
記住我




Currently, data describing the effects of MSI2 on tumor initiation and progression have been extensively explored [19, 21]. However, no study has focused on the role of MSI2 in tumor iron death, particularly in CRC. To explore the ferroptosis effect of MSI2 on CRC, we first analyzed MSI2 expression through the TCGA CRC datasets via the GEPIA tool and found that the transcript level of MSI2 was higher in tumor tissue than in normal tissue (Fig. 1A). Consistently, compared with that in adjacent normal colon tissues, the mRNA expression of MSI2 was upregulated in our 50 pairs of clinical CRC specimens (Fig. 1B). Elevated protein levels of MSI2 were also found in CRC primary tumors compared to the normal tissues in the CPTAC datasets, which was further confirmed by immunofluorescence histological examination of our clinical CRC patient samples (Fig. 1C and D). Next, to clarify the association between MSI2 and ferroptosis in CRC, we collected 620 CRC patients from the TCGA database and 779 normal tissues from GTEx. And CRC patients were classified into high and low expression groups based on MSI2 transcript levels, and we observed that the expression of GPX4, SLC7A11, FANCD2, NFE2L2 and HSPA5 was upregulated in CRC patients (Fig. 1E). Among them, the transcriptional expression of ferroptosis inhibitor molecules was upregulated in MSI2-High patients, such as SLC7A11, NFE2L2, HSPA5, FANCD2, HELLS, etc. (Fig. 1F). And KEGG pathway enrichment also showed that the differentially expressed genes involved in the small cell lung cancer, prostate cancer, hepatocellular cancer and colorectal cancer pathways were significantly up-regulated, while genes involved in the oxidative phosphorylation, non-alcoholic fatty liver disease, fat digestion and absorption, chemical carcinogenesis-reactive oxygen species and neuro-degenerative disease pathways were significantly down-regulated (Fig. 1G). Notably, MSI2 expression was significantly associated with ferroptosis inhibitor molecules, which were upregulated in patients with high MSI2 expression; including BRD4, NFE2L2, STAT3, HSPA5, CREB1, SLC7A11, ATF4, FANCD2, JUN, CD44, HELLS, CISD1 and SCD (Fig. 1H). Furthermore, the positive correlations between MSI2 and ferroptosis inhibitor genes in TCGA CRC datasets were further identified, and the positively correlated genes included BRD4 (R = 0.24, p = 6.8e-10), NFE2L2 (R = 0.56, p = 1.48e-51), STAT3 (R = 0.42, p = 1.89e-27), HSPA5 (R = 0.50, p = 6.93e-41), CREB1 (R = 0.68, p = 5.72e-84), SLC7A11 (R = 0.58, p = 6.44e-56), ATF4 (R = 0.24, p = 1.47e-09), FANCD2 (R = 0.35, p = 6.15e-19), CD44 (R = 0.31, p = 2.22e-15), HELLS (R = 0.47, p = 1.17e-34), CISD1 (R = 0.23, p = 1.29e-08) and SCD (R = 0.22, p = 4.87e-08) (Fig. 1I). Similarly, the positive correlations between MSI2 and ferroptosis inhibitor genes were also confirmed in the GEPIA and TIMER CRC datasets (Fig. 1J and K). More importantly, our TUNEL staining and histological analysis revealed that clinical CRC samples with low MSI2 expression exhibited more tissue necrotic cell death and higher rates of TUNEL-positive staining than those with high MSI2 expression (Fig. 1L). Collectively, these findings suggest that MSI2 expression is positively correlated with the expression of ferroptosis inhibitor genes, which may control iron death in CRC.
