

記住我
Osteosarcoma (OS) is the most common primary malignant bone tumor in children and adolescents, affecting >1,000 patients annually in the United States alone (1–4). Osteosarcomas are characterized by a loss of mineralization (5); moreover, the majority of OS patients have subclinical micrometastases, whereas 30-40% have full-blown metastatic disease (6, 7). Currently, the standard OS treatments consist of surgical removal of the tumor followed by adjuvant chemotherapy, which causes severe side effects and chemoresistance in patients (3, 4, 8–11). Several clinical trials evaluating nonconventional OS therapies that combine immunotherapy and cytostatic drug treatments are currently underway in an effort to address this problem (11). However, despite current treatment options, 30 to 40% of OS patients still die within 5 years of diagnosis (10). The prognosis for osteosarcoma patients depends on whether it has metastasized at the time of diagnosis, which involves induction of the epithelial-to-mesenchymal transition (EMT), a process that is not only activated in cancer but also in disease-induced fibrosis (12), and non-disease processes like wound re-epithelialization and embryonic mesoderm formation (13).
Immunotherapy has thus far been ineffective against OS due to its ability to evade the immune system (14–17). It is unknown how resistance develops, but it may result from the complex immunological and/or epigenetic reprogramming of OS that creates a tumor permissive environment to support malignancy (3, 4, 10). In addition, recent studies have shown that the OS microenvironment contains activated cancer associated fibroblasts (CAFs) that can influence desmoplasia, a process in which excessive fibrosis leads to chemoresistance and metastasis of malignant cancer cells, and inhibition of immune cell infiltration that increases tumor burden (18). Furthermore, genome-wide association studies have linked genetic variations within a neurotransmitter receptor gene to increased risk of OS development, suggesting neuronal-tumor interactions in the manifestation of the disease (19). Evidence also suggests that nonsense-mediated RNA decay (NMD) is uniquely increased in OS, which, for example, decreases the number of NMD-target neoantigens on OS tumor cells that are required to elicit the cytotoxic killing of cancer cells by immune cells (14–17). NMD is part of the post-transcriptional RNA surveillance pathway to clear aberrant transcripts harboring premature termination codons (PTCs) often generated by abnormal or ineffective biogenesis of mRNAs or by somatic mutations in cancers (15, 20–23). To date, the selective targeting and modulation of the NMD pathway to overcome OS resistance for therapeutic considerations has not been reported.
Anti-cancer properties of vitamin D have been studied in major cancer types, however our knowledge of vitamin D’s impact on OS is still very limited. Vitamin D deficiency is a global health issue, with the lowest levels linked to more advanced cancers (24–32). Clinical randomized studies have demonstrated that vitamin D supplementation reduces the risk of invasive cancer and/or mortality in subgroups of people (25, 26, 33–38), suggesting that increased circulating vitamin D plays a protective role. Vitamin D is composed of two major forms, vitamin D2 (ergocalciferol) and vitamin D3 (cholecalciferol) (30). In humans, vitamin D3 is synthesized in the skin in response to UVB exposure, while vitamin D2 is obtained from plant sources in our diets with lower efficacy. Both forms of vitamin D are biologically inert and must be converted and hydroxylated to 25(OH)D by vitamin D-25-hydroxylase in the liver, which is the primary measurement of vitamin D status (39). 25(OH)D is further hydroxylated in the kidneys or within specialized cell types by 25(OH)D-1-OHase (CYP27B1) to produce 1-alpha, 25-dihydroxyvitamin D (also known as 1,25(OH)2D), the biologically active derivative of vitamin D (30, 40–44). The 1,25(OH)2D effects are mediated by the vitamin D receptor (VDR), an intracellular nuclear receptor superfamily member, that can promote cell cycle arrest and apoptosis via post-transcriptional and gene regulatory means (40, 44–54). In mammals, the physiological and molecular effects of vitamin D3 on bone health and function have been experimentally and clinically validated (30). Vitamin D3 is known to directly promote the differentiation of normal bone-forming osteoblast-lineage cells and primary human and murine osteoblast cells in vivo (55) and in vitro (48, 51), respectively. Although numerous studies have implicated a suppressive role for vitamin D3 in invasive cancer development (e.g., breast, prostate) and improved cancer patient and animal survival (1–4, 26–30), the functional role of vitamin D3 and the VDR in OS is unclear. Uncertain is whether vitamin D3 can be utilized to address a number of crucial concerns about OS biology and therapy, such as the regulation of transformation, cancer fibrosis, NMD, and metastasis. Using both in vitro and in vivo systems, this work examined the influence of the VDR and vitamin D3 on the regulation and pathogenesis of OS through impact on OS growth, migration and potential cancer cell immunorecognition.
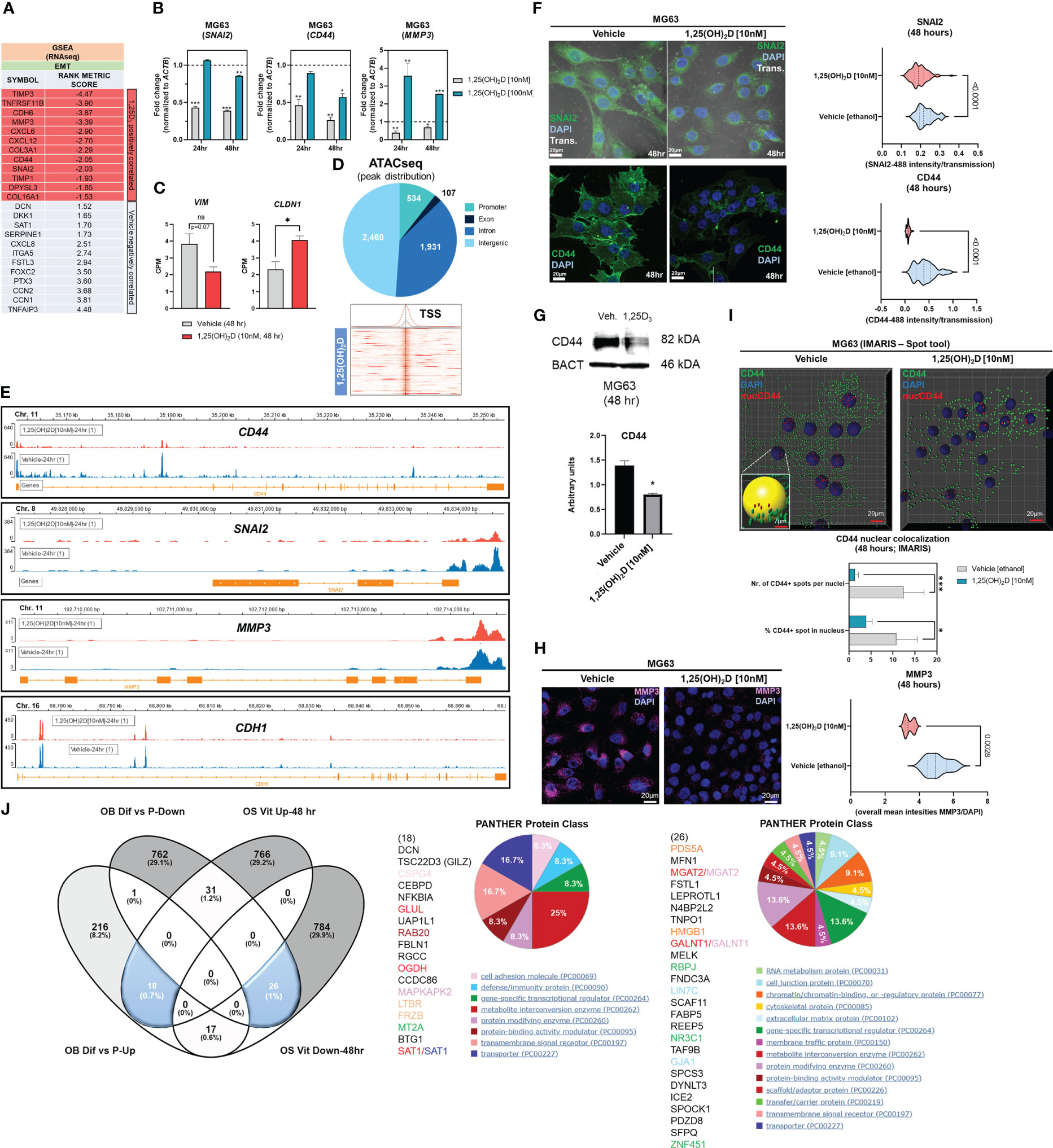
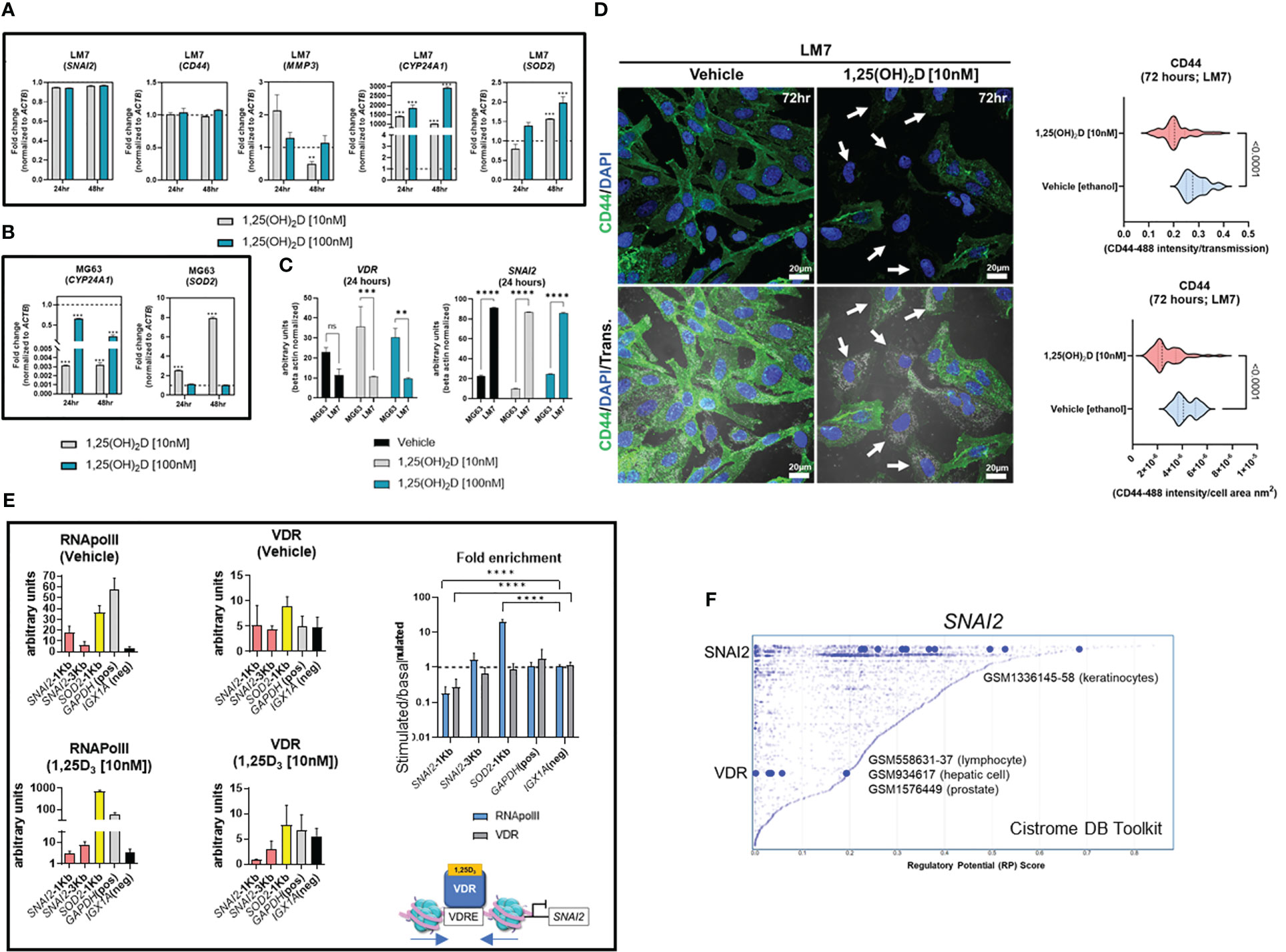
Results1,25(OH)2D promotes the enrichment of the EMT pathway in MG63 osteosarcoma cellsGene set enrichment analysis (GSEA) (56) was used to identify novel pathways using RNAseq data from MG63 OS cells treated with 1,25(OH)2D (GEO accession: GSE220948). Using the annotated gene set file M5930 from the Molecular Signatures Database (MSigDB), the EMT gene set was identified as being significantly enriched (Figure 1A) (57). Using a physiologically-relevant concentration range (10-100nM) (58), 1,25(OH)2D inhibited the expression of EMT inducers and regulators such as SNAI2 (also known as the transrepressor SLUG), CD44, and MMP3, as confirmed by quantitative PCR (qPCR) (Figure 1B). Higher 1,25(OH)2D concentrations increased MMP3, both of which are known to induce apoptosis(47,48,27). In addition, 1,25(OH)2D downregulated and upregulated, respectively, VIM and CLDN1, which were not annotated in M5930, indicating that vitamin D3 may inhibit EMT via downregulation of intermediate filaments and the promotion of cell-to-cell adhesion (Figure 1C) (59, 60). ATACseq was used to assess 1,25(OH)2D-dependent genome-wide chromatin accessibility of genes (Figure 1D), and potential promoter regulatory regions were identified within CD44 and SNAI2, but not MMP3, VIM, and CLDN1 (Figures 1E, S1A). Intriguingly, 1,25(OH)2D did not regulate the major epithelial cadherin, CDH1, which is known to be repressed by SNAI2 (61), indicating that other essential cell-cell mediators (such as claudins) are involved in OS (Figure 1E). Moreover, after 1,25(OH)2D treatment, the protein expression of SNAI2, MMP3, and CD44 all decreased (Figures 1F-H, S1B). Surprisingly, 1,25(OH)2D treatment decreased CD44 nuclear localization, indicating that 1,25(OH)2D may inhibit the proteolytic processing of CD44’s intracellular cytoplasmic carboxy-terminal domain in order to limit tumor progression and metastasis via alternative genomic interactions (Figure 1I) (62).
