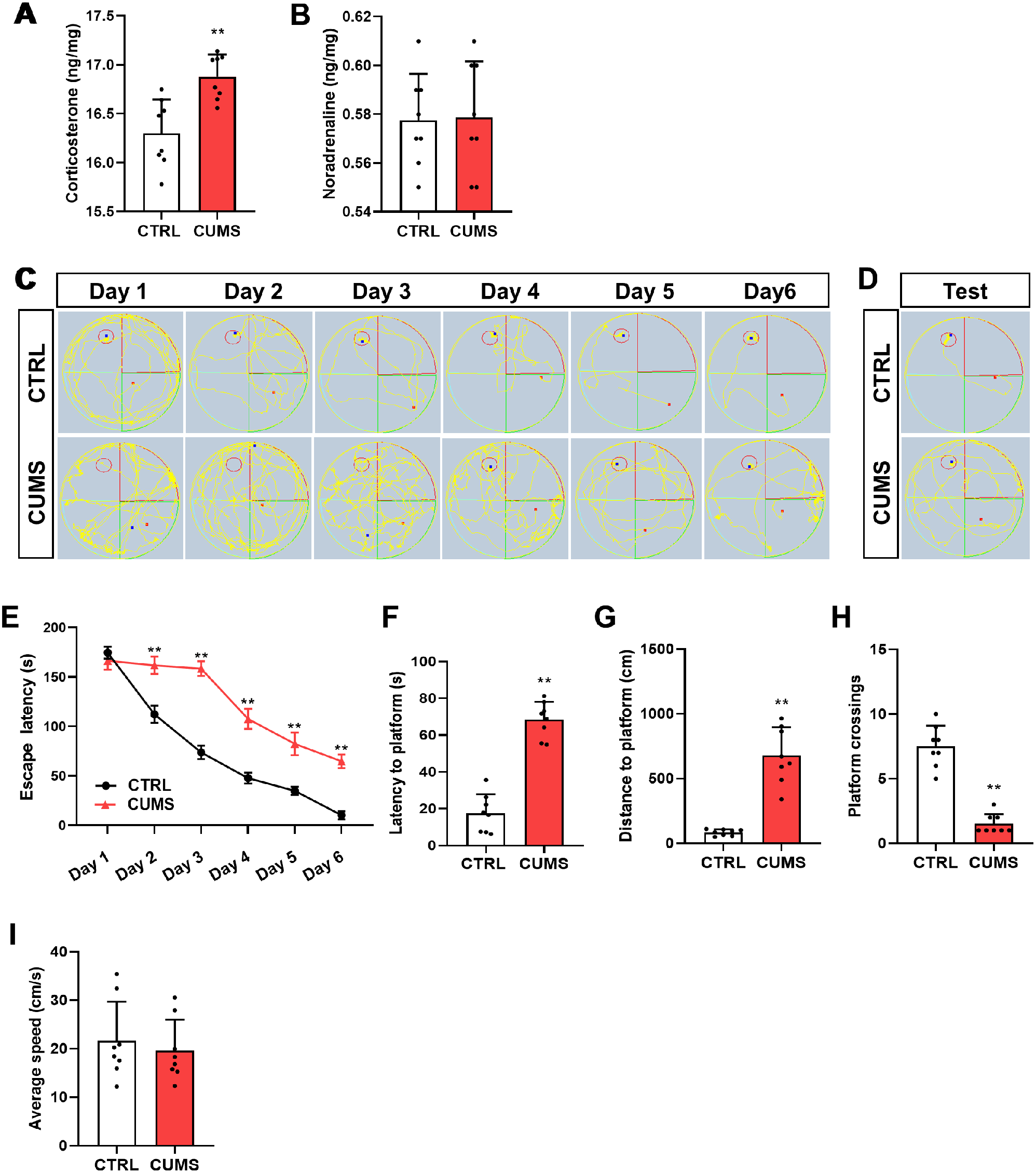
記住我





Abnormal seminiferous tubules, characterized by a SCO phenotype, were observed in both Pramex1 sKO or Pramel1 sKO male mice [23, 24]. To determine the frequency of SCO seminiferous tubules, we conducted the whole-mount IFS on seminiferous tubules of neonatal mice with the germ cell-specific marker TRA98. The SCO tubules were identified by the absence of germ cells, as evidenced by the lack of TRA98-positive cells in the whole-mount IFS (Fig. 1A). The occurrence of SCO tubules at P3 and P7 neonatal testis was 1.57 ± 0.08% and 7.78 ± 0.18% in Pramex1 sKO mice, 7.70 ± 0.52% and 8.42 ± 0.08% in Pramel1 sKO mice, respectively (Fig. 1A, B). In contrast, WT mice exhibited only ~ 1% (Fig. 1A, B). The rate of SCO tubules was significantly different at P3 (P < 0.05) but not at P7 (P > 0.05) between Pramex1 and Pramel1 sKO mice. These findings suggested that both Pramex1 and Pramel1 affect spermatogenesis in neonatal testes albeit potentially impacting germ cells differentially at distinct time points.