Fig. 1
MSI2 expression is increased in CRC and positively correlated with the expression of ferroptosis-related inhibitory molecules. A The expression of MSI2 is upregulated in the CRC TCGA database from GEPIA (COAD, N = 349, T = 275; READ, N = 318, T = 92). B MSI2 mRNA expression was measured by qRT‒PCR in 50 paired clinical CRC specimens. C MSI2 protein expression was increased in colon primary tumors from CPTAC (N = 100, T = 97). D Representative IFC images of MSI2 expression in our clinical CRC specimens. Blue: DAPI, green: MSI2; Scale bars, 200 μm. E Heatmap of ferroptosis-related gene expression in MSI2-high (n = 310) and MSI2-low (n = 310) patients from TCGA CRC datasets and in normal tissues (n = 779) from GTEx. The different colors represent the expression trend of the indicated gene in different samples. F The volcano plot of the differentially expressed ferroptosis-related genes in MSI2-high (n = 310) and MSI2-low (n = 310) patients, “Adjusted p < 0.05 and Log2(Fold Change MSI2-high/MSI2-low) > 1.3 or < − 1.3” were defined as the threshold for DEGs. G The top up-regulated and down-regulated KEGG pathway enriched by the differentially expressed genes in MSI2-high (n = 310) and MSI2-low (n = 310) patients. H The mRNA expression distribution of ferroptosis-related inhibitory molecules in MSI2-high (n = 310) and MSI2-low (n = 310) patients. The abscissa represents differentially expressed ferroptosis-related inhibitory genes, and the ordinate represents the expression distribution of genes. I The positive correlations between MSI2 and ferroptosis-related inhibitory genes were analyzed by Spearman correlation analysis in the TCGA CRC datasets, n = 620. J Heatmap of positive correlations between MSI2 and ferroptosis-related inhibitory genes were analyzed by Spearman correlation analysis and normalized by GAPDH in the GEPIA COAD and READ database, n = 367. K The positive correlations between MSI2 and ferroptosis-related inhibitory genes were analyzed by Spearman correlation analysis in the TIMER COAD database, n = 457. The ferroptosis-related genes set (H-K) were obtained from the public available FerrDb databases (http://www.zhounan.org/ferrdb/current/). L Representative images of necrotic cell death and TUNEL staining in MSI2-high and MSI2-low clinical CRC adenocarcinoma tissues. Blue: DAPI, red: MSI2, green: TUNEL; H&E, Scale bars, 200 μm (left) and 50 μm (right); IFC, Scale bars, 200 μm and 100 μm. and statistical analysis of TUNEL-positive cell percentages from MSI2-high and MSI2-low adenocarcinoma tissues. These results are presented as the mean ± SD values; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001; B paired 2-tailed Student’s t test, A, C, H Wilcox test, E Kruskal‒Wallis test, I-K Spearman correlation analysis and L unpaired 2-tailed Student’s t test
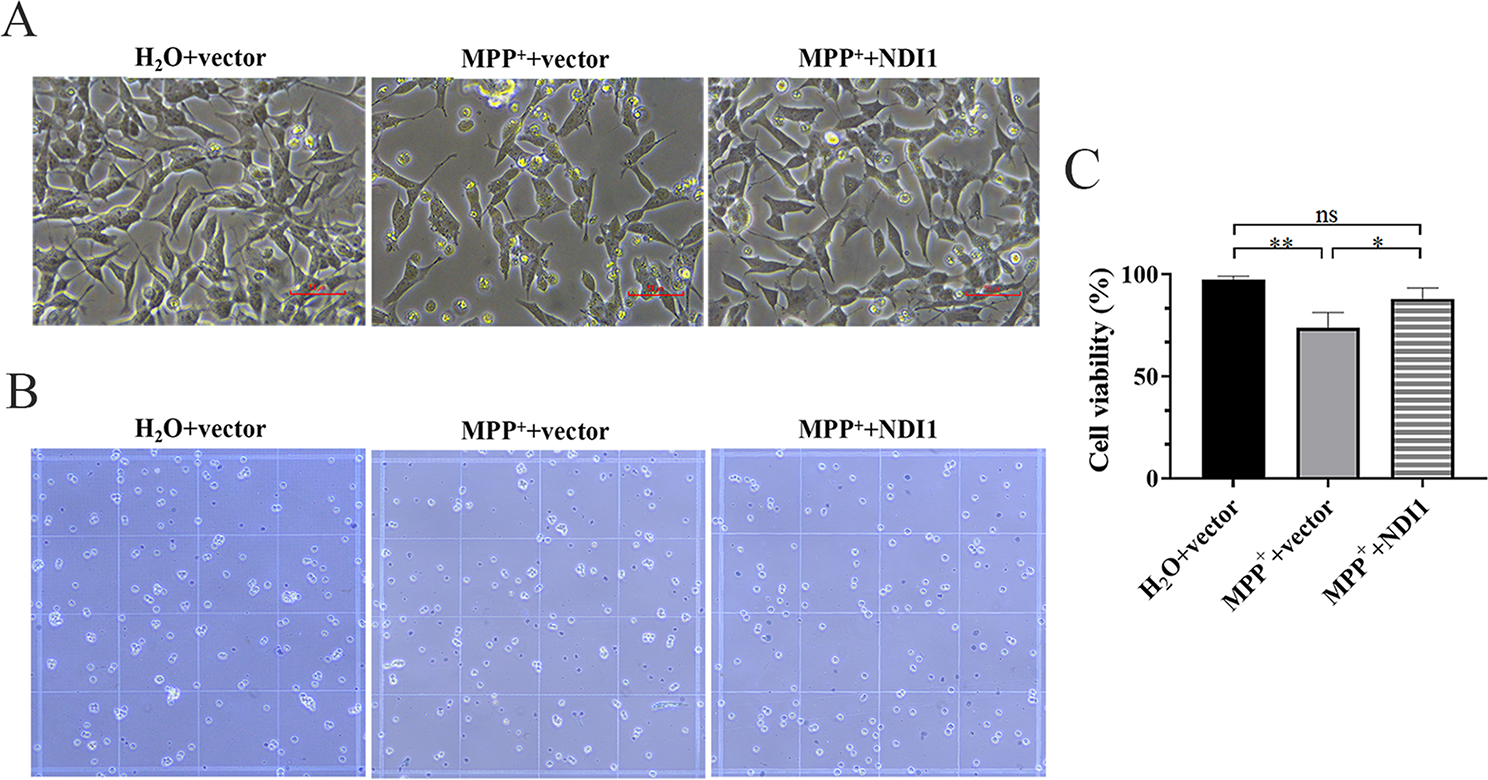
MSI2 Deficiency Represses the Proliferation, Migration and Invasion of CRC Cells In VitroTo clarify how MSI2 modulates CRC functions, we first examined the expression of MSI2 mRNA and protein in CRC cell lines and found that the MSI2 protein was upregulated in SW620 and LOVO cells but downregulated in HT29 cells (Fig. S1A and B). Therefore, we constructed stable cell lines and verified the expression of MSI2 protein and mRNA by Western blotting and qRT‒PCR, respectively (Fig. 2A and B). Furthermore, by using DepMap analysis of the CCLE datasets, the gene effect scores of MSI2 were also assessed in colorectal cancer cell lines and revealed that almost all the CRC cell line scores were less than 0 (Fig. S1C), and these negative scores implied cell growth inhibition and/or cell death following MSI2 gene knockout in each cell line. To determine whether MSI2 affects CRC