
記住我



In this work, we generated induced pluripotent stem cell (iPSC)-derived neurons from 4 non-familial, early-onset Alzheimer’s disease (EOAD) patients (age at onset (AAO) 51–56 years) and 4 non-demented control (NDC) subjects whose age at biopsy was 76–82 years. Samples were provided by the UC San Diego Alzheimer’s Disease Research Center (ADRC). Individual clones (n = 3) from all 8 subjects were transformed into iPSCs and differentiated into neurons as previously described [6]. RNA was extracted from frozen neuron pellets (Fig. 1A). All 4 EOAD patients displayed diffuse cerebral atrophy by Magnetic Resonance Imaging (MRI) and decreased cognition as evidenced by either Mini-Mental State Examination (MMSE) scores less than 15 [7] or Montreal Cognitive Assessment (MoCA) score less than 25 [8], indicating a moderate to an advanced stage of EOAD progression (Fig. 1B). To the best of our knowledge, this is the first report using iPSC-derived neurons to model non-familial EOAD. RNA-seq was used to characterize gene expression dysregulation in EOAD and assess possible underlying mechanisms.
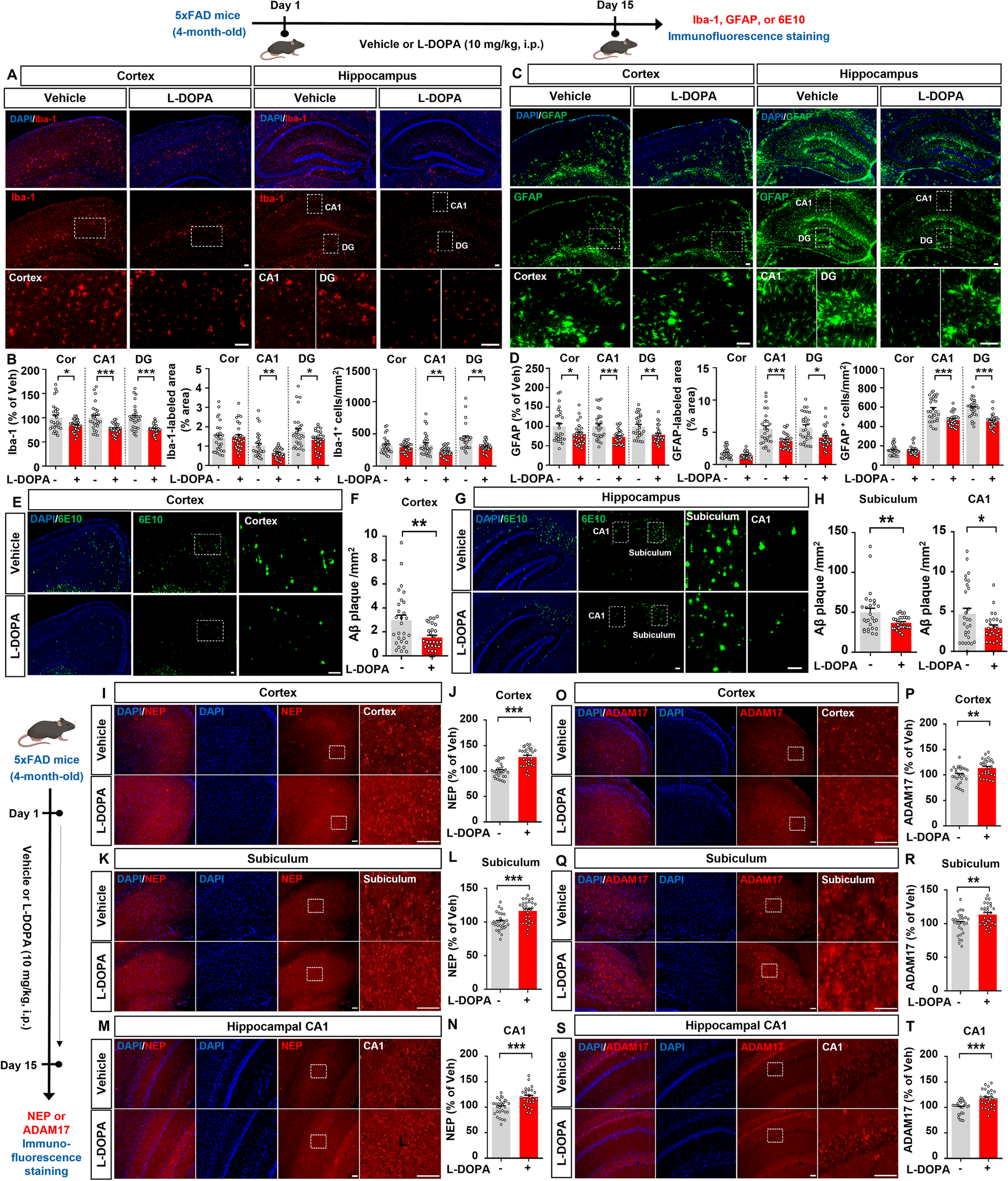
Fig. 1
A Non-demented controls (NDCs) and EOAD iPSCs were differentiated using dual SMAD inhibitors (first step) and basic-FGF withdrawal (second step). B Metadata for NDC and EOAD subjects used in this study. C Multi-dimensional scaling (MDS) analysis after batch correction by experimental condition, sex and sequencing batch of filtered normalized RNA-seq data. D RNA-seq volcano plot of differentially expressed genes (DEGs) for EOAD patients relative to all NDCs as determined by kimma with an FDR p-value < 0.05. E Quasi-proportional Venn diagram overlap of DEGs across the four EOAD patients relative to all NDC subject neurons. F MDS (left) and UMAP (right) analysis based on NDC and EOAD neurons generated in this study using filtered, normalized RNA-seq data. G RNA expression profile heatmap corresponding to all filtered genes. H MDS (left) and UMAP (right) analysis based on NDC and EOAD neurons generated in this study using filtered, normalized RNA-seq data for neuron lineage genes. I RNA expression profile heatmap corresponding to either neuron lineage genes (left) or post-mitotic genes (right) clustered by subject experimental condition (NDC or EOAD)
RNA isolated from EOAD and NDC neurons were of high quality, with RNA Integrity Numbers (RIN) ranging from 7.8 to 9.8. We assessed how the neurons from EOAD patients clustered with respect to NDC subjects in multi-dimensional scaling (MDS) space following correction for sex, sequencing batch, and experimental condition. This revealed no clear clustering of NDC or EOAD samples but rather a slight overlap between them (Fig. 1C). This may not be surprising, as EOAD displays heterogeneity in clinical presentation [2]. Differential expression analysis between two conditions revealed a small number of differentially expressed genes (DEGs) (n = 14) using the same covariates in the linear design model (Fig. 1D). To