Fig. 1
SCO seminiferous tubules observed in the testis of mutant mice. A Representative images of whole-mount IFS with TRA98 (red) on seminiferous tubules of WT, Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice at P3 and P7. TRA98 was used to label germ cells. The SCO regions lack germ cells and are represented by seminiferous tubules without any TRA98 + cells. White dashed line: SCO regions. Nuclei counterstained with DAPI (blue). Bar = 100 µm. B Percentage of SCO segments (%) based on the whole-mount IFS and IFS from P3-35. Significance was assessed among the four groups (WT, Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice) for each time point (P3, P7, P21 and P35). Data were expressed as mean ± SEM. Values that do not share a common superscript (a–c) were different significantly (P < 0.05). C Representative images of IFS with SOX9 (red) counterstained with DAPI (blue) on testis cross-sections of the WT, sKO and dKO (at P21, P28 and P35) mice. SOX9 was a Sertoli cell specific-marker. Dense Sertoli cells were observed along the basement membrane of the SCO tubules where no germ cells were identified (outlined with a white dotted line). The selected regions from the P28 dKO image are enlarged in D. D Enlarged images from the P28 dKO section in C. Testis cross-sections from P28 dKO were stained with germ cell-specific markers TRA98 and DDX4, as well as spermatocyte and spermatid-specific marker MSY2. No signal was detected in SCO tubules, whereas strong signals were observed for either TRA98 and DDX4 (green in the left image) or TRA98, DDX4, and MSY2 markers (yellow in the right image) in normal tubules adjacent to SCO tubules. Bar = 100 µm
SCO tubules in Pramex1 sKO persisted until P35, in line with previous findings in Pramel1 sKO testes [24] (Fig. 1B–D). The SCO tubules were clustered rather than randomly distributed across the entire cross-section. By P21 and P35, the frequency of SCO tubules remained consistent in both Pramex1 sKO (7.42 ± 0.39% and 6.99 ± 0.94%) and Pramel1 sKO (6.60 ± 0.63% and 7.22 ± 0.81%) mice, respectively (Fig. 1B, C) (P < 0.05). The SCO tubules in the testis cross-section were confirmed by SOX9 for Sertoli cells and TRA98, DDX4 and MSY2 for germ cells (Fig. 1C, D). These findings indicated that the single gene ablation of Pramex1 or Pramel1 results in a similar occurrence of abnormal seminiferous tubules during spermatogenesis.
The severe defects in dKO males suggested a synergistic enhancement between the Pramex1 and Pramel1 genesTo investigate potential genetic interactions between Pramex1 and Pramel1 genes during gametogenesis, we developed a Pramex1/Pramel1 dKO mouse model (Table 1). We bred the Pramex1 sKO and Pramel1 sKO homozygous to produce the heterozygous F1 dKO mice. Theoretically, the F2 progeny from the F1 mating had a 6.25% chance of being dKO females and an equivalent likelihood (6.25%) of being dKO males. However, no dKO females were observed in F2, and only 1.35% of pups had the dKO male genotype (P = 4.52 × 10–8) (Table 3). To enhance the production of dKO mice, heterozygous females (Pramex1−/−; Pramel1−/+) were mated with dKO males, which would theoretically yield pups with 25% probability of being either dKO males or dKO females in the F3 generation. Surprisingly, the observed frequency of dKO males and females in F3 generation was only 6.49% and 5.85%, respectively, significantly lower than theoretical prediction (P = 1.4 × 10–14) (Table 4).
Table 3 The number (and percentage) of pups produced in the F2 generationTable 4 The number (and percentage) of pups produced in the F3 generationIn addition to PCR-genotyping, Western blotting (WB) was applied to exam whether PRAMEX1 or PRAMEL1 protein was expressed in the sKOs and dKO testes at P7. The PRAMEL1- (or PRAMEX1-) specific antibody detected a ~ 57 kDa (or ~ 56 kDa) band (expected molecular weight for the mouse PRAMEL1 or PRAMEX1) in WT and Pramex1 (or Pramel1) sKO mice, but not in Pramel1 (or Pramex1) sKO and Pramex1/Pramel1 dKO testis (Fig. 2A), confirming the deletion of the protein in the corresponding mutant mice. Interestingly, the PRAMEX1 protein expression was 1.71-fold higher in the Pramel1 sKO testis when compared to WT (P < 0.01) (Fig. 2B). Similarly, the PRAMEL1 protein was 1.56-fold higher in the Pramex1 sKO testis in comparison to WT (P < 0.01) (Fig. 2B). These results suggested that the ablation of either Pramex1 or Pramel1 may induce an upregulated expression of the other gene in the testis. The gene deletion in three mutant mice at P7 and the upregulation of the other gene in the two sKO mice were confirmed at the gene transcriptional level by qRT-PCR with Pramex1- and Pramel1-specific primers (Fig. 2C, D). This compensatory expression has the potential to alleviate the impact of individual mutations, implying that more pronounced defects could manifest when both genes are simultaneously deleted.