cell proliferation, we first performed CCK-8 assays to assess the viability of stable cells after MSI2 knockdown in SW620 and LOVO cells or overexpression in HT29 cells and found that MSI2 increased the viability of stable cells in vitro (Fig. 2C). The cell cloning ability was further detected by colony formation assays, and the results indicated that MSI2 deficiency significantly suppressed the proliferation and colony formation of stable cells in vitro (Fig. 2D and E). Moreover, in vitro cell migration and invasion assays were performed to explore whether MSI2 knockdown could inhibit the migration and invasion of CRC cells. And the migration and invasion of tumor cells were markedly suppressed after MSI2 knockdown in SW620 and LOVO stable cells but significantly promoted in HT29 stable cells upon MSI2 overexpression (Fig. 2F and G). Mitochondrial membrane potential (MMP) is the key indicator of mitochondrial function that controls cell viability and death. We further used the MMP Assay Kit with JC-1 to detect the monomer and aggregates of MMP by FACS to quantify MMP changes after MSI2 knockdown. The percentages of JC-1 aggregates (PE+) in stable SW620 and LOVO cells were clearly decreased after MSI2 knockdown, whereas JC-1 monomer (PE−) percentages were increased (Fig. 2H and I). Consistent with this finding, an immunofluorescence assay with JC-1 staining of stable SW620 and LOVO cells confirmed that the JC-1 aggregate (PE+) intensity was decreased after MSI2 knockdown; the shift from aggregates to monomers indicates the loss of MMP in MSI2-deficient CRC cells (Fig. 2J and K). Taken together, these data suggest that MSI2 deficiency decreases the viability and inhibits the proliferation, migration and invasion of CRC cells in vitro.
Fig. 2
MSI2 deficiency inhibits the proliferation, migration and invasion of CRC cells in vitro. A Western blotting for MSI2 expression in SW620, LOVO, and HT29 stable cells. B MSI2 mRNA expression was determined by qRT‒PCR in SW620, LOVO, and HT29 stable cells. C, The cell viability and proliferation were monitored by CCK-8 at 0–96 h in SW620, LOVO, and HT29 stable cells. D-E Representative images and statistical analysis of the colony formation assay of stable cell lines showed that MSI2 deficiency significantly reduced colony formation. F-G Representative images and statistical analysis of Transwell migration and invasion of SW620, LOVO, and HT29 stable cell lines showed that MSI2 deficiency significantly inhibited migration and invasion. H FACS analysis of the mitochondrial membrane potential assay with JC-1 showed that MSI2 deficiency significantly increased JC-1 monomer (PE− fluorescence) percentages. I Statistical FACS analysis percentages of JC-1 monomers (PE− fluorescence) and JC-1 aggregates (PE+ fluorescence) in SW620 and LOVO stable cells. J Representative IFC images of the mitochondrial membrane potential assay with JC-1 in SW620 and LOVO stable cells. Blue: DAPI, red: JC-1 (PE), green: JC-1 (FITC); Scale bars, 200 μm. K Statistical IFC analysis of JC-1 aggregates and JC-1 monomers in SW620 and LOVO stable cells. These results are presented as the mean ± SD values; *p < 0.05, ***p < 0.001, ****p < 0.0001; B, C, E, G, I, K unpaired 2-tailed Student’s t test