Figure 1 1,25(OH)2D treatment of MG63 cells results in enrichment of EMT pathway genes. (A) Gene set enrichment analysis (GSEA) was performed to identify a collection of a priori EMT-related genes that were over-represented in our RNAseq experiment and may be associated with 1,25(OH)2D anti-cancer effects. The MSigDB database was utilized for GSEA at http://www.broadinstitute.org/gsea. (B) Quantitative PCR (qPCR) study of genes associated with EMT. Tukey’s multiple comparison test, two-way ANOVA; p ≤ ****0.0001, ***0.001, **0.01, *0.05 (n=3). (C) MG63 RNA-seq study of EMT genes that are not contained in the MSigDB database. Test of independence (T); p ≤ *0.05 (n=1). (D) Top: ATAC-seq analysis and peak distribution of MG63 cells treated for 24 hours with 1,25(OH)2D. Bottom: ATAC-seq heatmap of 1,25(OH)2D regulated genes about the transcription start sites (TSS). (E) Visualization of ATAC-seq tracks utilizing the IGV genome browser in order to identify accessible chromatin areas and read peaks. Data represents 24 hours of treatment with 10nM 1,25(OH)2D in MG63 cells. All scales indicate the minimum and maximum reads in that window using IGV auto-scale. (F) Left: Immunofluorescence study of SNAI2 and CD44 in MG63 cells 48 hours after treatment with 1,25(OH)2D. Bars = 20µm. Right: Quantitative study of SNAI2 and CD44-coupled fluorescence intensities in MG63 samples treated with 1,25(OH)2D versus vehicle. Individual cell areas and transmitted light were used to equalize intensities. Between the vehicle and treatment data sets, an unpaired t test was conducted, with two-tailed p-value summaries displayed in the graphs. The violin plots (n=4) depict the quartiles and medians (darker dashes). (G) Western blot study of CD44 utilizing MG63 cells treated for 24 hours with 10nM 1,25(OH)2D. To facilitate comparisons, LiCOR band intensities were converted to arbitrary units. Test of independence; p ≤ *0.05 (n=3). (H) Left: Immunofluorescence confocal imaging study of MG63 cells reveals a decrease in MMP3 expression 48 hours after treatment with 10nM 1,25(OH)2D. Right: The violin plots reflect n=4 experimental conditions with an average of 20-40 cells (Unpaired T test). (I) An IMARIS colocalization analysis of MG63 cells treated for 48 hours with 10nM 1,25(OH)2D. Compared to 1,25(OH)2D-treated controls, CD44 nuclear localized expression was more prevalent in untreated controls. Two-way ANOVA Multiple comparisons using Tukey’s test; p ≤ ***0.001, *0.05 (n=3). (J) Left: A comparative transcriptome study of differentiated (Dif) and proliferating (P) normal osteoblasts in comparison to 1,25(OH)2D-treated MG63 OS cells. Normal osteoblast gene set generated from GSE39262. Set expression value (A) to 4 and cut-off to 2-fold change. Right: Panther analysis of subsets of up- and down-regulated genes. NS, not significant.
A comparison to normal osteoblast states (differentiation versus proliferation) was conducted to determine the relative biological effects of 1,25(OH)2D on OS. We compared the normal osteoblast transcriptomic data from GSE39262 with our RNAseq data from MG63 OS cells (Figure 1J). Compared to MG63 genes that were either upregulated or downregulated in response to 1,25(OH)2D, there was little overlap (0.7% and 1%, respectively) between the genes that define differentiated normal osteoblasts (Figure 1J left). Except for decorin (DCN), the remaining genes regulated by 1,25(OH)2D did not overlap with either normal proliferation or differentiation genes, indicating that one of vitamin D3’s function toward OS may involve other processes such as EMT regulation (Figure 1J middle). 1,25(OH)2D upregulated the cell adhesion factor CSPG4 and the WNT antagonist FRZB, but a greater proportion of genes (>29%) were excluded from differentiation or proliferation pathways. Similarly, several cell proliferation/DNA replication cancer-inducing genes, such as PDS5A, HMGB1, FABP5, and TAF9B, were downregulated in differentiated osteoblasts, but they represented less than 1% of the downregulated 1,25(OH)2D-dependent genes in MG63 cells, indicating that the majority of vitamin D’s effects on OS may involve functional regulation of processes such as EMT (>29%) instead (Figure 1J right).
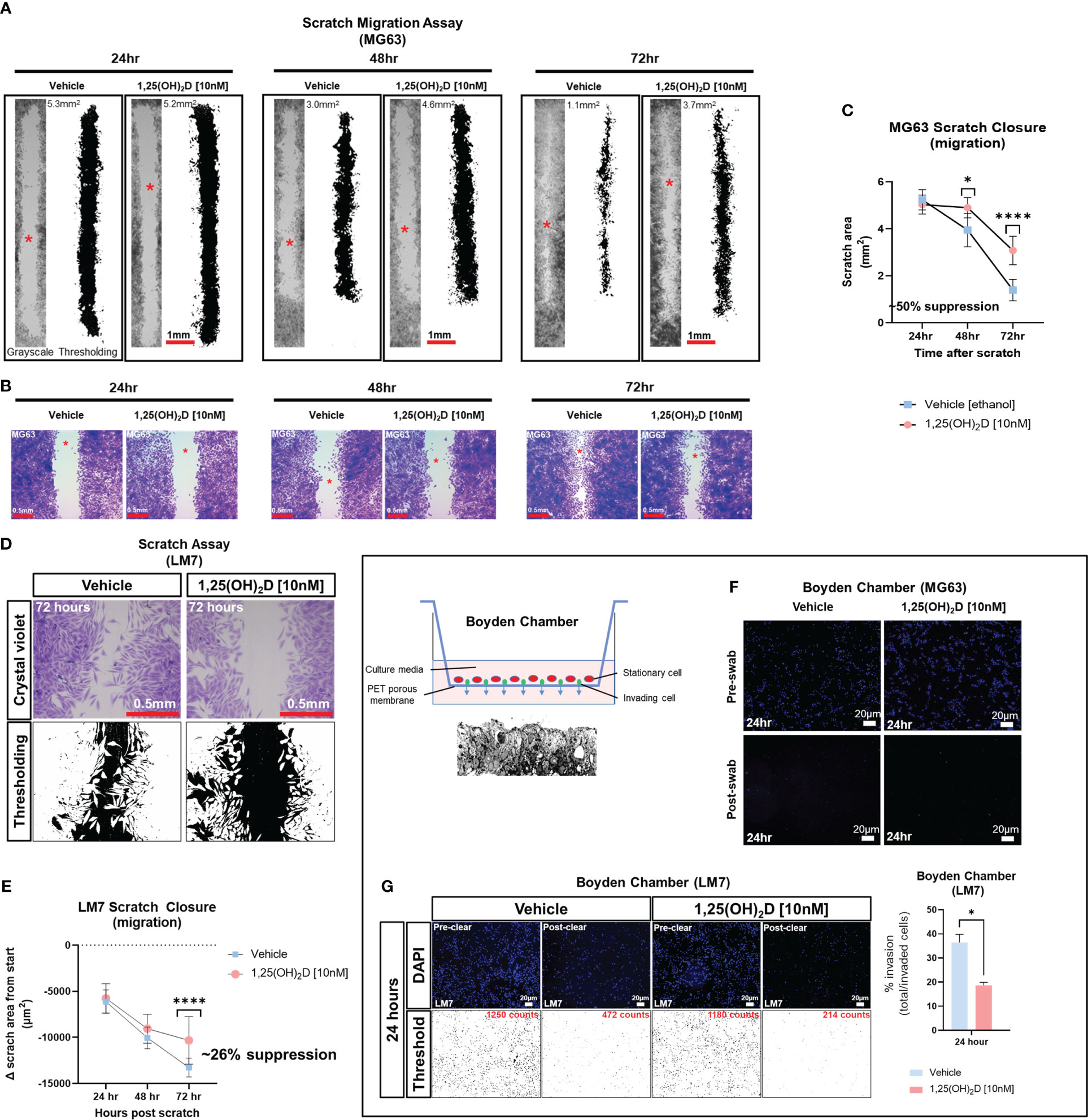
1,25(OH)2D inhibits wound induced EMT and migration of osteosarcoma cellsDespite the findings of the GSEA, the biological effects of 1,25(OH)2D on EMT was unclear. To determine the effect of 1,25(OH)2D on cell migration, we performed an in vitro scratch assay on OS cell models MG63 and LM7, which are low and highly metastatic, respectively (63). After 72 hours of 10nM 1,25(OH)2D treatment, MG63 cell migration from the leading edge was reduced by approximately 50% compared to vehicle (Figures 2A–C). In contrast, after 72 hours of treatment with 10nM 1,25(OH)2D, scratch migration of LM7 cells was inhibited by 26% (Figures 2D, E), indicating relative resistance. The Boyden Chamber assay, which quantifies the number of invading cells atop a cell-permeable polyester porous membrane over a larger well containing osteogenic media, was also used to examine the response to 1,25(OH)2D. MG63 cells remained on the surface of the membrane regardless of treatment conditions, confirming their low metastatic potential (Figure 2F). In contrast, metastatic LM7 cells invaded the membrane after 24 hours, with a statistically significant reduction in the number of invading cells after 1,25(OH)2D treatment compared to vehicle control (Figure 2G). The results indicate that 1,25(OH)2D inhibits EMT in both OS cell models to varying degrees.