determine whether heterogeneity between the EOAD patients was causing the low number of DEGs, we compared each EOAD relative to all control subjects. While EOAD patient #3 displayed a higher amount of DEGs relative to all NDC subjects (n = 323), all other individual EOAD patients demonstrated a similar magnitude of DEGs (Additional file 1: Fig. S3A) with only 2 common DEGs among them (Fig. 1E). Next, we sought to assess the variance within individual clones of the same subject, regardless of EOAD or NDC, in MDS space for all filtered genes; we observed sparse grouping within each subject (less in EOAD but more in NDC) and slight overlap between the two groups (Fig. 1F, left). To assess whether a dimension reduction approach would reveal a better separation of EOAD and NDC neurons, we performed UMAP on the normalized, filtered counts. Here, we observed similar clustering within replicates of the same EOAD samples but worse clustering for the NDC samples; this could suggest variable differentiation of clones within patient samples as well as across samples during the iPSCs differentiation process into neurons (Fig. 1F, right). Furthermore, examining heatmap and dendrogram clustering of the overall expression profile of the filtered genes (n = 21,546) demonstrates that the expression patterns from both EOAD and control neurons did not show apparent differences, with no distinct patterning observed between the two groups (Fig. 1G).
To assess the neuronal differentiation state of the EOAD and NDC neurons, we first focused on genes that regulate neuron lineage (curated from GO: Biological Process and Reactome genesets, n = 1078). MDS revealed a slightly greater variance within the EOAD and NDC conditions but no visible separation between the two (Fig. 1H, left). When performing UMAP analysis, while we observe a single cluster of the majority of EOAD patient clones, there is greater separation of NDC clones within and between NDC subjects overall, suggesting a variable neuron lineage state across all samples (Fig. 1H, right). This suggests that iPSC-derived EOAD neurons do not show a similar separation from healthy neurons as observed in familial Alzheimer’s disease (FAD) neurons with PSEN1, PSEN2, and APP mutations [4, 5].
Next, we sought to assess the expression profile of these neurons based on only neuron lineage genes. We saw a similar trend to the findings reported above: no noticeable gene expression differences observed in key marker genes for both EOAD and NDC neurons (Fig. 1I, left). As such, gene expression differences between NDC and EOAD were not well captured under the conditions of our study. We then observed a subset of these neuron lineage genes that mainly regulate post-mitotic neuron maturation (NEFL, SOX2, UCHL1, TUBB3, MAP2, DCX, NEUROD1 and APOE) [9,10,11,12,13] (Fig. 1I, right). These genes were selected since they are the most common markers for post-mitotic neurons found in literature [9,10,11,12,13] and we observe similar expression profiling for both NDC and EOAD neurons.