MSI2 Deficiency Promotes Ferroptosis in CRC Cells in VitroAs we described above, MSI2 was positively correlated with ferroptosis inhibitor genes, promoting cell proliferation and suppressing MMP collapse. We wondered what effect MSI2 might have on iron death in CRC cells. We first examined the total reactive oxygen species (ROS) level of stable SW620 and LOVO cells by H2DCFDA staining and FACS analysis, we found that the total ROS levels were increased after MSI2 knockdown in SW620 and LOVO stable cells (Fig. 3A). As an iron-dependent form of cell death, ferroptosis is triggered by the accumulation of toxic lipid reactive oxygen species, particularly lipid hydroperoxides [51]. Next, the intracellular lipid ROS level was further detected using C11-BODIPY staining and FACS analysis. Notably, the lipid ROS levels were also significantly increased after MSI2 knockdown in SW620 and LOVO stable cells (Fig. 3B). Furthermore, to confirm the effect of MSI2 on cell death process, we used FACS to examine the cell mortality rates of stable SW620 and LOVO cells following incubation with different concentrations of erastin, an inducer of ferroptosis, for 12 h. We observed an increased percentage of cell death in the SW620 and LOVO MSI2 knockdown groups stable cell lines after erastin treatment (Fig. 3C), which indicates that MSI2 deficiency promotes erastin-induced iron death. However, it was found that there was no significant difference in SW620 and LOVO cell mortality between the SC and SH groups without any treatment (Fig. S2A), as well as the mRNA expression levels of apoptosis-related genes after MSI2 knockdown in SW620 and LOVO stable cells, including BCL2, BAX, CASP3 and CASP8 (Fig. S2B). And there was also inconsistency in the expression association analysis between MSI2 and pro-apoptotic and anti-apoptotic genes, such as BCL2 and CASP3 (Fig. S2C). These findings are consistent with a recent study that the loss of MSI2 does not affect the apoptosis of CRC cells [31]. More importantly, Ferrostatin 1 (Fer-1) is a potent inhibitor of ferroptosis, which can inhibit erastin-induced ferroptosis and reduce the accumulation of lipid peroxides. Therefore, FACS analysis of SW620 and LOVO stable cells showed that erastin and MSI2-induced cell iron death could be reversed by ferroptosis inhibitor Fer-1 (Fig. 3D), further indicating that MSI2 deficiency triggers CRC ferroptosis rather than cell apoptosis. Moreover, the cell viability was remarkably reduced in the SW620 and LOVO MSI2 knockdown groups after incubation with increasing dose of erastin. Conversely, the cell viability was significantly increased in HT29-OE group cells compared to NC group cells (Fig. 3E). In addition, we also determined the levels of intracellular ferrous iron and total iron in stable cells and found that the total iron and ferrous iron concentrations were both increased after incubation with different concentrations of erastin. Importantly, the intracellular ferrous iron and total iron levels were both significantly increased in SW620-SH and LOVO-SH group cells compared to SC group cells but significantly downregulated in HT29-OE group cells compared to NC group cells (Fig. 3F and G). Intracellular reduced glutathione (GSH) is an important indicator of the cellular redox state [52]. Consistent with the above finding, after treatment with different concentrations of erastin, we found that the intracellular reduced GSH levels were both significantly decreased in SW620-SH and LOVO-SH group cells compared to SC group cells but increased in HT29-OE group cells compared to NC group cells (Fig. 3H), which indicates that the cellular redox state in MSI2 knockdown cells markedly shifts from the reduced state to the oxidized state. More importantly, we used transmission electron microscopy to evaluate the mitochondrial morphology of stable cells. The SW620-SH, LOVO-SH and HT29-NC group cells had obvious mitochondrial abnormalities, and the rate of shrunken mitochondria was significantly increased after MSI2 knockdown in SW620 and LOVO stable cells, but decreased in HT29 cells upon MSI2 overexpression (Fig. 3I and J). Together, these data suggest that MSI2 deficiency triggers ferroptosis in CRC cells in vitro.