Figure 2 1,25(OH)2D inhibits osteosarcoma cell migration and invasion. (A) MG63 cells were subjected to scratch assays for 24-72 hours in the presence of vehicle or 10nM 1,25(OH)2D. ImageJ thresholding was employed to compute surface areas. The red asterisks indicate the region of increased magnification and crystal violet staining depicted in (B). (B) Staining with crystal violet of migratory MG63 cells. (C) Analysis of the MG63 scratch assay. The graph illustrates n=4 experimental setups for each condition, along with the average scratch area. Two-way ANOVA Multiple comparisons using Tukey’s test; p ≤ *0.05 and ****0.0001. (D) Metastatic LM7 cells were subjected to scratch studies for 24-72 hours with vehicle or 10nM 1,25(OH)2D. (E) Analysis of the LM7 scratch assay. The graph illustrates n=4 experimental setups for each condition, with the average scratch area indicated. Two-way ANOVA Tukey’s multiple comparison test; p ≤ ****0.0001. (F) Boyden chamber assay performed using MG63 cells. Right, overview of assay setup. Below is a sectional view of the membrane. (G) Boyden chamber assay using metastatic LM7 cells after 24 hours. Thresholding performed with ImageJ. Right: Quantitative analysis with Student’s t test; p ≤ *0.05 (n=3).
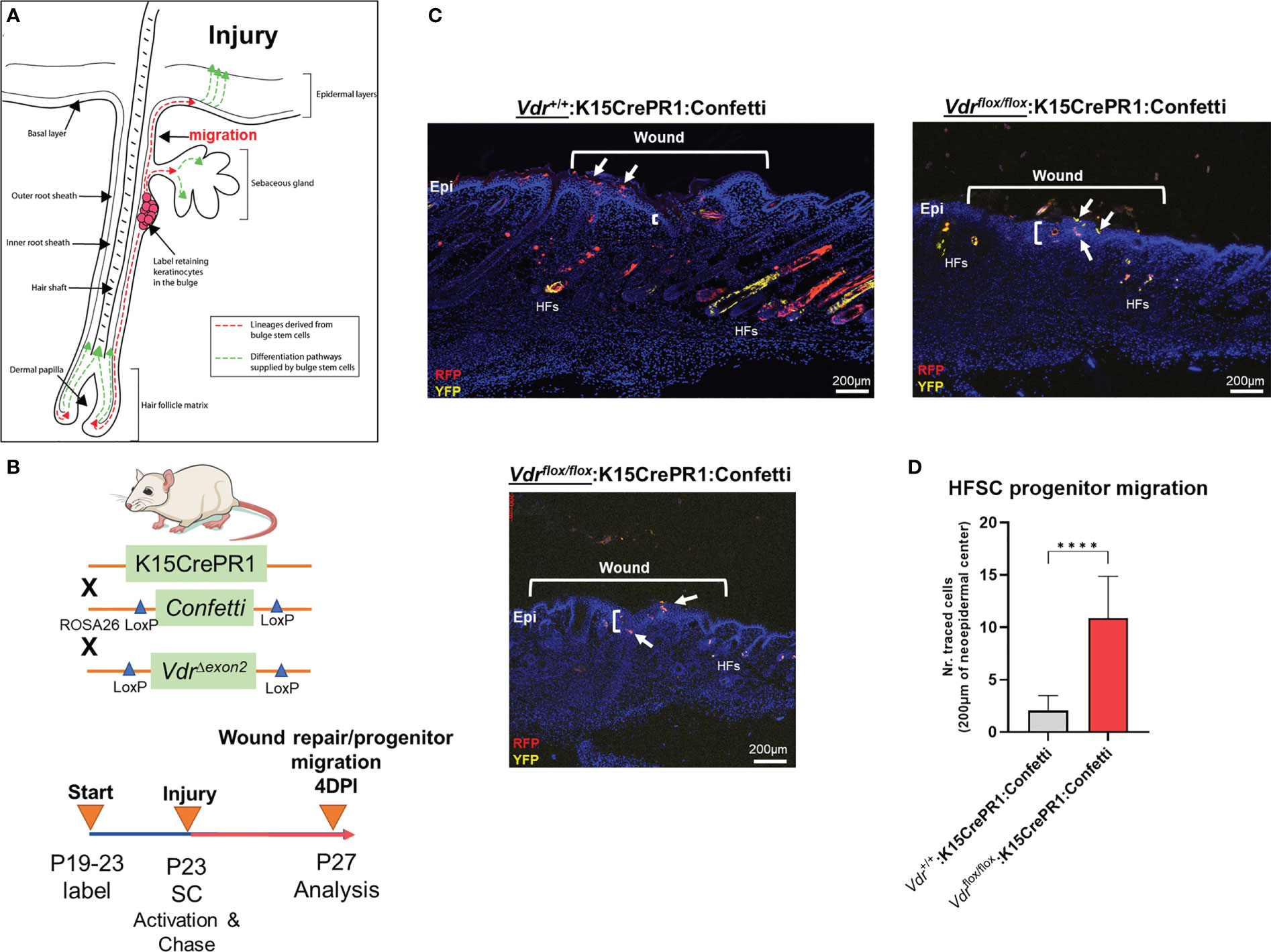
Experimentally induced EMT of mouse hair follicle stem cells reveals that Vdr signaling inhibits cell migration during normal cutaneous wound healingThe role of the VDR in OS migration prompted us to analyze whether migration is also affected in a different context such as skin wound repair. In addition to in vitro scratch assays of OS cells, we investigated cell migration of bulge stem cells in hair follicles, which is a well-established model whereby progenitor cells undergo EMT to repair wounds at distant regions (Figure 3A) (13). We used a lineage tracing approach to quantify the number of bulge stem cell progenitors that reside in the wound epicenter over time as a proxy for migration. To carry out these experiments, the RU486 inducible keratin 15 (K15)-crePR1:Vdrflox/flox:Confetti reporter mouse line was created, and lineage traced for four days after wounding to capture the early stage of the process (Figure 3B). Ablation of the Vdr within bulge stem cells increased the number of Confetti-labeled progenitors localized to the epicenter of wounds but not in individual hair follicles, resulting in enhanced wound closure (Figure 3C, upper left and lower panels; Figure 3D). Furthermore, bulge stem cell specific Vdr ablation resulted in thicker wound neoepidermis, an indication of increased EMT and fibrosis occurring following Vdr ablation. Mice with functional Vdr within bulge stem cells, on the other hand, had contiguous clonal streaks of Confetti-positive cells within individual hair follicles, indicating commitment of bulge stem cell progenitors down the hair follicle lineage (Figure 3C, upper left panel). Furthermore, a subset of bulge stem cell progenitors was fate restricted to the epidermal lineage but only migrated to the wound periphery four days after injury. These findings suggest that the Vdr regulates the fate of bulge stem cells and inhibits the migration of progenitor cells in healthy animals, which are consistent with its role in blocking migration of cancer cells.
Figure 3 Vdr signaling inhibits cell migration in vivo. (A) Hair follicles include oligopotent stem cells that, in response to environmental stimuli such as injury, can create hair follicle and/or epithelial progenitor cells. These cells can move to their final destinations to accomplish specific functions, such as fibrosis-based wound healing and hair follicle development. (B) Top: In order to explore the involvement of the Vdr in injury-induced cell migration in vivo, a conditional Vdr ablation and hair stem cell Confetti reporter mice was developed. Bottom: Male experimental mice were treated topically with RU486 between the post-natal (P) ages of 19 and 23 days to activate Cre recombinase and label the initial pool of keratin 15 (K15)-positive hair follicle stem cells. Following 3-mm full-thickness skin lesions to activate stem cells (SCs), the animals were tracked for four days to assess the lineage and migration of progenitor cells (YFP, yellow fluorescent protein; RFP, red fluorescent protein). (C) Left: At 4 days post-injury (DPI), the control animals’ wounds had not fully healed, as evidenced by the thin neo-epidermis, the neo-epidermal gap, and the migration of tagged progenitor cells to the wound margin. Right and Bottom: Experimental animals with Vdr ablation exhibit quicker wound closure, increased thickness (fibrosis) of the neo-epidermis, and the presence of tagged progenitor cells in the center of the neo-epidermis, indicating accelerated migration and improved tissue healing. In addition, Vdr deletion in hair stem cells hindered the development and contribution of progenitor cells to individual hair follicles, demonstrating that the Vdr plays an essential role in hair formation. (D) Quantification of hair follicle stem cell progenitors in the wound’s epicenter. Student t test; p ≤ ***0.001 (n=4-5).