Fig. 2
Deletion of PRAMEL1 and PRAMEX1 in the Pramex1/Pramel1 dKO mice. A Western blot (WB) was used to detect the PRAMEL1 and PRAMEX1 expression in the WT, Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice at P7. B Relative expressions of PRAMEL1 and PRAMEX1 in the Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice to WT (WT was set as 1) were detected in WB. C Cycle threshold (CT) value of qRT-PCR for Pramel1, Pramex1 and Actb genes was compared between the WT, Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice. CT values above 37 indicate minimal amounts or none of target nucleic acid. D Relative expression of Pramel1 and Pramex1 in the Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice to WT (WT was set as 1) were detected by qRT-PCR. PRAMEL1 was not deleted in Pramel1 sKO and Pramex1/Pramel1 dKO mice and ablation of PRAMEX1 in Pramex1 sKO and Pramex1/Pramel1 dKO mice. In addition, higher expression of PRAMEL1 and PRAMEX1 was observed in Pramex1 sKO and Pramel1 sKO, respectively. Significance was assessed among the four groups (WT, Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice) for each antibody (anti-PRAMEL1 and anti-PRAMEX1). Data were expressed as mean ± SEM. Values that do not share a common superscript (a–c) were found to differ significantly (P < 0.05)
To determine the functional interaction of the Pramex1 and Pramel1 genes, we characterized the phenotypes of the dKO mice. As shown in Fig. 1, at P3 and P7, compared to both Pramex1 (1.57 ± 0.08%; 7.78 ± 0.18%) and Pramel1 (8.42 ± 0.08%; 7.70 ± 0.51%) sKO mice, the dKO testis showed a higher frequency of SCO tubules (P < 0.01), with 11.97 ± 0.18% and 18.29 ± 0.78% respectively. The elevated SCO tubule frequency in dKO mice at P7 compared to P3 coincided with the period of SCO region formation in Pramex1 sKO mice. In neonatal testis at P3 and P7, the SCO frequency in male dKO mice was 18–20% higher than the combined frequency in Pramex1 sKO and Pramel1 sKO males, suggesting synergistic enhanced defects in the dKO mice compared to sKO mice. Consistent with the SCO frequency in neonatal testis, the frequency of SCO tubules in the testes of dKO mice remained consistently high at approximately 18–20% during the first round of spermatogenic process from P21 to P35 (Fig. 1B–D).
Apart from the SCO tubules, the dKO mice displayed a greater quantity of apoptotic cells compared to the numbers observed in the sKO mice for Pramex1 and Pramel1. At P7, dKO mice showed a higher number of TUNEL + cells per tubule (1.18 ± 0.06) than Pramex1 sKO (0.69 ± 0.01) and Pramel1 sKO (0.51 ± 0.15) mice (P < 0.01) (Fig. 3A, B). Similarly, the percentage of tubules with TUNEL + cells in the dKO (39.49 ± 1.34) at P7 was close to the sum of Pramex1 (18.40 ± 1.03) and Pramel1 sKO (24.66 ± 5.31) mice (Fig. 3C). Like Pramex1 sKO and WT mice, dKO mice displayed a peak of germ cell apoptosis at P14 (Fig. 3A, B). The apoptotic results, including the number of TUNEL + cells per tubule (3.36 ± 0.12) and the percentage of tubules with TUNEL + cells (65.7 ± 3.70), in the dKO mice at P14 were similar to those in Pramex1 sKO mice (P > 0.05) but higher than those in Pramel1 sKO and WT mice (P < 0.01) (Fig. 3C). A 2.1-fold increase in TUNEL + cells and a 30% higher percentage of tubules with TUNEL + cells were observed in the dKO compared to WT mice at P14. At P21, apoptotic cell numbers were comparable to those at P14 in all four groups (Fig. 3B, C). Similarly, at P14, both dKO and Pramex1 sKO males exhibited higher apoptotic cell counts compared to WT and Pramel1 sKO mice at P21. As the mice progressed to P28 and P35, there was a general decrease in apoptotic cell numbers across all groups. In conclusion, the higher incidence of abnormalities, such as SCO tubules and apoptotic cells, observed in the dKO mice was anticipated to lead to more pronounced phenotypic defects.