Fig. 3
MSI2 deficiency promotes the ferroptosis of CRC cells in vitro. A FACS and statistical analysis of total ROS levels (H2DCFDA) in SW620 and LOVO stable cells, gray indicates the positive control. B FACS and statistical analysis of lipid-ROS levels (C11-BODIPY) in SW620 and LOVO stable cells, gray indicates the positive control. C, FACS and statistical analysis of cell death rate in SW620 and LOVO stable cells after treatment with erastin (1–10 µg/mL) for 12 h. D, FACS and statistical analysis of cell death rate in SW620 and LOVO stable cells after treatment with erastin (5 µg/mL) and Fer-1 (10 and 20 µM) for 8 h. E The cell viability was monitored by CCK-8 and treated with different concentrations of erastin for 24 h in stable SW620, LOVO and HT29 cells. F The levels of intracellular ferrous ions were determined by treatment with different concentrations of erastin for 8 h in stable SW620, LOVO and HT29 cells. G The levels of intracellular total iron were determined by treatment with different concentrations of erastin for 8 h in stable SW620, LOVO and HT29 cells. H The levels of intracellular reduced GSH were determined by treating SW620, LOVO and HT29 stable cells with different concentrations of erastin for 8 h. I Representative images of electron microscopy revealed obvious mitochondrial abnormalities in MSI2-deficient stable cells. Scale bars, 5 μm (up) and 1 μm (bottom). J Statistical analysis of the shrunken mitochondria rate in SW620, LOVO and HT29 stable cells. These results are presented as the mean ± SD values; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001; A-H, J unpaired 2-tailed Student’s t test
MSI2 Deficiency Triggers CRC Ferroptosis by Downregulating the MAPK Signaling Cascade to Inhibit HSPB1 PhosphorylationTo determine how MSI2 regulates CRC ferroptosis, we performed 4D label-free proteomics analysis in SW620 stable cells to identify the genes corresponding to differentially expressed proteins (Fig. 4A and B). After MSI2 knockdown, the genes corresponding to differentially expressed protein included 391 downregulated genes and 460 upregulated genes, of which 265 (31.14%) were located in the nucleus, 245 (28.79%) were located in the cytoplasm, and 108 (12.69%) were located in the mitochondria (Fig. 4C-E). HSPs, which function as molecular chaperones, have been found to be involved in a variety of cellular stress, redox homeostasis and iron cell death processes, especially aberrant transduction involving the MAPK signaling pathway [33]. We then analyzed the differentially expressed HSP family genes and found that the genes encoding proteins such as HSPA5, HSPB1, TRAP1, HSF1 and HSP90 were downregulated after MSI2 knockdown, while the genes encoding proteins such as HSPH1 and HSP1AB were upregulated (Fig. 4F). Previous studies have demonstrated the pivotal inhibitory effects of HSPA5, HSPB1, HSF1 and HSPH1 in cancer ferroptosis [13, 34, 53]. Notably, HSPB1 was markedly downregulated in SW620-SH group cells following MSI2 knockdown (Fig. 4G). In addition, HSPB1, is one of the important downstream targets for MAPK signal transduction to control cell death and proliferation, GSEA enrichment analysis further revealed that the MAPK signaling pathway (NES=-1.2852) was downregulated after MSI2 knockdown (Fig. 4H). Consistent with this finding, the expression of HSPB1 was significantly positively correlated with that of MSI2 in the GEPIA CRC datasets (p = 6.2e-08, R = 0.28) (Figs. 1I and 4I). Consistent with the results from TCGA in Fig. 1G, GSEA enrichment analysis of SW620 proteomics also suggested that MSI2 was involved in the regulation of other important signaling pathways, such as oxidative phosphorylation (NES=-2.0253), Huntington’s disease (NES=-1.9115), Parkinson’s disease (NES=-1.8789) and Alzheimer’s disease (NES=-1.7749) (Fig. 4J), and recent studies have also discovered that these neuro-degenerative diseases exhibited the key features of ferroptosis: lipid peroxidation and iron accumulation [54]. More importantly, GSEA enrichment analysis of ferroptosis-related genes set suggested that the suppressor genes set (NES=-1.4683) was significantly downregulated after MSI2 knockdown, while the driver genes set (NES = 0.6627) was not significantly upregulated (Fig. 4J). Also, enriched up-regulated GO molecular functions (MFs) included cation binding, metal ion binding and ion channel binding (Fig. 4K). Moreover, the associations between MSI2 expression and multiple signaling pathways score from TCGA CRC database were further analyzed, and it was found that MSI2 positively correlated with tumor proliferation signature (p = 1.1e-06, R = 0.19), but negatively correlated with oxidative phosphorylation (p = 4.7e-39, R=-0.49), genes upregulated by ROS (p = 1.49e-20, R=-0.36), glutathione metabolism (p = 1.74e-1, R=-0.27), Fatty acid degradation (p = 9.49e-06, R=-0.18) and arachidonic acid metabolism (p = 2.04e-17, R=-0.33) (Fig. 4L). Furthermore, the expression of pro-apoptotic genes CASP3 and BAX protein was down-regulated in the SW620-SH group, while the expression of CASP8, BCL2L1 and BCL2L13 protein was up-regulated. The apoptosis signaling pathway in GSEA enrichment analysis also did not show significant differences, indicating that the apoptotic effect of MSI2 on CRC is not particularly pronounced (Fig. S2D and E).