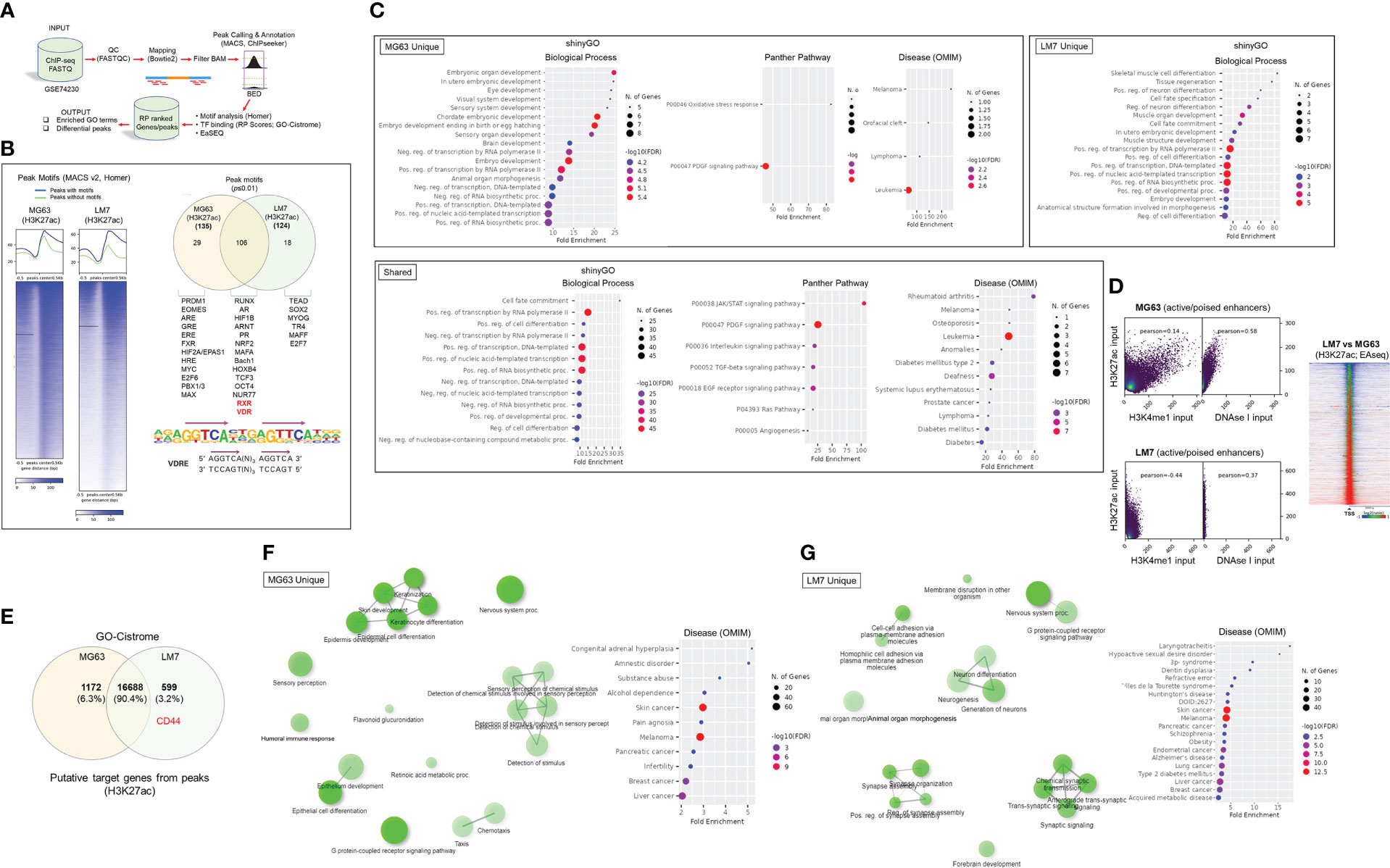
Epigenomic and putative target gene and activity analysis reveals the integration of the VDR with neurotransmitter pathways in osteosarcoma cell modelsGiven the distinct features of the OS subtypes, we sought to better define molecular and epigenetic regulators within the models. We re-analyzed epigenomic data obtained from MG63 and LM7 cells, as well as data obtained from primary patient OS samples and those that had spread to lung tissues (GSE74230), by focusing on transcription factor (TF) motif binding, differential peaks, and gene ontology (GO) analysis of acetylated (ac) H3K27 histone modifications that reflect active transcription (Figure 4A) (64). TF factor motif analysis within H3K27ac peaks revealed 29 MG63-specific TF binding sites, including those for EOMES, PRDM1, EPAS1 (HIF2A), E2F6, and MYC/MAX, which reflect known early development, tumor-initiation, and stemness factors (Figure 4B, Worksheet 1) (65). GO analysis of motif-binding TFs confirmed their role in embryonic development, including brain development, in part due to the function of EOMES (also known as T box transcription factor 2, TBR2) in stem cell mediated neurogenesis (Figure 4C) (66). In addition, pathway analysis revealed the involvement of oxidative stress responses, a known regulator of cell migration (67), and the PDGF signaling pathway, which integrates major signaling hubs to promote stem cell self-renewal (68). Eighteen TF binding sites specific to LM7 were identified for TEAD, TR4, MYOG, SOX2, and E2F7, which support mesenchymal gene expression to disrupt ECM and cell-cell adhesions to promote cell migration (Figure 4B) (69–71). Of note, fibroblasts can transform into skeletal muscle cells via myogenin (MYOG) (45), indicating a potential regulatory link between metastatic OS and other musculoskeletal cancers such as rhabdomyosarcoma (72), as revealed by the GO analysis (Figure 4C). RUNX binding was observed in both OS cell lines, which is particularly important in OS development in its aberrant transcriptional control of osteoblast commitment (73). Importantly, H3K27ac modifications were linked to VDR and RXR heterodimeric vitamin D response element (VDRE) binding in both cell models, indicating that 1,25(OH)2D regulates multiple functions of OS subtypes (Figure 4B). Interestingly, GO Disease analysis revealed that subtype-specific TFs were strictly associated with cancers, whereas shared TFs were associated with a variety of cancer types as well as diseases such as type 2 diabetes mellitus, a risk factor for cancer development. In addition, by comparing H3K27ac with H3K4me1 (i.e., histone changes associated with enhancers) modifications and DNase I hypersensitive sites, the degree of active versus poised enhancer utilization within the OS subtypes was determined at the epigenome-wide level. In MG63 cells, there was a weaker correlation between H3K27ac and H3K4me1 than in LM7 cells (0.14 versus 0.44), indicating that enhancer elements are poised to help define a more undifferentiated cell state (Figure 4D left) (74). This was also shown by comparing overall H3K27ac marks between LM7 and MG63 cells, showing more gene activity in LM7 cells (Figure 4D right).
Figure 4 Epigenomic and putative target gene analysis of osteosarcoma cell models. (A) Framework for ChIP-seq analysis and peak calling across MG63 and LM7 OS cell lines derived from GSE74230. Data quality specifications for FASTQ files processed with the Peak calling and Annotation pipeline. Using enriched GO keywords based on differential peaks, biological process, route, and disease functional annotations were generated. Using Homer and GO-Cistrome, motif analysis and transcription factor regulatory potential (RP) scores were obtained. Utilizing EaSeq, heatmaps and differential peaks were generated. (B) Left: Homer was utilized to identify histone modification peaks (H3K27ac) and motif discovery between LM7 and MG63 cells. In HOMER, Summits.bed files from MACS2 (Model-based analysis of ChIP-Seq) were utilized to identify recognized motifs. The x-axis analysis utilized fixed-size peak discovery on histone markers. Right: A subset of peak motifs that are enriched in the LM7 and MG63 cell lines. Motifs having a p-value 0.01 are displayed. Below: Sequence of the consensus VDR direct repat 3 response element (VDRE) is shown. (C) Gene ontology enrichment analysis of OS motifs with ShinyGo version 0.75. The x- axis represents fold enrichment, which indicates the extent to which genes of a certain pathway/biological process/disease are overrepresented. Fold enrichment is defined as the proportion of genes in the list that belong to a pathway divided by the proportion of genes in the background. The hues show the magnitude of the negative log10 of the false discovery rate (FDR), which indicates the likelihood that the enrichment occurred by coincidence (larger value means smaller FDR). (D) Assessing epigenome-wide poised or active enhancers between MG63 and LM7 cells. Right: BED files were used to perform Pearson correlation analysis between H3K27ac/H3K4me1/DNase I peaks. Left: Heat map showing comparison of H3K27ac peaks between cell lines. (E) Probable target genes generated from H3K27ac peaks identified using GO-Cistrome. (F) Gene ontology enrichment analysis of MG63 OS H3K27ac mapped genes using ShinyGo version 0.75, with network analysis shown in green. (G) Gene ontology enrichment analysis of identified LM7 OS H3K27ac genes utilizing ShinyGo version 0.75 Green depiction of network analysis.
Next, using GO-Cistrome, candidate target genes mediated by H3K27ac were identified and assessed (Figure 4E) (75). Approximately 13.3% and 9.3% of the H3K27ac peaks in LM7 and MG63 were located within 1KB of the promoter regions of genes (Figure S2). There were 1172 and 599 H3K27ac-defining MG63 and LM7 specific gene signatures, with CD44 identified as an active LM7 target gene (Worksheets 2, 3). The GO Network and Disease analysis of MG63 putative genes was consistent with the motif analysis, which found correlations with epidermal differentiation, neuronal/sensory systems, and invasive malignancies (Figures 4F, S3, File S1). Compared to MG63 cells, LM7 H3K27ac-linked genes were significantly associated with neurogenesis and neurological disorders, as well as cell-cell adhesion molecules (Figures 4G, S3, File S2). Given the likelihood that osteosarcomas may act as neuroendocrine carcinoids, we examined transcription of ionotropic and metabotropic glutamate receptors following 1,25(OH)2D treatment of MG63 cells. Only three of the available 24 mammalian receptors and subunits were expressed in MG63 cells (i.e., GIRK3, GRIN3A, and GRIN2D), with two of the three receptors downregulated by 1,25(OH)2D (Figure S4) (GSE220948). The ionotropic glutamate [kainate] receptor 3 (GIRK3) was downregulated at both time points following 1,25(OH)2D treatment, whereas the ionotropic glutamate [NMDA] receptor subunit epsilon-4 (GRIN2D) was only downregulated after 48 hours and was identified as a putative H3K27ac-target gene in LM7 cells (Worksheet 3). These results imply that vitamin D3 may decreased tumor burden by inhibiting OS-nerve interactions.
Vitamin D modulates MG63 tumorigenicity by inhibiting nonsense-mediated RNA decay in a SMG6-SMG7-dependent mannerTo gain insights into the biological aspects of epigenetic modifications that are differentially represented in the OS subtypes, we utilized EaSeq (Figure 5A). There were approximately 2,634 more H3K27ac gene-associated peaks than H3K4me1 peaks due to the fine-tuning regulatory effects of enhancers (Worksheets 4, 5). By comparing 1,25(OH)2D resistant LM7 to MG63 cells, overrepresented target genes of H3K27 acetylation implicated the involvement of the mRNA surveillance pathway, but underrepresented genes engaged the HIF-1 and TNF signaling pathways, indicating a putative decrease in immunogenicity and immunosurveillance of LM7 metastatic cells (Figure 5B). LM7 cells overexpressed NMD machinery components involved in the assembly of the surveillance complex at aberrant PTCs (e.g., eRF1, UPF2, UPF3) and cleavage of RNA templates involving exo/endonucleolytic pathways (SMG5, SMG6, SMG7) (Figure 5C). Interestingly, the RNA-binding protein MUSASHI (encoded by MSI1), which lies upstream of the surveillance complex, was increased in LM7 cells indicating the regulation of translation of defective transcripts vulnerable to nonsense-mediated translation repression (NMTR) (76).