Fig. 3
Germ cell apoptosis was increased in the sKO and dKO testis. Testis was evaluated at P7, P14, P21, P28 and P35 and apoptosis was assessed by TUNEL assay in the WT, Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice. A Representative TUNEL (green) staining of testis sections in the WT, Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice at P7 to P15. More apoptotic cells occur in the lumen of seminiferous tubules in the dKO than sKO and WT mice. Bar = 20 µm. B Number of TUNEL-positive (TUNEL+) cells in the mouse seminiferous tubules for the four groups of mice. C Percentage of seminiferous tubules with apoptotic cells (TUNEL+). Apoptotic cells were significantly increased in the dKO mice than that sum of Pramex1 sKO and Pramel1 sKO. Data were expressed as mean ± SEM
To further validate the compensatory capacity of the two genes and comprehend the impact of a mutation in one gene on the other, we assessed various phenotypes in the dKO males, including testis weight, testis index, sperm counts, and litter size, and compared the observed value with a referenced value based on a gene additive genetic model (see Methods). The testis weight and testis index of dKO mice were significantly lower than sKOs and WT (Fig. 4A, B) (P < 0.05). Furthermore, we assessed caput and cauda sperm production from the first round of spermatogenesis at P41, and from subsequent rounds of spermatogenesis in the cauda at P60, P120, and P365. The dKO males displayed a reduction in sperm count ranging from 12 to 58% compared to WT, which were 13–29% lower than referenced value at all ages studied (P < 0.05) (Fig. 4C). The decrease in sperm count in dKO males at P120 was confirmed by CASA analysis, though no significant difference were found in sperm motility between dKO and WT mice (Table 5). Additionally, all sKO, dKO and WT males, from the age of P41, were paired with adult WT females to evaluate their fecundity. The average number of offspring per litter, produced by Pramex1 sKO, Pramel1 sKO, dKO and WT males (n = 6 ~ 7), was 6.50 ± 0.41, 6.39 ± 0.36, 4.08 ± 0.30, and 7.76 ± 0.38, respectively (Fig. 4D). The number of offspring produced by dKO males was reduced by 47.41% compared to WT males (P < 0.01) (Fig. 4D). These observations were significantly lower than the referenced anticipated value (Fig. 4C-D) suggesting a synergistic enhancement between the two genes.
Fig. 4
Severe reproductive defects observed in the Pramex1/Pramel1 dKO mice. A Testis weight of WT, Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice obtained during the time-course study from P7 to P365. B Testis index (= testicular weight (g) / body weight (g) × 100) was documented in WT, Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice from P7 to P365 (n = 3 ~ 5). C Sperm count in epididymis of WT, Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice at P41 (caput and cauda), P60 (cauda), P120 (cauda) and P365 (cauda) of age (n = 3 ~ 5). The referenced sperm count of the dKO was drawn as red solid line in the figure. The formula for the referenced anticipated value of dKO, which calculate effect of two genes deletion without gene interaction in the additive model, was as follows: (\(\mathrm=Pramel1 }\times \frac}}}\)[44,45,46] (n = 3 ~ 5). D Litter size of mating tests of male fecundity. The experimental male mice (P60) were mated with adult WT female. The referenced litter size of the dKO was drawn as red solid line. About 6 ~ 7 male mice were tested and about 30 litters were counted for each group. Significance was assessed among the four groups (WT, Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice) for C and D. Data were expressed as mean ± SEM. Values that do not share a common superscript (a–d) were found to differ significantly (P < 0.05)
Table 5 Cauda sperm count and motility analysis using CASA at P120Reduction of undifferentiated spermatogonia in Pramex1/Pramel1 dKO miceTo understand the cellular mechanism behind the elevated rate of SCO tubules and reduced sperm count in the dKO mice, we quantified undifferentiated spermatogonia (ID4 + cells/mm2) at two developmental stages, juvenile (P35) and mature (P120) (Fig. 5A). At P35, both Pramel1 sKO (21.86 ± 0.48/mm2) and dKO (19.01 ± 0.60/mm2) mice had significantly fewer undifferentiated spermatogonia compared to the WT mice (23.71 ± 0.32/mm2) (P < 0.01) (Fig. 5B), whereas difference between the Pramex1 sKO (22.91 ± 0.47/mm2) and WT mice was not statistically significant (P > 0.05). While the Pramel1 sKO mice exhibited an 8% reduction in undifferentiated spermatogonia compared to the WT mice (P < 0.01), the dKO mice displayed a significantly more pronounced decrease (20%) (P < 0.01), which was 10% lower than the referenced value (Fig. 5B) (P < 0.01). With the progression of aging in the mice, we noted a progressively scattered arrangement of undifferentiated spermatogonia, attributed to the elongation of seminiferous tubules. This phenomenon led to a reduction in the quantity of ID4 + cells within a given unit size at P120 compared to P35 (Fig. 5A, B). At P120, the dKO mice (7.31 ± 0.14/mm2) as well as both the Pramex1 sKO (9.93 ± 0.16/mm2) and Pramel1 sKO (9.19 ± 0.09/mm2) mice had significantly fewer ID4 + cells compared to the WT mice (10.98 ± 0.21/mm2) (P < 0.01) (Fig. 5A, B). The dKO mice exhibited a 33.41% reduction in ID4 + cells than that of the WT mice, while Pramex1 sKO and Pramel1 sKO mice showed reductions of 9.61% and 16.35%, respectively (P < 0.01) (Fig. 5A, B). The referenced undifferentiated spermatogonia number for the dKO was 8.30 ± 0.08/mm2, which was 12% more than what was observed in dKO mice (P < 0.01) (Fig. 5B). These results indicated that the simultaneous ablation of both genes leads to a greater reduction in the ability of male mice to maintain the undifferentiated spermatogonia compared to the combined effects of ablating either one of the genes, confirming the synergistic enhancement between the two genes at the cellular level.