Fig. 4
MSI2 dysregulates HSP family gene expression and multiple signal transduction processes. A Principal component analysis (PCA) plot of stable SW620-SC and SW620-SH cells in the proteomics analysis. B Pearson correlation analysis between stable SW620-SC and SW620-SH cells in the proteomics analysis. C The distribution, localization and proportion of significantly genes corresponding to differentially expressed proteins in SW620 cell proteomics. D In total, 460 upregulated protein genes and 391 downregulated protein genes were identified. E Heatmap of all the genes corresponding to differentially expressed proteins identified by proteomics. F Heatmap of differential HSP family protein genes identified in proteomics. G Circular heatmap demonstrating the expression of the most differentially expressed HSP family protein gene in proteomics. H The downregulated Gene set enrichment analysis (GSEA) of KEGG pathway was enriched in the MAPK signaling pathway. I, The positive correlation between MSI2 expression and HSPB1 expression was analyzed by Spearman correlation analysis and normalized by GAPDH in the GEPIA COAD and READ database (p = 6.2e-08, R = 0.28), n = 367. J GSEA analysis of differential proteins revealed the most significantly downregulated KEGG pathways, such as oxidative phosphorylation, Huntington’s disease, Parkinson’s disease, Alzheimer’s disease and ferroptosis suppressor genes set, while the ferroptosis driver genes set was upregulated, and ferroptosis-related genes set were obtained from FerrDb databases. K The up-regulated GO molecular functions (MFs) were enriched in metal ion binding and ion channel binding. L Associations between MSI2 expression and aberrant signaling pathway score from TCGA CRC datasets (n = 620), such as the positive correlation with tumor proliferation signature (R = 0.19), the negative correlations with genes upregulated by ROS (R=-0.36) and glutathione metabolism (R=-0.27)
Next, to fully elucidate the underlying mechanism by which MSI2 regulates ferroptosis, COG/KOG and KEGG pathway enrichment analyses were performed to assess the dysregulation of signal transduction. Among the differentially expressed genes, 26 genes were involved in lipid transport and metabolism processes, and 16 genes were involved in inorganic ion transport and metabolism processes (Fig. 5A). KEGG pathway analysis also showed the other abnormal enrichment of non-alcoholic fatty liver disease, oxidative phosphorylation, glutathione metabolism and chemical carcinogenesis-reactive oxygen species pathways (Fig. 5B and C). Through FerrDb database and SW620 cells proteomics, we further analyzed the expression of ferroptosis driver and suppressor genes and found that the driver genes, such as HMOX1, ACSL4, ACSF2, ATG5, EMC2 and GOT1, were upregulated after MSI2 knockdown. However, the suppressor genes, such as HSPB1, HELLS, BRD4, AKR1C2, JUN, SRC, GPX4 and FTH1, were downregulated after MSI2 knockdown (Fig. 5D and E). More importantly, the identified differentially expressed genes were further validated by Western blotting, and the expression of HSPB1, p-HSPB1(Ser78), FTH1 and GPX4 was decreased after MSI2 knockdown, while the expression of TFRC and ACSL4 was increased (Fig. 5F). The downregulation of phosphorylated HSPB1 (Ser78) was further confirmed by immunofluorescence in HT29-NC, SW620-SH and LOVO-SH group cells (Fig. 5G). Consistently, the mRNA levels of HSPB1, FTH1 and GPX4 were decreased after MSI2 knockdown, while ACSL4 levels were increased (Fig. 5H). We wondered whether MSI2 could directly interact with HSPB1 protein, and the co-immunoprecipitation results showed that there was no direct interaction between MSI2 and HSPB1 at the protein level (Fig. 5I). Moreover, the PPI networks of HSPB1 were collected from STRING, and HSPB1 was found to be closely linked with MAPK family genes, such as MAPKAPK2, MAPK1, MAPKAPK3 and MAPK13 (Fig. 5J). Previous studies have revealed the important functions of MAPKAPK2-HSPB1 axis phosphorylation in tumor progression and oxidative stress injury [46,47,48,49]. Similarly, as shown in Figs. 4H and 5B and C, abnormal signal transduction also occurred in the MAPK signaling pathway. Therefore, we further evaluated the differentially expressed MAPK signal transduction genes by proteomics analysis of SW620 stable cells, and the protein expression of MAPKAPK2 and MAPK13 was significantly decreased after MSI2 knockdown (Fig. 5K). Additionally, evidence for crosstalk between MSI2 and p-ERK(P42/44) has been observed in pancreatic cancer and leukemic cells [55, 56]. Intriguingly, we next determined the direct interaction between MSI2 and p-ERK(P42/44) in CRC cells and HEK293T cells by co-immunoprecipitation (Fig. 5L). And the downregulation of p-ERK(P42/44) level was further validated in MSI2 knockdown stable CRC cells by IFC (Fig. 5M). Finally, we examined and confirmed by Western blotting that the protein expression of p-ERK(P42/44), p-P38, MAPKAPK2, p-MAPKAPK2, MAPK13 and PCNA was decreased in stable cells after MSI2 knockdown (Fig. 5N). In conclusion, these findings suggest that MSI2 deficiency triggers CRC ferroptosis by downregulating the MAPK signaling cascade to inhibit HSPB1 phosphorylation.