Figure 5 Differential peak analysis of osteosarcoma lines reveals overrepresentation of the NMD pathway. (A) Predicting distinct H3K27ac and H3K4me1 peaks in OS cells using the EaSeq software. Greater differential peaks between cell lines and histone markers based on Log2 fold variations inside designated regions relative to the transcription start site (TSS). (B) Gene ontology enrichment analysis of LM7 OS H3K27ac overrepresented and underrepresented putative target genes with ShinyGo version 0.75. The mRNA surveillance pathway is highlighted among shown biological processes. (C) Pathview visualization of the nonsense-mediated RNA decay (NMD) pathway in relation to the H3K27ac target genes affected in LM7 cells. Targets that were overrepresented at the exon junction and surveillance and cleavage complexes are shown in red. (D) RNA-Seq evaluation of nonsense-mediated translation suppression and NMD machinery genes in MG63 cells. Notable was SMG6, which is responsible for the cleavage of pre-mRNA, resulting in its degradation. (E) In MG63 OS cells, 1,25(OH)2D selectively upregulates known NMD target genes. MG63 cells treated with 10nM 1,25(OH)2D for 48 hours exhibited an elevated 3:1 ratio of upregulated known NMD target genes relative to non-NMD target genes. The RNA-seq data set’s statistically significant genes (FDR ≤ 0.05) were compared. The symmetry of the data distribution was also characterized by descriptive statistics (skewness/kurtosis). (F) In MG63 OS cells, 1,25(OH)2D does not selectively upregulate non-NMD target genes. After treatment with 1,25(OH)2D, the ratio of upregulated to downregulated genes in a random sample of non-NMD target genes is 1:1. (G) Analysis of the receiver operating characteristic (ROC) curve for statistical comparisons and identification of NMD or non-NMD target gene classes. To compare the two classifiers, the area under the ROC curve (AUC) was computed, which quantifies the overall performance of the test to discriminate between genes in the presence of 1,25(OH)2D that are NMD-target or non-NMD target genes relative to the null hypothesis (0.5). (H) Phosphorylated UPF1S1127 expression in MG63 cells after 24 hours of 10nM 1,25(OH)2D. Bar = 20µm. (I) NDM inhibitor 14 (NMDI-14) treatment of MG63 for 24 hours. Two-way ANOVA test with Tukey’s multiple comparisons (n=3); ***p ≤ 0.001, ****p ≤ 0.0001. (J) DsiRNA SMG6 knockdown of MG63 cells for 24 hours. Negative control (NC) duplexes used for comparisons. All conditions were compared to transfection reagent (BioT) only treatments. Two-way ANOVA test with Tukey’s multiple comparisons (n=3); **p ≤ 0.01, ****p ≤ 0.0001. NS, not significant.
We next investigated vitamin D3 regulation of NMD in MG63 and LM7 cells. We observed a statistically significant downregulation of autoregulatory NMD machinery as well as NMTR genes following 10nM 1,25(OH)2D treatment of MG63 cells (Figure 5D) (77). Interestingly, there was a statistically significant downregulation of SMG5 among a subset of detectable NMD genes in LM7 cells treated with 10nM 1,25(OH)2D for 48 hours as well (Figure S5A). Next, we examined the impact of 1,25(OH)2D on known NMD target genes. On the basis of four investigations in which NMD effectors were inhibited in human cervical cancer cells (77–80), we compiled a list of NMD-target and non-NMD-target genes (Figure S6). Based on this analysis, more than 4,000 genes were identified as NMD nontarget genes, whereas more than 900 genes were recognized as NMD targets by at least one study; whereby, 50 genes were classified as NMD core targets by all studies. These curated NMD-target and nontarget genes were used to analyze the RNA-seq dataset generated from MG63 cells. Thirty nine of the 50 core NMD-target genes were mapped to MG63 cells, and overexpressed in a 3:1 ratio (-0.6 skewness and 2.2 kurtosis) relative to those that were downregulated in response to 1,25(OH)2D treatment (Figure 5E). TSC22 Domain Family Member 3 (TSC22D3), also known as GILZ, which encodes a glucocorticoid-inducible anti-proliferation transcription factor, was the most elevated NMD target in MG63 OS cells (81). Stanniocalcin-2 (STC2), which is involved in normal osteoblast differentiation (82), was the second most elevated NMD-target gene in response to 1,25(OH)2D, as well as CLDN15 and BTN2A1, which may promote cell-to-cell adhesion to limit migration (83) and the expression of a phosphoantigen to enhance T cell activity (84), respectively. In contrast, a multiple-comparison study of NMD nontarget genes demonstrated a 1:1 (-0.3 skewness and 0.4 kurtosis) association between overexpressed and downregulated genes (Figure 5F). In addition to the descriptive statistics, a receiver operating characteristic (ROC) curve analysis was performed to investigate the capacity to distinguish between genes that are one of two classifiers (i.e., NMD target or non-target genes) in the presence of 1,25(OH)2D relative to the null hypothesis (area under the curve, 0.5). The C statistics revealed statistical significance (p = 0.03) between the two classifiers based on ROC curve analysis (Figure 5G).
To gain a better understanding of how 1,25(OH)2D controls the NMD pathway in MG63 cells, we investigated UPF1, which is a key RNA helicase that assembles the upstream surveillance complex at the exon junction complex of aberrant mRNAs dictated by its phosphorylation status (Figure 5C) (85, 86). After 24 hours of 1,25(OH)2D treatment, there was no significant change in UPF phosphorylation compared to vehicle treated MG63 cells (Figure 5H), suggesting that changes in UPF2/UPF3 expression may be the key NMD upstream factors regulated by 1,25(OH)2D (Figure 5D). To better understand the role of downstream NMD pathway factors on regulation of NMD-target (e.g., GILZ, STC2) and non-target (e.g., FKBP5) genes in MG63 cells, we pharmacologically and genetically ablated components of the RNA degradation complex. First, we utilized the small molecule NMD inhibitor, NDMI-14, which prevents SMG7-UPF1 interactions (87), and observed a modest increase in GILZ after 24 hours of treatment (Figure 5I). There was a small, yet significant increase in FKBP5, which may reflect stress responses of high NMDI-14 concentrations, given its function as a Hsp90-associated co-chaperone (88). Next, we utilized dicer-substrate short interfering RNAs (DsiRNA) to knockdown (KD) SMG6 in MG63 cells and observed a modest increase in STC2, as well as a statistically significant, yet small increase in GILZ (Figures 5J, S4B). Among the conditions tested, the findings suggest that GILZ is dependent on both SMG7 and SMG6, while STC2 is modulated by SMG6 only, and that 1,25(OH)2D exhibited a greater impact on NMD target genes likely reflecting its simultaneous effects on multiple upstream and downstream components of the NMD pathway to suppress OS.
LM7 resistance to 1,25(OH)2D is dictated by high SNAI2 and low VDR levelsTo better understand the functional role of 1,25(OH)2D on metastasis, the transcriptional responses of EMT genes on highly metastatic LM7 cells were compared to those of MG63 cells. The decrease in expression of SNAI2 in MG63 cells was less pronounced in LM7 cells following 1,25(OH)2D treatment, while CD44 expression was unaffected (Figure 6A). There was a concentration-dependent effect to 1,25(OH)2D on MMP3 expression 24 hours after treatment, with 10nM increasing MMP3 levels, suggestive of resistance. In addition, the dose and duration of 1,25(OH)2D in LM7 cells resulted in a maximal 2-fold overexpression of SOD2, a potent antioxidative enzyme that inhibits tumor progression via reduced free radical production, which was less pronounced compared to MG63 cells (Figure 6B). Interestingly, there was significant overexpression of the mitochondrial monooxygenase, CYP24A1, which catabolizes and deactivates 1,25(OH)2D via 24-hydroxlation, in LM7 cells after 1,25(OH)2D treatment (Figure 6B). Also, by evaluating the relative levels of VDR and SNAI2 transcripts, MG63 cells were characterized as VDRhighSNAI2low whereas LM7 cells were VDRlowSNAI2high, providing more evidence for 1,25(OH)2D resistance in LM7 cells (Figure 6C). By increasing the duration of 1,25(OH)2D from 48 to 96 hours, LM7 cells overcame the resistance as depicted by a significant decrease in CD44 protein expression levels (Figure 6D).
Figure 6 LM7 metastatic cells are resistant to 1,25(OH)2D and SNAI2 is a VDR-target gene. (A) qPCR study of EMT, vitamin D, and anti-oxidative transcripts in LM7 cells following treatment with 1,25(OH)2D. Two-way ANOVA Multiple comparisons using Tukey’s test; p ≤ **0.01, ***0.001 (n=3). (B) qPCR study of vitamin D and anti-oxidative transcripts in MG63 cells following treatment with 1,25(OH)2D. Two-way ANOVA Tukey’s test for multiple comparisons; p ≤ 0.001 (n=3). (C) Normalized VDR and SNAI2 gene expression in LM7 and MG63 cells. Two-way ANOVA Test for multiple comparisons using Tukey’s method; p ≤ **0.01, ***0.001, ****0.0001 (n=3). (D) 96 hours of 10nM 1,25(OH)2D treatment decreases CD44 expression levels in LM7 cells. Arrows depict cells with largest differences in CD44 expression. Unpaired T test (n=4). (E) ChIP study of VDR-RNApol2 chromatin interactions in 1,25(OH)2D-treated MG63 cells. A two-way ANOVA was performed on the fold-change between stimulated and baseline states. Dunnett’s multiple comparison test relative to IGX1A; p ≤ ****0.0001 (n=3). (F) Analysis of transcription factors that regulate SNAI2 using the Cistrome DB Toolkit. The static plot with RP scores on the X- and Y-axes depicts transcription factors, whereas the blue dots represent individual public ChIP-seq data sets. Listed next to each transcription factor are public GEO data series. NS, not significant.