Since neither EOAD nor NDC samples clustered together, we next assessed whether EOAD and NDC neurons have established a mature neuronal identity [14] or whether there may be alternative cell lineages in our cultures by first looking at different neural lineage stages ranging from neural progenitors, glia, and neurons (immature and mature) derived from human post-mortem brain and patient-derived iPSC single-cell mean expression data [9, 12, 15,16,17,18]. When we interrogated marker genes specific to these major cell types via gene-level expression across EOAD and NDC neurons, we observed the highest average marker gene expression associated with neurons, followed by glial and progenitor cells, but without statistically significant expression differences between EOAD and NDC (Additional file 1: Fig. S1A). When using all marker genes, we observe a slight decrease in average gene-level expression when comparing expression data from all EOAD relative to all NDC neurons (Additional file 1: Fig. S1A). It is worth noting that the potential existence of populations of these earlier lineage cell types could contribute to the relative variability seen in our iPSC-derived EOAD and NDC neuron cultures. We then proceeded to look at the average gene-level expression of different EOAD and NDC subject lines using cell type markers from categories such as progenitor cells, glial cells, immature neurons, and mature neurons (Additional file 1: Fig. S1B–E) and cell subcategories such as neural progenitor cells (NPCs), oligodendrocytes, excitatory neurons, and inhibitory neurons (Additional file 1: Fig. S2D–G). For all cell types and subcategories, we find no significant differences between EOAD and NDC neurons (Additional file 1: Figs. S1B–E, S2D-G). Next, we performed clustering in MDS space within the different cell type classes to see if the separation of the two conditions changed with marker gene expression of different stages of neuronal lineage; we observed tighter clustering within experimental condition (EOAD or NDC) despite slight overlap for glial cells (Additional file 1: Fig. S1G) followed by mature neurons (Additional file 1: Fig. S1I), immature neurons (Additional file 1: Fig. S1H), and progenitor cells (Additional file 1: Fig. S1F). Furthermore, we also observe an increase in the separation of EOAD and NDC neurons along dimension 1 in MDS space for mature neuron marker genes (Additional file 1: Fig. S1I), providing evidence of the heterogeneity involved. Looking closely at different groups of cellular subtype markers (Additional file 1: Fig. S2A–C) across different EOAD patients relative to all NDC subjects, we observe no significant expression differences in any cellular subtype (Additional file 1: Fig. S2D–G). When we observe these cellular subtypes in MDS space, there is tighter clustering involved within the experimental condition, despite the slight overlap in marker gene expression for neural progenitor cells (Additional file 1: Fig. S2H) and oligodendrocytes (Additional file 1: Fig. S2I) when compared to excitatory and inhibitory neurons (Additional file 1: Fig. S2J, K) which are clustered more sparsely. This suggests that heterogeneity increases as EOAD and NDC iPSCs progress to a more mature state characterized by excitatory and inhibitory neurons.
Using a standard protocol for making human iPSC-derived neurons, we have not effectively discriminated between NDC and EOAD neurons. Our findings suggest that this approach for examining the biology of EOAD may fail either to adequately differentiate EOAD and NDC neurons or capture changes in gene expression characteristic of AD. The lack of patient clustering and variance observed in the EOAD study may be because diverse etiological factors contributing to aging-associated epigenetic changes (e.g., RNA modifications and non-coding RNA regulation) [19] are not preserved during iPSC reprogramming [20]. We also investigated the expression levels of marker genes associated with major alternative cell types and their cell subtypes to determine whether they contribute to the EOAD and NDC neuron cell culture; we quantified average expression levels and MDS clustering with no adjustments made to account for the presence of variable neuronal populations, which revealed an increasing separation between EOAD and NDC cultures for mature neuron marker genes. Furthermore, we can suggest that the differentiation protocol via SMAD inhibition and bFGF removal used to generate the EOAD and NDC neurons did not ultimately result in a purely differentiated mature neuron population. It is likely that there is a mix of different neuronal lineages, including those from an earlier lineage state (more specifically progenitor cells) and glial cells (not necessarily representative of oligodendrocytes, but likely representative of astrocytes, microglia and OPCs), but it is unclear their relative contribution to the neuron culture. As such, we were not able to capture the phenotype differences between EOAD and NDC neurons. This is due to the following: (1) the variation of differentiation across clones of patients, (2) the variability within EOAD patients relative to NDC subjects and (3) the relative immature state of the neuron cultures.
Methods that preserve the epigenetic signatures may provide better models for studying sporadic AD. It is likely that induced neurons (iNs) that undergo direct neuronal conversion from fibroblasts via small molecule reprogramming [21] can capture a more mature neuron state of the EOAD brain and merit further studies. Previous studies have shown that age-dependent cellular programs of patients with late-onset sporadic AD derived from direct iNs are characterized by downregulation of mature neuronal properties (i.e., loss of mature neuronal fate, neuronal dedifferentiation) and upregulation of cell cycle re-entry. Additionally, age-related changes in the epigenetic landscape appear to underlie a hypo-mature neuron state in iNs, thus directing toward a more de-differentiated state of sporadic AD [14, 20]. Ultimately, utilizing an iN model may be required to preserve both EOAD gene expression and epigenetic signatures, thus informing the cellular environment required to understand the neuronal biology of EOAD.
留言 (0)