Fig. 5
MSI2 deficiency triggers CRC ferroptosis by downregulating the MAPK signaling cascade to inhibit HSPB1 phosphorylation. A The functions of genes corresponding to differentially expressed proteins were annotated based on KOG/COG, and the main enriched metabolic pathways were lipid transport and metabolism and inorganic ion transport and metabolism. B-C Genes corresponding to the differentially expressed proteins were functionally annotated based on the KEGG database and were mainly enriched in nonalcoholic fatty liver disease, oxidative phosphorylation, chemical carcinogenesis-reactive oxygen species and MAPK signaling pathway. D Circular heatmap demonstrating the most significant differential expression of ferroptosis-related genes in stable SW620 cell proteomics analysis. E Heatmap showing the differential expression of ferroptosis-related driver or suppressor genes in stable SW620 cell proteomics analysis. F Verification of differential gene expression in SW620, LOVO, and HT29 stable cells by using Western blotting for MSI2, HSPB1, p-HSPB1(Ser78), ACSL4, FTH1, TFRC and GPX4. G Representative IFC images of p-HSPB1(Ser78) expression in SW620, LOVO and HT29 stable cells. Blue: DAPI, red: p-HSPB1(Ser78); Scale bars, 50 μm. H HSPB1, ACSL4, FTH1 and GPX4 genes mRNA expression were determined by qRT‒PCR in SW620, LOVO, and HT29 stable cells. I Co-immunoprecipitation assays showed that there was no direct interaction between MSI2 and HSPB1 in SW620 and LOVO cells. FT means flow through wash fraction, input and anti-IgG as controls. J The protein–protein interaction (PPI) networks between HSPB1 and MAPK family genes were constructed by STRING. K Differential MAPK signaling pathway genes expression in stable SW620 cell proteomics analysis. L Co-immunoprecipitation assays showed that MSI2 could interact with p-ERK(p-P42/44) in SW620, LOVO and HEK293T cells. The input and anti-IgG as controls. M The protein levels of p-ERK(p-P42/44) were determined by IFC in SW620, LOVO and HT29 stable cells. Blue: DAPI, green: p-ERK(p-P42/44); Scale bars, 50 μm. N Western blotting for MAPK signaling cascade gene expression, such as ERK(P42/44), p-ERK(p-P42/44), P38, p-P38, MAPKAPK2, MAPK13, p-MAPKAPK2 and PCNA, in SW620, LOVO, and HT29 stable cells. These results are presented as the mean ± SD values; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001; H unpaired 2-tailed Student’s t test
MSI2 Deficiency Restrains CRC Malignancy by Inhibiting MAPK/HSPB1 axis Phosphorylation In VivoTo evaluate the effect of MSI2 on the biological behavior of CRC in vivo, we first established a CRC cell line xenograft model in M-NSG mice. We observed that the tumors formed by LOVO cells with stable MSI2 knockdown were consistently smaller and lighter than those formed by control cells, while tumors formed by HT29 cells with stable MSI2 overexpression showed the opposite trends, indicating that MSI2 knockdown can effectively inhibit tumor growth in vivo (Fig. 6A-C). Next, we used MSI2 knockout (MSI2-/-) transgenic and wild-type (MSI2+/+) mice to construct a colitis-associated colon cancer model by administering azoxymethane (AOM) intraperitoneally and feeding with 3% DSS for three cycles (Fig. 6D). Consistent with the above findings, deletion of MSI2 markedly reduced the occurrence and number of tumors in the mouse colon (Fig. 6E). Analysis of IHC staining results for MSI2, HSPB1, p-HSPB1(Ser78) and Ki67 showed that MSI2 knockout could reduce the expression of HSPB1, p-HSPB1(Ser78) and the tumor proliferation capacity in transgenic CAC mice (Fig. 6F). Moreover, we performed histological staining analysis of xenografted mouse tumors, including H&E staining and IHC of SW620, LOVO and HT29 stable cells. Similarly, the IHC results showed that MSI2 knockdown significantly decreased the expression of HSPB1, p-HSPB1(Ser78), p-P38, Ki67 and PCNA and increased the expression of ACSL4 in SW620 and LOVO stable cells. Conversely, overexpression of MSI2 markedly increased the expression of HSPB1, p-HSPB1(Ser78), p-P38, Ki67 and PCNA and decreased the expression of ACSL4 in HT29 stable cells (Fig. 6G and H). In addition, we investigated the role of MSI2 in CRC metastasis in vivo. The establishment of CRC liver metastasis models was performed by injecting SW620 and LOVO stable cells into the M-NSG mouse spleen, and MSI2 knockdown significantly inhibited the liver metastasis of stable SW620 and LOVO cells, which was further confirmed by histological analysis (Fig. 6I-K). Similarly, the CRC lung metastasis model was constructed by injecting SW620 and LOVO stable cells into the tail veins of M-NSG mice, and the results showed that MSI2 knockdown also inhibited lung metastasis of stable SW620 and LOVO cells (Fig. 6L-N). All these data reveal that MSI2 deficiency suppresses the growth and metastasis of CRC in vivo by inhibiting MAPK/HSPB1 axis phosphorylation.