Fig. 5
Reduced number of undifferentiated spermatogonia in the Pramex1/Pramel1 dKO mice. A Representative images of IFS with ID4 (red) in testis cross-sections for WT, Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice at P35 and P120 (n = 3). Bar = 20 µm. B Number of ID4 + spermatogonia/mm2 based on IFS result for the four groups at P35 and P120. The referenced number of ID4 + undifferentiated spermatogonia in the dKO was drawn as red solid line. The IFS experiments in A-B were replicated three times. Significance was assessed among the four samples (WT, Pramex1 sKO, Pramel1 sKO, and dKO mice) at two ages (P35 or P120). Data were expressed as mean ± SEM. Values that do not share a common superscript (a–d) were found to differ significantly (P < 0.05)
Synergistic repression of RA/RAR signaling by Pramex1 and Pramel1 during spermatogenesisOur previous studies indicated that both Pramex1 and Pramel1 function in spermatogenesis by repressing the RA signaling [23, 24]. To understand how the synergistic interactions of Pramex1 and Pramel1 affect RA signaling, we administered trans-RA and its inhibitor (WIN18,466) in WT, Pramex1 sKO, Pramel1 sKO, and dKO mice at P2. We then evaluated the frequency of SCO tubules at P7 using whole-mount IFS with the TRA98 antibody (Fig. 6A). After RA treatment, the frequency of SCO tubules in WT, Pramex1 sKO, Pramel1 sKO, and dKO testis increased by 14%, 47%, 68%, and 73%, respectively (Fig. 6B), compared to the non-RA treatment group (Fig. 1B). This result indicated that RA treatment not only exacerbated the defect in the sKO and dKO mice but also affected the WT mice. Although the frequency of SCO tubules in either Pramex1 sKO (11.43 ± 0.31%) or Pramel1 sKO (12.95 ± 0.84%) was elevated after the RA treatment when compared to WT (1.91 ± 0.27%) (P < 0.01), it was significantly lower in the dKO mice (31.65 ± 0.82%) (P < 0.01) (Fig. 6A, B).
Fig. 6
The synergistic enhancement between Pramex1 and Pramel1 was through retinoic acid (RA) signaling pathway. A Representative images of whole-mount IFS with TRA98 (red) on seminiferous tubules of WT, Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice at P7, these animals were treated with RA or WIN18,446 (RA inhibitor) at P2. TRA98 was used to label germ cells. The SCO regions, labeled with white dashed line, lacked germ cells which were seminiferous tubules without any TRA98 + cells. Nuclei counterstained with DAPI (blue). Bar = 100 µm. B SCO segments along the length of seminiferous tubules (%) based on the whole-mount IFS for the RA/WIN18,446 treatment in the four groups (n = 3). Significance was assessed among the four groups (WT, Pramex1 sKO, Pramel1 sKO, and Pramex1/Pramel1 dKO mice) for each treatment (RA or RA inhibitor). C Co-immunoprecipitation (co-IP) analyses of PRAMEX1 and RARα in P7 WT, Pramex1 sKO and Pramel1 sKO testis tissue. YBX2 served as a control. D Based on the band intensity in the WB for WT testis tissue, the protein expression level relative to input WT testis (set as 1) was measured (n = 3). Significance was assessed among the three samples (input, IP/w PRAMEX1, and IP/w RARα) for each antibody (PRAMEX1, RARα and YBX2). E Based on the band intensity in the WB, the protein expression level relative to input Pramel1 sKO testis (set as 1) was measured (n = 3). Significance was assessed among the three samples (input, IP/w PRAMEX1, and IP/w RARα) for each antibody (PRAMEX1, RARα and YBX2). Data were expressed as mean ± SEM. Values that do not share a common superscript (a–d) were found to differ significantly (P < 0.05)
Upon inhibitor treatment, the frequency of SCO tubules in WT (1.51 ± 0.70%), Pramex1 sKO (1.62 ± 0.08%), and Pramel1 sKO mice (1.86 ± 0.86%) decreased to the baseline, suggesting that the SCO phenotype was recovered in the sKO mice. In contrast, SCO tubules were still present in the dKO mice, although in a much lower frequency (7.20 ± 0.43%) (Fig. 6A, B), implying a partial recovery in the dKO mice. The results indicated that the responsiveness of exogenous RA was altered in the sKO and dKO mice, and the dosage of RA inhibitor (WIN18,466) used in this study was adequate to offset the impact of single gene deletion, but not adequate to the double gene deletion. Thus, this data suggested a synergistic enhancement of RA signaling in the dKO mice, which required a higher dosage of inhibitor to counteract.