VDR-mediated suppression of the EMT transcriptional repressor SNAI2 occurs via chromatin condensationBased on the ATACseq and qPCR data we hypothesized that SNAI2 is a direct VDR target gene. Using NUBIscan (89), a putative VDRE direct repeat (DR3) binding site was found within 1KB upstream of the transcriptional start site (TSS) exclusively in the SNAI2 locus, but not in the SOD2 locus (Figure S7). MG63 cells treated with 1,25(OH)2D or vehicle were subjected to conventional chromatin immunoprecipitation quantitative PCR (ChIP-qPCR) to measure VDR binding and transcriptional activity mediated by RNAPolII at the SNAI2 distal and proximal promoter regions (Figure 6E). After 1,25(OH)2D treatment, there was decreased enrichment of the VDR and RNAPolII in the proximal promoter region of SNAI2 compared to the intergenic control (IGX1A). After 1,25(OH)2D treatment, SOD2 transcriptional activity increased, although this was not VDR-dependent. Given the likelihood that VDR regulates SNAI2 transcriptional activity directly, the Cistrome DB toolset was used to identify additional VDR : SNAI2 chromatin interactions (90). Three GEO datasets were identified when the 1KB domain of the SNAI2 TSS was investigated for potential VDR binding in lymphocytes, hepatocytes, and prostate cancer cells (Figure 6F). The identification of putative SNAI2 binding sites in keratinocytes, which are highly migratory cells of the body (Figure 6F) (67), also suggests the possibility of intracrine feedback effects that maintain high SNAI2 levels in LM7 cells. Overall, LM7 cells are more resistant to 1,25(OH)2D than MG63 cells, which may be a direct result of CYP24A1 catabolic effects on ligand and decrease VDR responses at the SNAI2 locus.
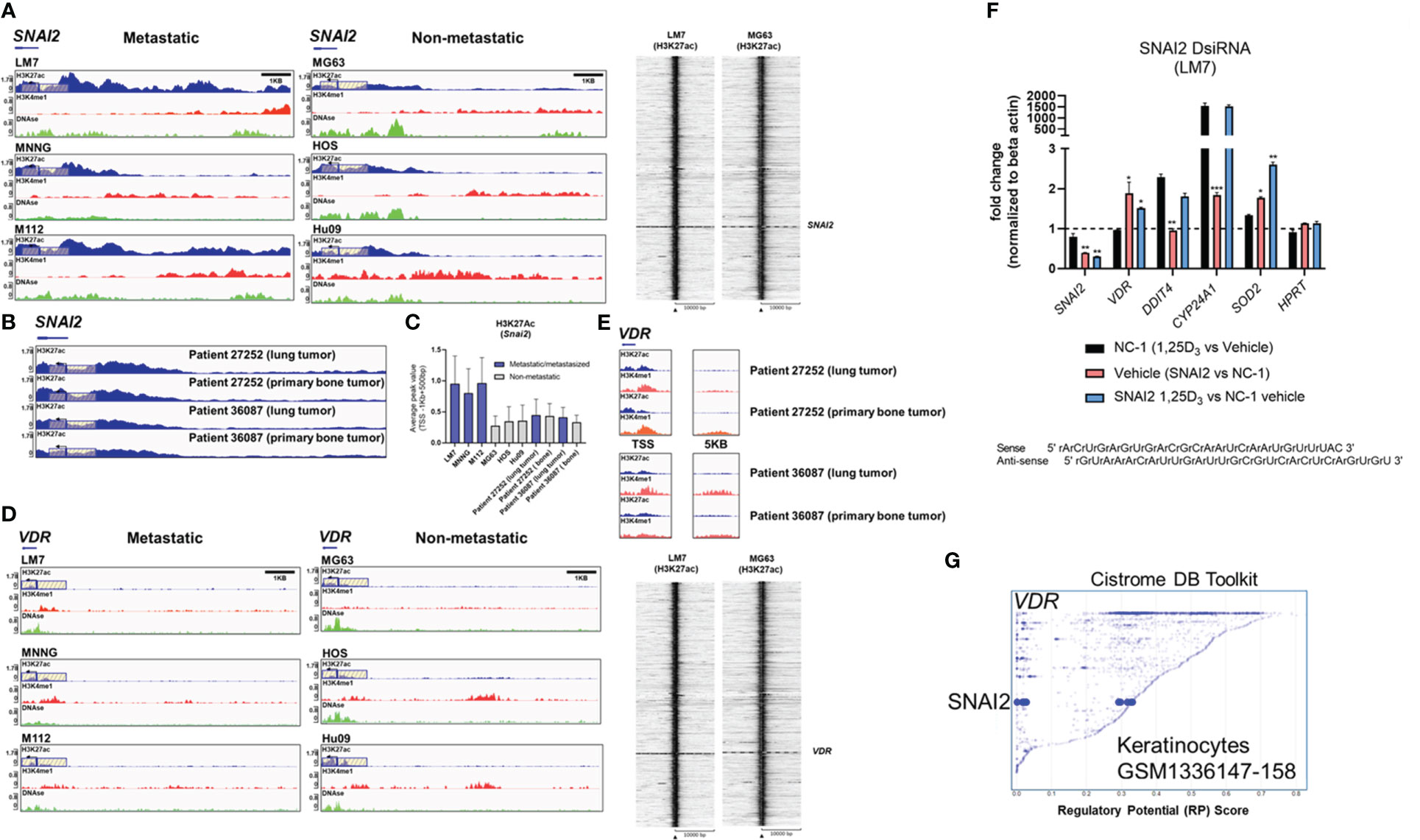
H3K27 is differentially acetylated at the SNAI2 and VDR loci in highly metastatic and low metastatic osteosarcomasTo gain a better understanding of SNAI2 and VDR regulation in osteosarcomas, we evaluated H3K27ac and H3K4me1 epigenetic alterations in a panel of highly metastatic and low metastatic human OS cell lines (GSE74230) (64). Compared to low metastatic cell lines MG63, HOS, and Hu09, the highly metastatic OS cell lines LM7, MNNG, and M112 had higher average H3K27ac peak values at the SNAI2 TSS (Figures 7A, C), which corresponded to the degree of DNase hypersensitivity (lower tracks). Intriguingly, there were no differences in SNAI2 H3K27ac peak values when comparing two OS patient primary bone and metastasized lung tumor samples (Figures 7B, C), possibly due to reduced migration in the lung tumor microenvironments compared to in vitro conditions of cell lines. In contrast, the low metastatic cell lines revealed greater H3K27ac peaks at the VDR TSS than the highly metastatic cell lines (Figure 7D). Similarly, there were no differences in H3K27ac and H3K4me1 epigenetic modifications at the VDR TSS and distal 5kb regions between OS patient primary bone or lung tumors (Figure 7E). These findings show that metastasizing OS cells exhibit increased SNAI2 H3K27ac, but low metastatic lines exhibit increased VDR H3K27ac, potentially making them more responsive to 1,25(OH)2D, a pattern that was abolished in metastasized patient lung tumor samples.
Figure 7 Differential H3K27 acetylation at the SNAI2 and VDR loci between OS subtypes and mediation of antioxidative pathways by vitamin D and SNAI2. (A) ChIP-seq studies and peak calling for the SNAI2 gene in metastatic and non-metastatic OS lines from GSE74230. Evaluation of H3K27ac, H3K4me1, and DNase I hypersensitive sites 500bp downstream and 1000bp upstream of the transcription start site (TSS). Quantification of peak values in (H) Right: Using EaSeq, a heat map of H3K27ac was created at the TSS. Using IGV’s auto-scale of BED files, all scales reflect the minimum and maximum reads in that window. (B) ChIP-seq study of H3K27ac changes at the SNAI2 gene in primary bone and lung tumor tissues from OS patients. Samples generated from GSE74230. (C) Quantification of the average peak values of OS cell lines that are poorly metastatic, metastatic, and patient samples. (D) ChIP-seq studies and peak calling for the VDR gene in metastatic and non-metastatic OS lines from GSE74230. Evaluation of H3K27ac, H3K4me1, and DNase I hypersensitive sites 500bp downstream and 1000bp upstream of the transcription start site (TSS). Using IGV’s auto-scale of BED files, all scales reflect the minimum and maximum reads in that window. Using EaSeq, a heat map of H3K27ac was created at the TSS. (E) ChIP-seq study of H3K27ac changes at the VDR gene in primary bone and lung tumor tissues from OS patients. Derived samples from GSE74230. (F) Gene responses to DsiRNA-mediated SNAI2 knockdown in LM7 cells. The sense and antisense DsiRNA SNAI2 sequences are displayed below. Tukey’s multiple comparison test for two-way ANOVA; p ≤ *0.05, **0.01, ***0.001 (n=4). (G) Cistrome DB Toolkit study of VDR-regulating transcription factors such as SNAI2. The static plot with RP scores on the X- and Y-axes depicts transcription factors, whereas the blue dots represent individual public ChIP-seq data sets. Listed next to each transcription factor are the public GEO data series.
DsiRNA knockdown of SNAI2 reveals antioxidative responses mediated by vitamin D3To better comprehend the plausibility of VDR-SNAI2 reciprocal regulation, we utilized DsiRNA to KD SNAI2 in LM7 cells (Figures S8, S9). In order to explore the transcriptional consequences of DsiRNA KD of SNAI2 in LM7 cells, a comparative qPCR analysis was conducted. Negative control (NC-1) DsiRNA treated samples exhibited the predicted responses to 1,25(OH)2D, notably the downregulation of SNAI2 and the overexpression of DDIT4 (a potent VDR-dependent inhibitor of mTOR (51)), CYP24A1, and SOD2 (Figure 7F, black bars). Comparing vehicle treatment across SNAI2 and NC-1 DsiRNA KD samples, the expected >60% KD of SNAI2 and an intriguing elevation of VDR, CYP24A1 and SOD2 were detected, indicating direct dependence and negative regulation by SNAI2 (Figure 7F, red bar). To provide additional evidence of potential SNAI2 interactions at the VDR promoter site, we re-evaluated the Cistrome DB Toolkit and identified the datasets GSM1336147-158, which illustrate putative interactions in highly motile skin keratinocytes (Figure 7G) (91). In contrast, SNAI2 DsiRNA KD had no effect on DDIT4, suggesting SNAI2-independent signaling. The effect of SNAI2 DsiRNA KD on 1,25(OH)2D responses revealed further SNAI2 downregulation, providing additional evidence that SNAI2 is VDR-dependent (Figure 7F, blue bar). Due to an absence of autocrine VDR signaling, there was no significant difference in VDR expression between vehicle (Figure 7F, pink bar) and 1,25(OH)2D SNAI2 DsiRNA KD (Figure 7F, blue bar) responses. After 1,25(OH)2D treatment, both DDIT4 and CYP24A1 levels returned to NC-1 levels when SNAI2 was inhibited, demonstrating once again that DDIT4 is independent of SNAI2, but CYP24A1 was found to be co-dependent on VDR and SNAI2. Intriguingly, given the maximal effect of most genes following SNAI2 DsiRNA KD and 1,25(OH)2D treatment relative to the control, SOD2 levels continued to increase (Figure 7F, blue bar), indicating a repressive effect of SNAI2 on VDR expression. Overall, SNAI2 can modulate 1,25(OH)2D responses, thereby linking EMT, ROS, and vitamin D3 signaling into a novel OS network (92, 93).