Fig. 6
MSI2 deficiency suppresses CRC malignant features, including tumor proliferation and metastasis, by inhibiting MAPK/HSPB1 axis phosphorylation in vivo. A In vivo models of xenograft tumors from HT29 and LOVO stable cells were established in M-NSG mice, n = 6. B Quantitative analysis of xenografted tumor volume change from HT29 and LOVO stable cells, n = 6. C, Statistical analysis of xenografted tumor weight from HT29 and LOVO stable cells, n = 6. D Procedure for constructing colitis-associated colon cancer in vivo models of MSI2+/+ and MSI2−/− transgenic mice. E Representative image and statistical analysis of colitis-associated colon cancer tumor number in MSI2+/+ and MSI2−/− transgenic mice, n = 5. F Representative H&E, IHC images and statistical analysis of the IHC score (IOD per area) of MSI2, HSPB1, p-HSPB1(Ser78) and Ki67 in MSI2+/+ and MSI2−/− transgenic CAC mice model. Scale bars, H&E, 200 μm; IHC, 100 μm. G Representative H&E and IHC images of xenograft tumors from SW620, LOVO and HT29 stable cells, including MSI2, HSPB1, p-HSPB1(Ser78), p-P38, ACSL4, Ki67 and PCNA. Scale bars, H&E, 200 μm; IHC, 100 μm. H Statistical analysis of the IHC score (IOD per area) of MSI2, HSPB1, p-HSPB1(Ser78), p-P38, ACSL4, Ki67 and PCNA from SW620, LOVO and HT29 stable cells xenograft tumors. I-J Representative images and statistical analysis of liver metastasis nodules from SW620 and LOVO stable cells in M-NSG mice, n = 5. K Representative images of H&E staining of liver metastasis from SW620 and LOVO stable cells. The yellow star indicates the tumor area. Scale bars, 200 μm. L-M Representative images and statistical analysis of lung metastasis nodules from SW620 and LOVO stable cells in M-NSG mice, n = 5. N Representative images of H&E staining of lung metastasis from SW620 and LOVO stable cells. The dark star indicates the tumor area. Scale bars, 400 μm. These results are presented as the mean ± SD values; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001; B, C, E, F, H, J, M unpaired 2-tailed Student’s t test
HSPB1 Rescues the Phenotypes of MSI2 Deficiency on CRC Ferroptosis In Vitro and In VivoTo further address the role of the MSI2-HSPB1 axis in CRC ferroptosis, we investigated and determined whether the phenotypes of MSI2 deficiency could be reversed by rescuing HSPB1 expression during CRC ferroptosis. We first transfected the control vector and HSPB1 plasmid into SW620 and LOVO MSI2 knockdown cells to restore HSPB1 expression. The Western blotting results revealed that the protein expression of HSPB1, FTH1, PCNA and GPX4 was increased following rescue with the HSPB1 plasmid, but the expression of ACLS4 was decreased (Fig. 7A). Consistently, the mRNA expression levels of HSPB1, FTH1 and GPX4 were both upregulated after HSPB1 rescue, while ACLS4 expression was downregulated (Fig. 7B). Next, colony formation assays were performed to assess the capacity of CRC cell growth, and the results showed that HSPB1 rescue could improve the proliferation and colony formation ability of SW620 and LOVO MSI2 knockdown cells (Fig. 7C). Furthermore, we also examined intracellular total ROS and lipid-ROS levels after HSPB1 restoration in SW620 and LOVO MSI2 knockdown cells and found that the restoration of HSPB1 in MSI2 knockdown cells markedly reduced the levels of intracellular total ROS and lipid-ROS compared with those in the vector and untreated group (Fig. 7D and E). Similarly, the restoration of HSPB1 expression in SW620 and LOVO MSI2 knockdown cells also significantly reduced the cell death rate induced by erastin (Fig. 7F). More importantly, compared with those in the vector and untreated groups, the levels of intracellular ferrous iron and total iron were both decreased in the rescue group cells after HSPB1 transfection (Fig. 7G and H). However, in contrast, the intracellular reduced GSH levels were significantly increased in the rescue SW620-SH and LOVO-SH group cells compared to the vector group cells (Fig. 7I). In addition, by transmission electron microscopy, we observed that the mitochondrial morphology of the rescue group cells gradually recovered, and the proportion of shrunken mitochondria was significantly reduced after HSPB1 restoration in both SW620 and LOVO MSI2 knockdown stable cells (Fig. 7J and K). Moreover, we investigated whether restoring HSPB1 expression could reverse the tumor growth trend in vivo. Therefore, to validate our hypothesis, a xenograft tumor model in M-NSG mice was further constructed by injecting SW620 stable cells transfected with HSPB1, and we observed that restoration of HSPB1 expression significantly promoted tumor growth in vivo, including tumor volume and weight (Fig.
留言 (0)