To further elucidate the role of PRAMEX1 within the RAR signaling pathway, we carried out a co-immunoprecipitation (co-IP) experiment to verify the interaction between PRAMEX1 and RARα. Relative to the expression in the WT testis input sample, the presence of PRAMEX1 in the RARα co-IP products exhibited a 1.3-fold increase, suggesting an enrichment of PRAMEX1 within the RARα protein complex (Fig. 6C, D). Moreover, the expression of RARα also displayed a 1.5-fold stronger band in the PRAMEX1 precipitation in WT testis compared to input sample (Fig. 6C, D), whereas the negative control (YBX2) was only detected in the input sample but not in either RAR or PRAMEX1 precipitations (Fig. 6C, D). These results suggested a direct interaction between PRAMEX1 and RARα, similar to the interaction of PRAMEL1 with RARα (as demonstrated in the co-IP data in [24]). In line with Fig. 2, in Pramex1 sKO mice, no PRAMEX1 was detected in the testis or precipitation samples (Fig. 6C). Conversely, Pramel1 sKO mice displayed a 3.32-fold increase of PRAMEX1 in the RARα co-IP products and 2.48-fold increase of RARα in PRAMEX1 precipitation relative to testis sample (Fig. 6C, E). These results suggest that in the absence of Pramel1, a compensatory mechanism partially restores the interaction with RARα by upregulating the Pramex1 gene. Our findings regarding RA and RAR signaling imply that this restoration occurs as the upregulated gene assumes the role of the deleted gene in repressing RAR signaling.
Compensatory gene effects were observed in Pramex1 and Pramel1 sKO females but absent in dKO femalesThe fecundity of Pramex1 and Pramel1 sKO female mice was examined by a female mating test, where mature Pramex1 and Pramel1 sKO female mice (2-months old, n = 7) were bred with mature WT male mice continually for 6 months. The WT female mice (2-months old, n = 7) served as controls. The litter sizes of the female Pramex1 and Pramel1 sKO mice (7.84 ± 0.28 and 7.52 ± 0.75) were not significantly different from the WT female (7.76 ± 0.38) (P = 0.43 and 0.33) (Fig. 7A), suggesting that the fecundity of both Pramex1 and Pramel1 sKO females was normal. Apparently, the ablation of either Pramex1 or Pramel1 did not have a visible effect on female fecundity. These results raised a question about whether the Pramex1 and/or Pramel1 are male-specific, as a previous report suggested both Pramex1 and Pramel1 predominantly expressed in the testis, but not in the ovary [14, 38]. We conducted IFS with the PRAMEX1- and PRAMEL1-specific antibodies on mature WT ovarian sections and observed a clear enrichment of both PRAMEX1 and PRAMEL1 proteins in the cytoplasm of oocytes in secondary and antral follicle (Fig. 7B). If PRAMEX1 and PRAMEL1 are expressed in oocytes, why did the Pramex1 or Pramel1 sKO females not show any reproductive phenotypes? One feasible explanation could be that the Pramex1 and Pramel1 genes possess compensatory functions with each other in female mice. To test the hypothesis, a mating test was performed for the Pramex1/Pramel1 dKO females (bred with mature WT males for 6 months, n = 6). The average number of offspring in the dKO females was 3.31 ± 0.29, which was significantly lower than in Pramex1 and Pramel1 sKO (7.84 ± 0.28 and 7.52 ± 0.75) and WT females (7.76 ± 0.38) (P < 0.01) (Fig. 7A). The litter size of dKO females was 57% smaller than that of WT mice, paralleling the reduced fecundity observed in dKO male mice (Fig. 4D). These results indicated that the ablation of both Pramex1 and Pramel1 significantly affects the female reproductive capacity, confirming their compensatory role in female reproduction.