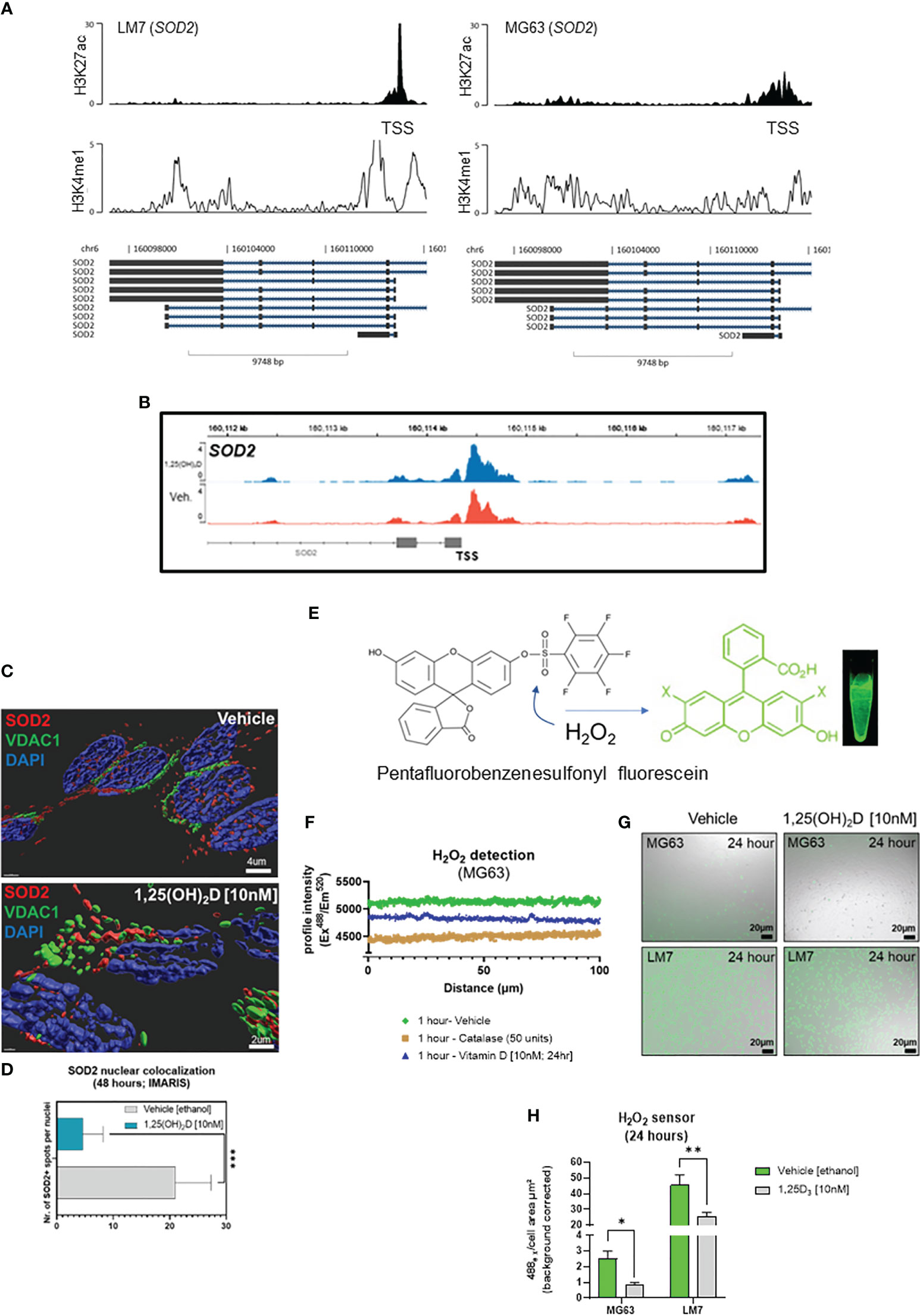
Vitamin D3 modulates SOD2 intracellular localization to inhibit osteosarcoma mitochondrial oxidative stressGiven that ROS regulates cell migration and vitamin D3 induces SOD2 expression (67), the epigenomic regulation of SOD2 in OS cell lines was evaluated. At rest, LM7 cells displayed increased H3K27 acetylation at the SOD2 TSS compared to MG63 cells, with augmentation of intronic and intergenic H3K4me1 modification (Figure 8A), indicating a pronounced antioxidative response mediated by active enhancer elements. ATACseq analysis indicated similar degree of chromatin accessibility at the SOD2 TSS in MG63 cells treated with 1,25(OH)2D or vehicle (Figure 8B), suggesting that SOD2 mRNA levels are likely regulated by post-transcriptional mechanisms (48). In addition, we analyzed SOD2 protein and observed that it was mainly located in the nucleus of MG63 cells under basal conditions and was seldom associated with VDAC1-positive mitochondria (Figure 8C). In contrast, 1,25(OH)2D directed the mitochondrial localization of SOD2, possibly allowing it to perform its primary ROS detoxifying functions (Figures 8C, D). Because of these OS subtype differences, we investigated ROS responses to 1,25(OH)2D treatment. Pentafluorobenzenesulfonyl fluorescein, a hydrogen peroxide (H2O2) reporter dye, revealed that 10nM 1,25(OH)2D for 24 hours promoted efficient scavenging of H2O2 in MG63 cells (Figures 8E, F) (67). Interestingly, in agreement with the epigenetics of SOD2, LM7 metastatic cells had an 18-fold increase in H2O2 compared to MG63 cells (Figure 8G), whereas 1,25(OH)2D was also able to suppress H2O2 levels, implying that anti-oxidative pathways regulated by vitamin D3 are shared in both OS subtypes (Figure 8H).
Figure 8 Vitamin D modulates nuclear-localized SOD2-to-SOX2 axis to antagonizes osteosarcoma oxidative stress, iron transport and pluripotency to dictate migratory status. (A) ChIP-seq study of the SOD2 region in LM7 and MG63 cells. (B) ATAC-seq study of MG63 cells at the SOD2 gene in the presence or absence of 1,25(OH)2D (10nM, 24 hours). (C) Confocal imaging of MG63 OS cells with SOD2 and mitochondrial VCAC1. Imaris surfaces for rendering 3D surfaces. (D) Quantitative analysis of nuclear SOD2. Student T test; p ≤ ***0.001 (n=5). (E) Schematic of the sensor pentafluorobenzenesulfonyl fluorescein for hydrogen peroxide (CAS 728912-45-6). (F) Intensity of the pentafluorobenzenesulfonyl fluorescein confocal profile over MG63 cells. Vehicle, catalase (to breakdown hydrogen peroxide), and 1,25(OH)2D were used to treat cells. (G) Representative confocal images demonstrating the formation of hydrogen peroxide by MG63 and LM7 cells under various circumstances. (H) Quantification of the presence of hydrogen peroxide in MG63 and LM7 cells. Tukey’s multiple comparison test, two-way ANOVA; p ≤ *0.05, **0.01 (n=4).
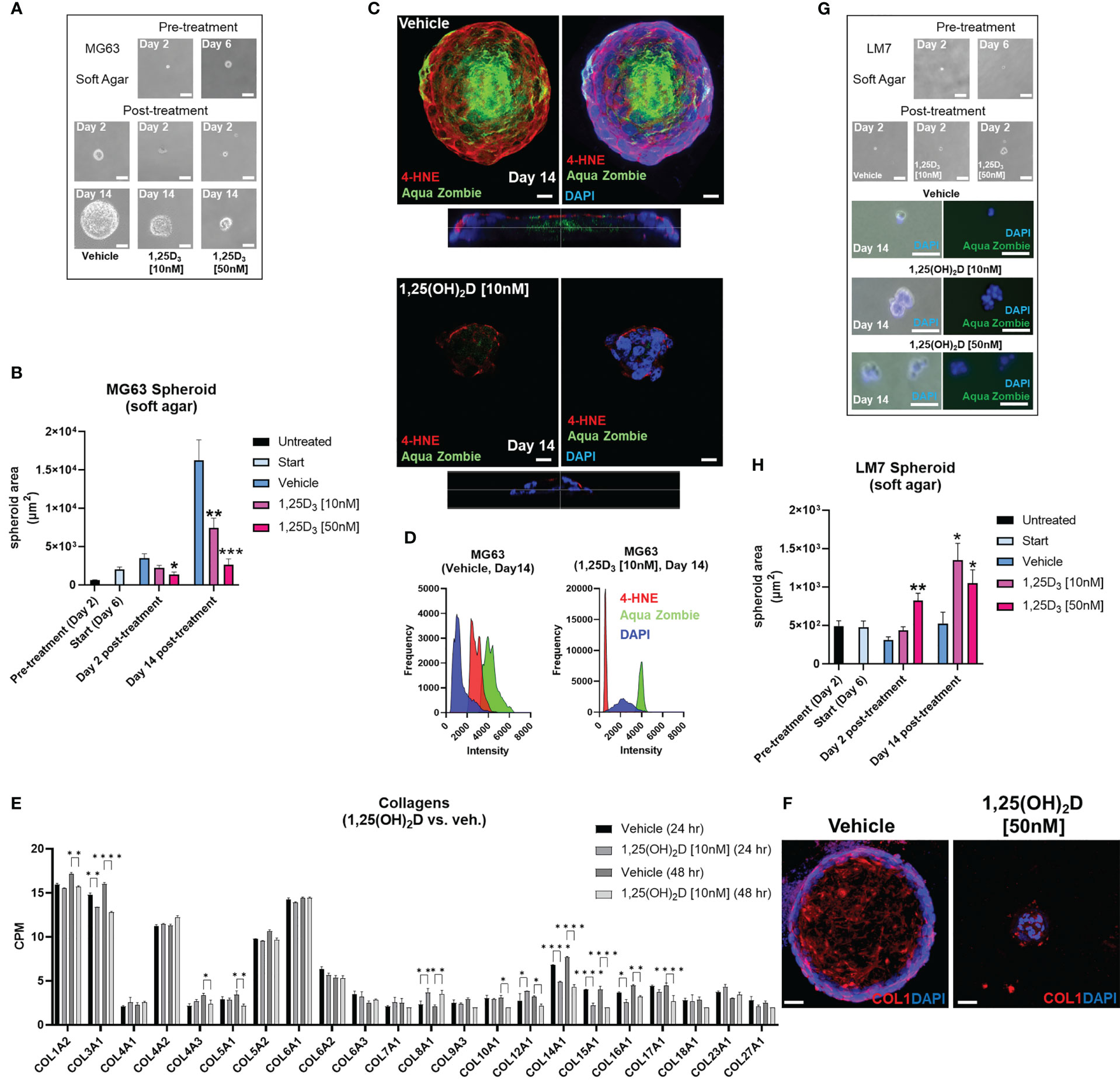
Vitamin D suppresses free radical generation and desmoplasia in low metastatic osteosarcoma and promotes MET of highly metastatic osteosarcoma in soft agar spheroidsNext, we used soft agar spheroids to determine if the antioxidative responses of 1,25(OH)2D are retained in the 3D environment. MG63 cells grew as round, well-circumscribed spheroids in soft agar for up to 14 days in culture (Figure 9A). When spheroids were treated with 10-50nM 1,25(OH)2D, there was a significant dose-dependent shrinkage (Figure 9B). Vehicle-treated MG63 spheroids exhibited a necrotic core (i.e. Zombie Aqua-positive), commonly seen in intact tumors, and surrounding cells with high 4-hydroxynonenal (4-HNE) expression, a marker for lipid peroxidation and free radicals (Figure 9C, bottom orthogonal views). In contrast, 1,25(OH)2D reduced the expression of 4-HNE in spheroids as well as cells expressing Zombie Aqua, indicating the presence of viable cells (Figure 9D). Given that vitamin D3 can elicit anti-fibrotic responses and that cancer is a fibrotic disease known to obstruct, in part, immune cell infiltration (12, 94), we examined collagen protein and mRNA expression in MG63 spheroids and cells. There are 28 types of human collagens encoded by 43 genes, and 22 collagen genes were expressed in MG63 cells. 1,25(OH)2D inhibited the transcription of half of the collagen types detected (11 out of 22), and suppressed mature type 1 collagen within spheroids (Figures 9E, F). Unlike MG63 cells, LM7 metastatic cells grown in soft agar did not form spheroids but instead remained as single viable cells with minimal growth after 14 days (Figure 9G). In contrast to MG63 cells, 1,25(OH)2D promoted the clustering of Zombie Aqua-negative LM7 cells in soft agar, implying the induction of mesenchyme-to-epithelial transition (MET) as a means of inhibiting metastasis (Figure 9H). Overall, the findings indicate that vitamin D3 inhibits free radical and collagen production in a cell intrinsic manner, and initiates cell-cell adhesion in a highly metastatic OS model.