Fig. 7
Pramex1 and Pramel1 exhibit compensatory effects on the fecundity of sKO female mice. A Litter size for female mating tests (n = 7 ~ 8). Data were expressed as mean ± SEM. Significance was assessed among the four mattings (WT♀ x WT♂, Pramex1 sKO♀ x WT♂, Pramel1 sKO♀ x WT♂, female dKO♀ x WT♂ mice). B IFS for PRAMEX1 and PRAMEL1 in ovary cross-section. Bar = 50 µm. C The litter size of different mattings suggests that the fecundity of both male and female Pramex1/Pramel1 dKO mice was decreased (n = 7 ~ 8). Significance was assessed among the four mattings (WT♀ x WT♂, WT♀ x dKO♂, Pramel1 sKO♀ x dKO♂, dKO♀ x dKO♂ mice) which producing 29, 30, 31, 40 litters from 7, 7, 8, 11 breeding pairs, respectively. Data were expressed as mean ± SEM. Values that do not share a common superscript (a–c) were found to differ significantly (P < 0.05)
During the generation of dKO mice, a significantly lower-than-expected rate of dKO females was produced in the F2 (P = 4.52 × 10–8) and F3 generations (P = 1.44 × 10–14), respectively (Table 1). To explore the underlying causes, we conducted a thorough analysis of gamete genotypes produced by F1 and F2 animals and their male-to-female sex ratio among offspring. In F2, the sex ratio of the offspring was 1:1.04 as expected (Table 3). The F1 heterozygous males and females were expected to produce four different genotypes of sperm and eggs, yielding 16 different genotypes of offspring in F2 generation, with an anticipated distribution of 6.25% for each genotype under a normal breeding condition (Table 3). However, the observed distribution of genotypes differed significantly from the prediction (P = 4.52 × 10–8) (Table 1). Among the 296 pups analyzed, none of them were dKO females (Pramex1−/−, Pramel1−/−), while only 4 (1.35%) were dKO males (Pramex1−/Y, Pramel1−/−), which were significantly lower than the expected 6.25%. In contrast, 30 pups (10.14%) were WT males (Pramex1+/Y, Pramel1+/+), which were significantly higher than the expected rate (Table 3). The total number of pups produced from "Pramex1−, Pramel1−" eggs and sperm were 37 (12.5%) and 50 (16.9%), respectively, which were significantly lower than the expected 74 pups (25%) (P = 5.96 × 10–6) (Table 3). These results suggested that loss of homozygous dKO embryos occurred during the heterozygous male and heterozygous female dKO mating, and the Y-bearing "Pramel1−, Y" sperm had a less effect on embryonic survive than the X-bearing "Pramex1−, Pramel1−" sperm.
A total of 154 pups were produced in the F3 generation by mating heterozygous dKO females (Pramex1−/+;Pramel1−/−) with homozygous dKO males (Pramex1−/Y;Pramel1−/−), which produced two genotypes of eggs "Pramex1−, Pramel1−" and "Pramex1+, Pramel1−" and two genotypes of sperm, the X-bearing "Pramex1−, Pramel1−" and Y-bearing "Pramel1−, Y" sperm (Table 4). Consistent with the observation in the F2 generation, the number of homozygous dKO female and male pups produced from the "Pramex1−, Pramel1−" eggs was 9 (5.9%) and 10 (6.5%), respectively, significantly lower than the expected 25% (or 38.5 pups) (P = 1.44 × 10–14) (Table 4). In contrast, the single gene deletion "Pramex1+, Pramel1−" eggs produced 59 female (38.3%) and 76 male (49.4%) pups, significantly higher than the expected 25%. Furthermore, the male-to-female sex ratio was skewed (1:0.79) in this mating (P = 0.1469) (Table 4).
We further analyzed the litter size data from our mating records, including mature WT, Pramel1 sKO, and dKO female mice bred with dKO males. As expected, the average litter size of WT♀ x dKO♂ (4.08 ± 0.30) was similar to Pramel1 sKO♀ x dKO♂ (3.94 ± 0.42) (P > 0.05) (Fig. 7C). The dKO♀ x dKO♂ mating produced an average of 2.03 ± 0.18 pups/litter, which was approximately 50% smaller than the average litter size of sKO♀ x dKO♂ (P < 0.01) (Fig. 7C). It is worth noting that the dKO♀ x dKO♂ mating produced a total of 81 pups (42♂ and 39♀) with a male-to-female sex ratio of 1:0.93 (P > 0.05).
Taken together, our data suggested that the function of Pramel1 was compensated by the Pramex1 gene in the Pramel1 sKO females. The function of "Pramex1−, Pramel1−" oocytes was affected more than X-bearing sperm "Pramex1−, Pramel1−". Therefore, embryos produced from this type of eggs had a less survival rate compared to those produced from "Pramex1−, Pramel1−" sperm. However, it was unclear if the lower-than-expected homozygous dKO pups found in this study was due to embryonic lethality (and loss) or germ cell loss during gametogenesis.
Follicle quantity and early embryonic count decrease in Pramex1/Pramel1 dKO femalesTo further explore the effects of the concurrent absence of PRAMEX1 and PRAMEL1 on oogenesis, we conducted a quantitative analysis of DDX4-positive primary follicles in dKO ovaries at P3. The Pramex1/Pramel1 dKO females (1038.47 ± 261.91/ovary) exhibited a notable 51% reduction in the number of follicles compared to WT mice (2126.57 ± 249.75/ovary) (P < 0.01) (Fig. 8A, B). This reduction aligns with the 57% decrease in litter size observed in female dKO mice when compared to WT females. The results indicated that the double deletion of the two genes led to a decrease in the number of follicles per ovary.
Fig. 8
The Pramex1/Pramel1 dKO females exhibit reduced number of follicles, which leads to embryo loss. A Representative images of IFS with DDX4 (green) in ovary cross-sections for WT and Pramex1/Pramel1 dKO mice at P3 (n = 3). Bar = 50 µm. B Total number of follicles per ovary. The whole ovary was sectioned, and the counting was performed every five sections, and the number follicles was multiplied by five to determine the total number of follicles per ovary. C–E Results from in vitro fertilization (IVF) experiment. The quality of oocyte with genotyping of "Pramex1+, Pramel1−" and "Pramex1−, Pramel1−" were detected through in-vitro fertilize with "Pramex1−, Pramel1−" or "Pramel1−,Y" spermatozoa. C Representative figures of embryos (2-cell and 8-cell) after in vitro fertilization for Pramel1 sKO and Pramex1/Pramel1 dKO oocyte and dKO spermatozoa. Bar = 100 µm. D Number of denuded oocytes and cumulus-oocyte complexes (COCs) in each animal for Pramel1 sKO and Pramex1/Pramel1 dKO (n = 3). E Rate of 2-cell and 8-cell embryos from Pramel1 sKO and Pramex1/Pramel1 dKO oocytes. The rate was calculated by dividing the number of two-cells or eight-cells embryos by the number of COCs. F Representative images illustrating the uteri at E6.5 in WT and dKO mice, aimed at enumerating implantation sites. Bar = 2 mm. G The number of implantation sites in WT and dKO female at E6.5. Data were expressed as mean ± SEM. ** indicates statistically significant difference (P < 0.01)
To evaluate the effect of the two genotypes of eggs "Pramex1−, Pramel1−” and "Pramex1+, Pramel1−" on fertilization and early embryo development, we conducted in vitro fertilization (IVF) with oocytes collected from dKO and Pramel1 sKO mice at P60. We assessed several parameters, including the number of cumulus enclosed and denuded oocytes, and the rate of embryos at two-cell and eight-cell stage. The total number of oocytes, including COCs and denuded oocytes, in the dKO group (22.33 ± 2.57/animal) was 45% lower than that in the sKO group (40.89 ± 2.43/animal) (P < 0.01). The proportion of oocytes contained in a COC was similar in both groups (dKO 23% and sKO 26%, P > 0.05) (Fig. 8C, D). This finding was consistent with our earlier results (Fig. 8A, B), demonstrating that dKO females exhibited a 51% decrease in follicle count within the ovaries. For IVF experiments, only oocytes within a COC were used. We found that the average number of dKO COC that were fertilized (2-cell) and progressed to the 8-cell stage were 1.22 ± 0.75 and 1.11 ± 0.70 while the number of 2-cell and 8-cell in sKO were 8.78 ± 0.87 and 8.33 ± 0.77 (Fig. 8C, E). In the dKO group, the rate (22.8 ± 10.54%) of 2-cells embryos was approximately 73% less than that in the sKO group (83.82 ± 10.35%) (P < 0.01) (Fig. 8E). Consistently, the rate of 8-cells embryos produced in the dKO group (20.58 ± 10%) was also significantly lower (74.2%) than that in the sKO mice (79.76 ± 11.18%) (P < 0.01) (Fig. 8E). In this study, we found that in vitro embryo productions rates from double-deletion male and female gametes were reduced at the two-cell stage, mirroring reduction in litter size observed in female dKO mating test.
In addition to evaluating early embryo development, we also examined the number of implantation sites in the uterus at E6.5 from a P60 female to determine the effect of double deletion. The uteri were obtained from WT and dKO females, while the sires of the embryos were WT and dKO males, respectively. The average number of implantation sites in the uteri of dKO female was 2.25 ± 0.74 which was significantly less (74%) than WT females (8.50 ± 1.00) (P < 0.01) (Fig. 8F–G). The number of implantation sites in the uteri of dKO females corresponds to the number of 2-cell and 8-cell embryos and the litter size in dKO mice (Fig. 8E and Fig.
留言 (0)