Figure 9 Vitamin D reduces free radical production, desmoplasia, and lipid peroxidation in non-metastatic osteosarcoma spheroids, while promoting MET in metastatic osteosarcoma spheroids in soft agar. (A) MG63 cell spheroid assay with soft agar. Bar = 100µm. (B) Measurement of (A) Tukey’s multiple comparison test for two-way ANOVA; p ≤ *0.05, **0.01, ***0.001 (n=4). (C) Confocal imaging of MG63 spheroids with the lipid peroxidation indicator 4-HNE and the dead cell dye Zombie Aqua. Orthogonal perspective on the base of each composite. Bar = 20µm (D) Quantification of C by evaluating the frequency and intensity of every marker. (E) RNA-seq study of collagen expression before and after 1,25(OH)2D treatment in MG63 cells. Tukey’s multiple comparison test; p ≤ *0.05, **0.01, ****0.0001 (n=2). (F) Type 1 collagen protein expression in MG63 spheroids. Bar = 40µm (G) Spheroid assay with soft agar for LM7 cells. 2-day bar = 100µm; 14-day bar = 20µm (H) Measurement of (F) Tukey’s multiple comparison test, two-way ANOVA; p ≤ *0.05, **0.01 (n=4).
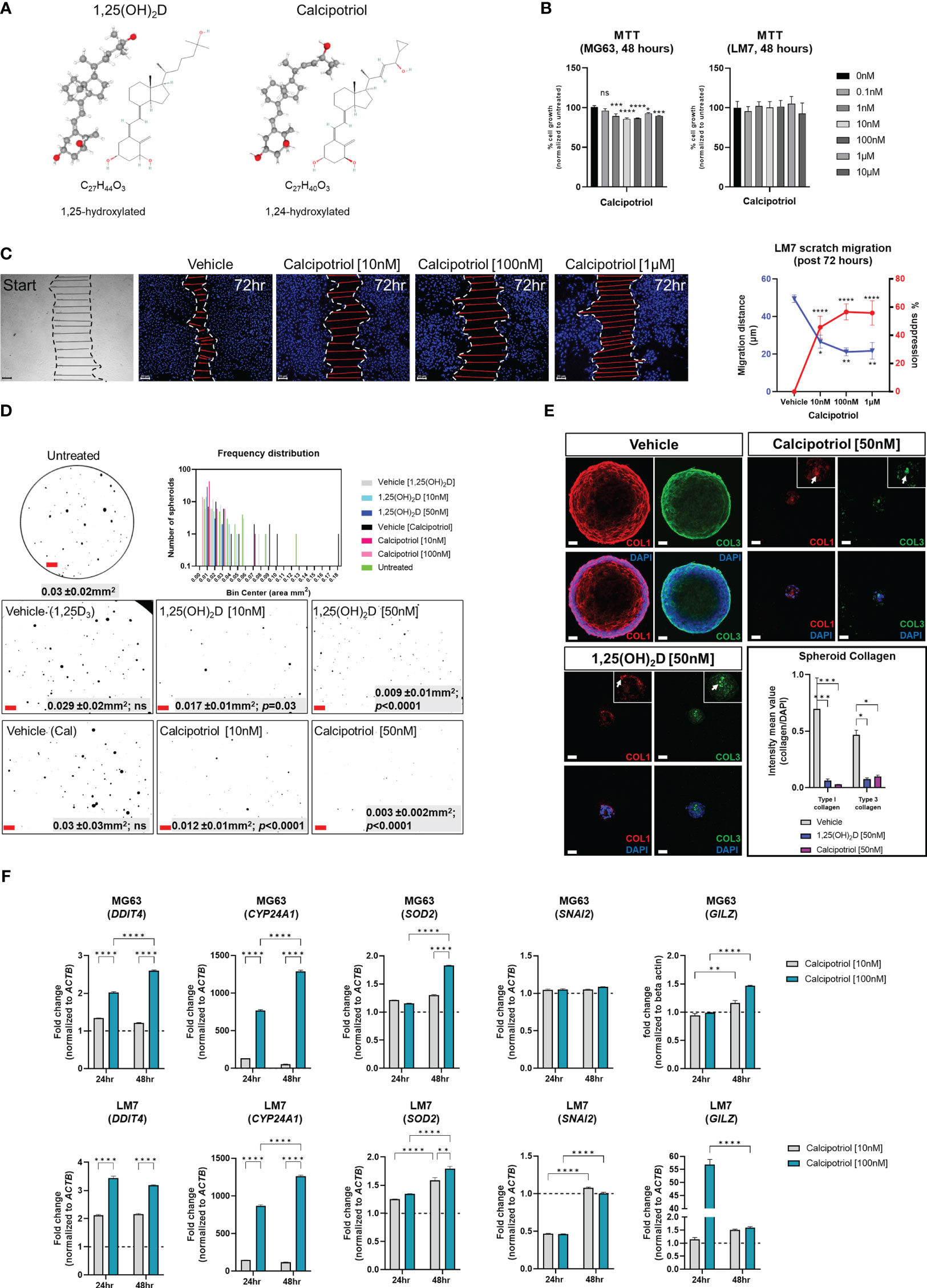
Calcipotriol, a clinically relevant vitamin D3 analogue, inhibits osteosarcoma migration, desmoplasia and growthBecause of its lower calcemic activity in regulating calcium metabolism for clinical application, we investigated the effects of calcipotriol, a synthetic analogue of vitamin D3 (Figure 10A) (58). The effects on cell proliferation were first assessed using the MTT assay, which revealed that calcipotriol inhibited MG63 cell proliferation starting at 1nM lasting 48 hours (Figure 10B). On the other hand, calcipotriol had no effect on LM7 proliferation during the treatment period. Instead, calcipotriol significantly inhibited LM7 migration compared to 1,25(OH)2D treatment (i.e., a ~50 vs 26% suppression; Figures 10C, 2E). Moreover, when compared to 1,25(OH)2D, calcipotriol significantly inhibited the growth of MG63 spheroids in soft agar (Figure 10D), with concomitant suppression of desmoplasia through decreased type 1 and 3 collagen production (Figure 10E). Interestingly, 1,25(OH)2D and calcipotriol both inhibited the laminar deposition of both types of fibrillar collagens, removing the outer surface of collagen “tracks” known to aid in EMT, metastasis, and the blocking of immune cells (Figure 10E, arrows) (95). The magnitude of calcipotriol effects on MG63 was not as pronounced as for 1,25(OH)2D, according to qPCR analysis (Figure 10F, upper). For example, after 48 hours, the SOD2 level induced by 1,25(OH)2D was close to 8-fold in MG63 cells (Figure 6B), but never exceeded 1.8-fold after calcipotriol treatment. Furthermore, calcipotriol treatment had no effect on SNAI2 in the study’s concentration range or duration when compared to 1,25(OH)2D treatment of MG63 cells (Figure 1B), reflecting its function as an unique VDR agonist. In contrast, qPCR analysis of LM7 cells revealed that the transcriptional readout was significantly enhanced after calcipotriol treatment compared to 1,25(OH)2D-treated cells (Figure 10F, lower). Calcipotriol, for example, reduced SNAI2 expression by half, a significant reduction compared to 1,25(OH)2D responses (Figure 6A). Similarly, calcipotriol stimulated a large transcriptional increase of GILZ 24 hours after treatment, indicating that the NMD pathway may have dynamic and rapid effects on aberrant gene expression. Overall, these findings show that calcipotriol can enhance the EMT and growth suppression of OS subtypes.
Figure 10 Effects of vitamin D3 synthetic analogue calcipotriol on MG63 and LM7 cells. (A) The structural formula of calcipotriol, a synthetic vitamin D3 analogue and VDR agonist. (B) MTT (3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide) assay to measure cell proliferation 48 hours post-treatment. One-way ANOVA combined with Dunnett’s multiple comparison analysis; p ≤ *0.05, ***0.001, ****0.0001 (n=2). Bar = 20µm (C) Scratch migration assay employing LM7 cells treated with calcipotriol. For each condition, the distance of the scratch was measured across the scratch. Tukey’s multiple comparison test for two-way ANOVA; p ≤ *0.05, **0.01, ****0.0001 (n=4). Bar = 20µm (D) Frequency distribution of spheroids in soft agar treated with calcipotriol. Composite represent the background corrected, threshold-applied ImageJ post-processed images. One-way ANOVA; Šídák’s multiple comparisons test versus untreated (n=3). Bar = 1mm (E) 1,25(OH)2D and calcipotriol decrease collagen production and laminar deposition within MG63 spheroids. MG63 cells in soft agar for 14 days were immunostained for types 1 and 3 collagen. Arrows depict collagen aggregations in the magnified insets. Two-way ANOVA Multiple comparisons using Tukey’s test; p ≤ *0.05, ***0.001 (n=3). Bar = 40µm (F) qPCR analysis of MG63 and LM7 osteosarcomas treated with calcipotriol. Two-way ANOVA Multiple comparisons using Tukey’s test; p ≤ **0.01, ****0.0001 (n=4). LM7 xenograft metastatic model in Nu/J (nude) mice. The images are from 5-week-old Nu/J mice that had LM7 cells delivered to their right flank. After xenograft, mice were immediately given either vehicle or calcipotriol (60 μg/kg b.w.) treatment until tissue harvest. NS, not significant.
Calcipotriol inhibits
留言 (0)